Resazurin-Based Assay for Quantifying Living Cells during Alkaline Phosphatase (ALP) Release

Abstract
:1. Introduction
2. Methodology
2.1. Reagents
2.2. Instrumentation
2.3. Cell Culture
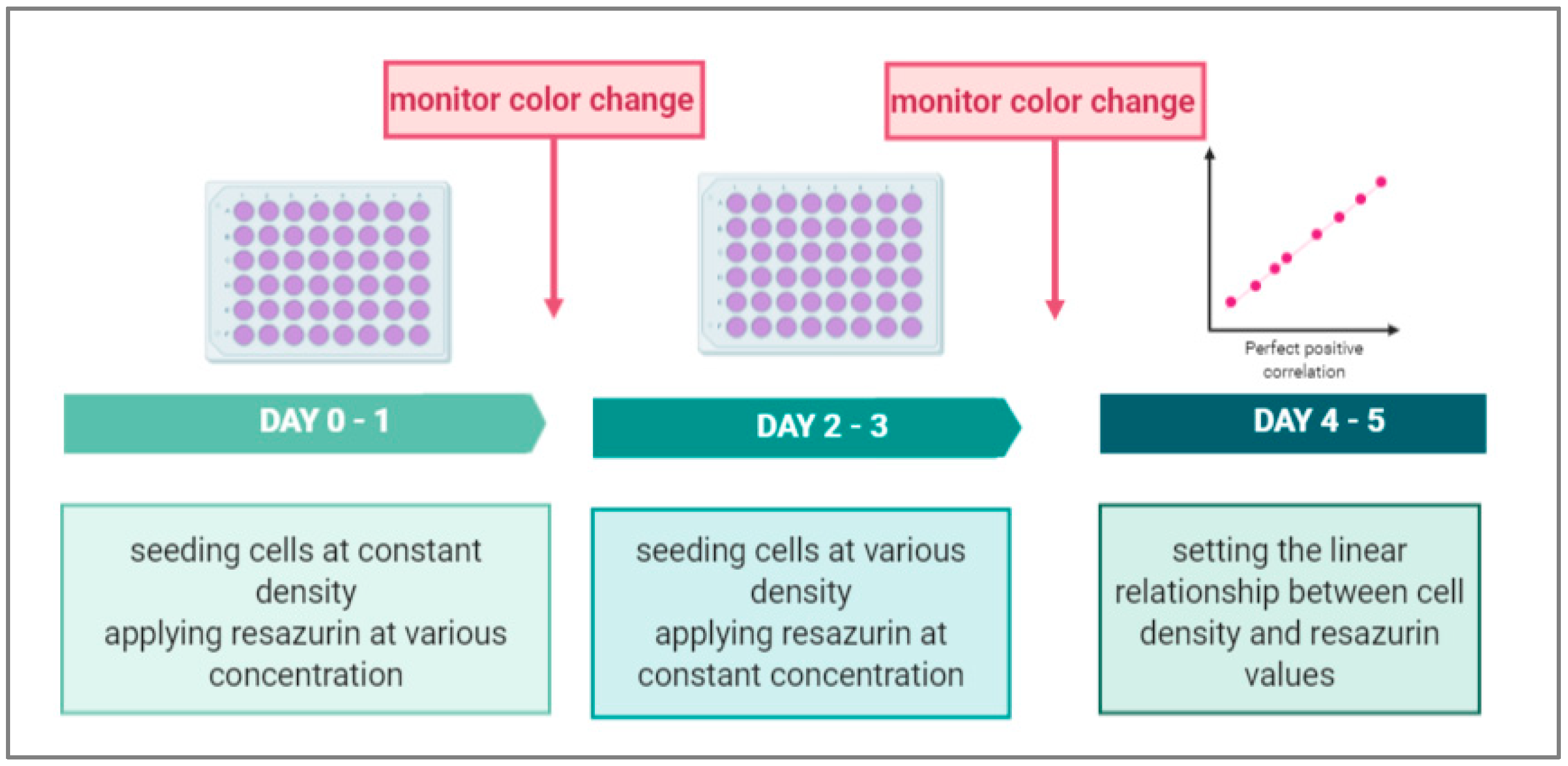
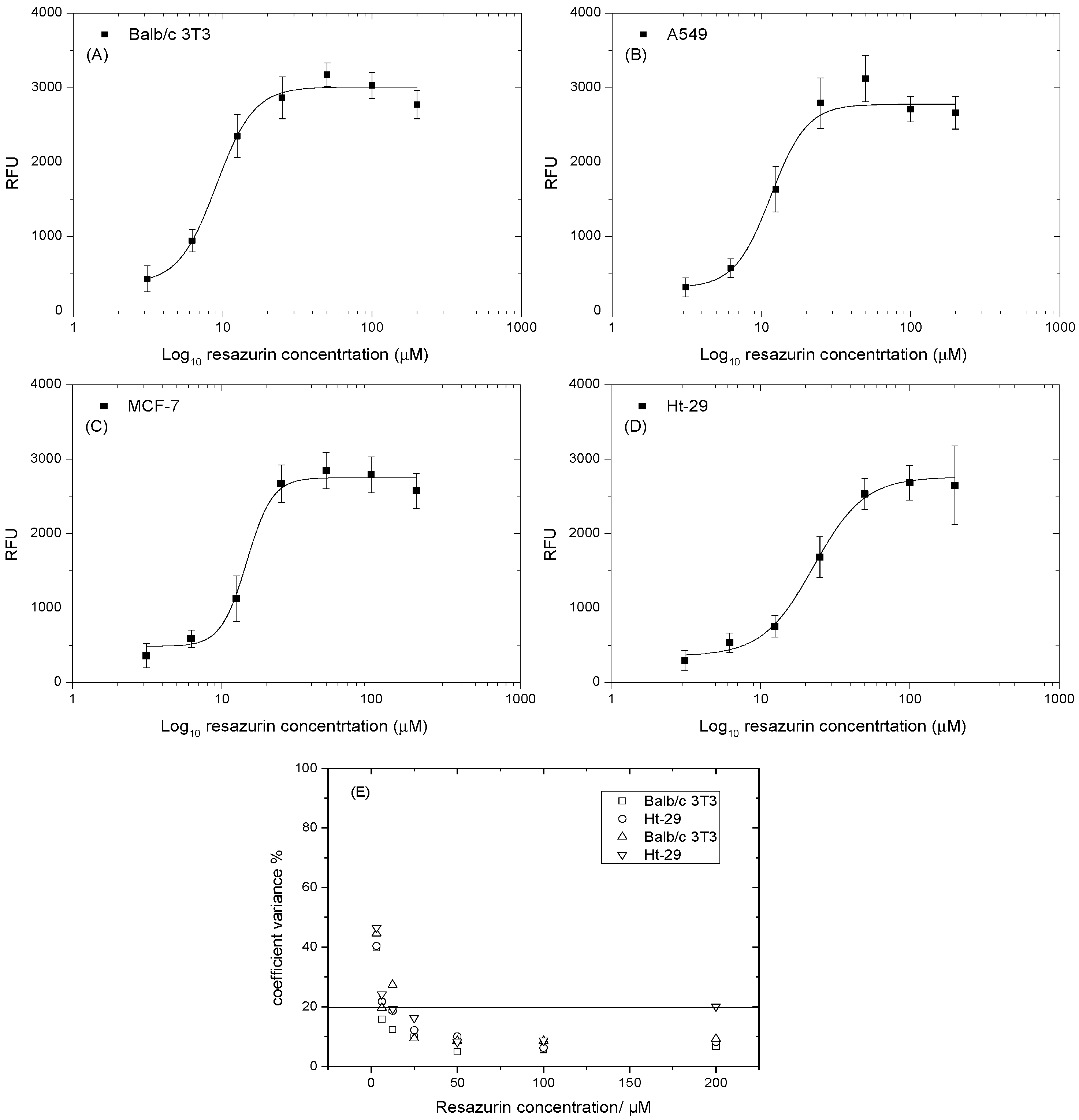
2.4. Determining the Optimal Concentration of Resazurin
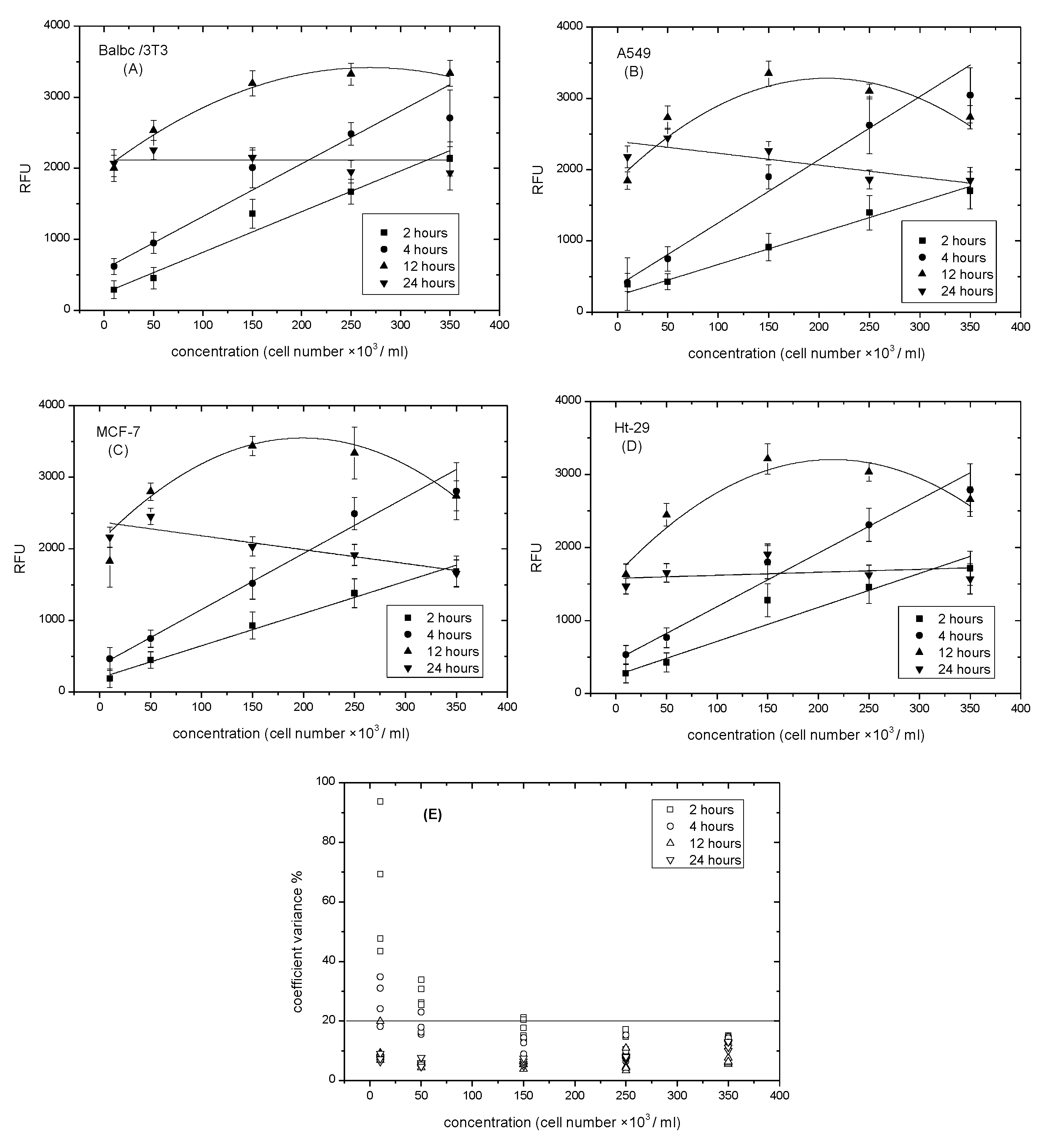
2.5. Determining the Optimal Time of Incubation with Resazurin
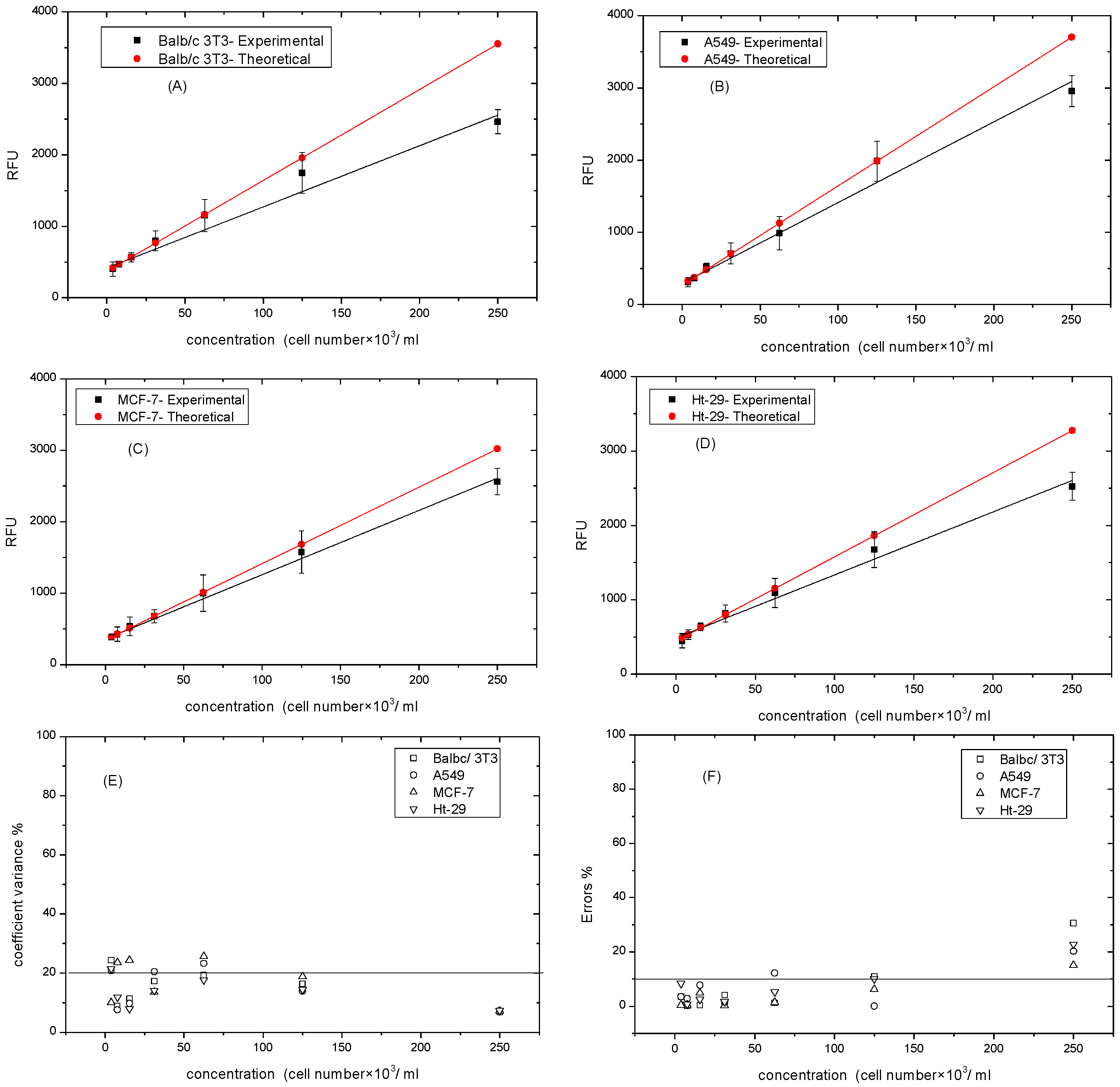
2.6. Determining the Calibration Curve of Resazurin and Cell Numbers
2.7. Determining the Optimal Density of Cell Lines
2.8. Determining Cell Number Based Resazurin Assay
2.9. Microscope Images
2.10. Hemocytometer Assay
2.11. Cell Viability at Post-Confluence Culture
2.12. Alkaline Phosphatase Release from Cells at Post-Confluence Culture
2.13. Data Analysis
3. Results and Discussion
3.1. Determining the Optimal Concentration of Resazurin
3.2. Determining the Optimal Time of Incubation with Resazurin
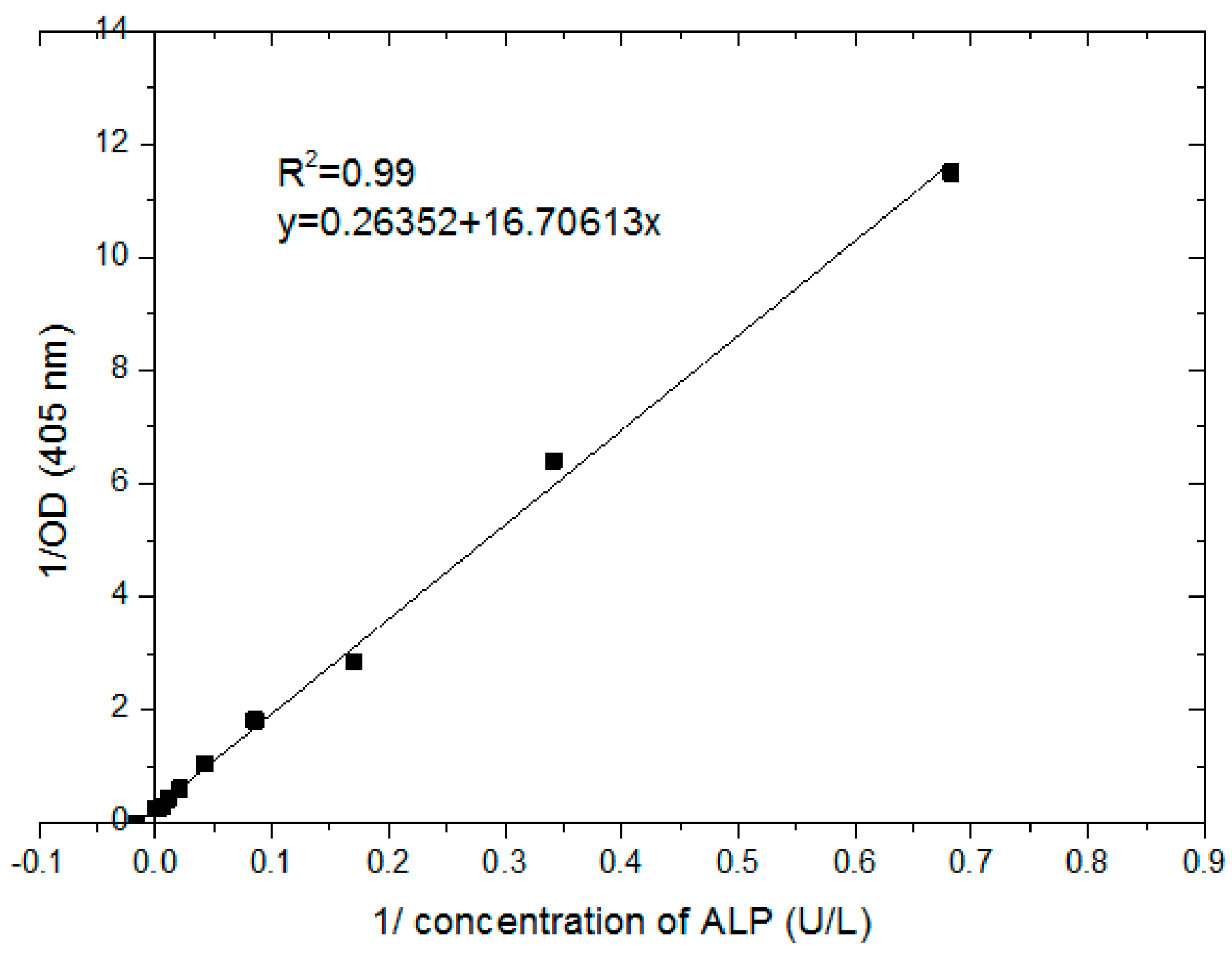
3.3. Determining the Standard Curve of Resazurin and Cell Numbers
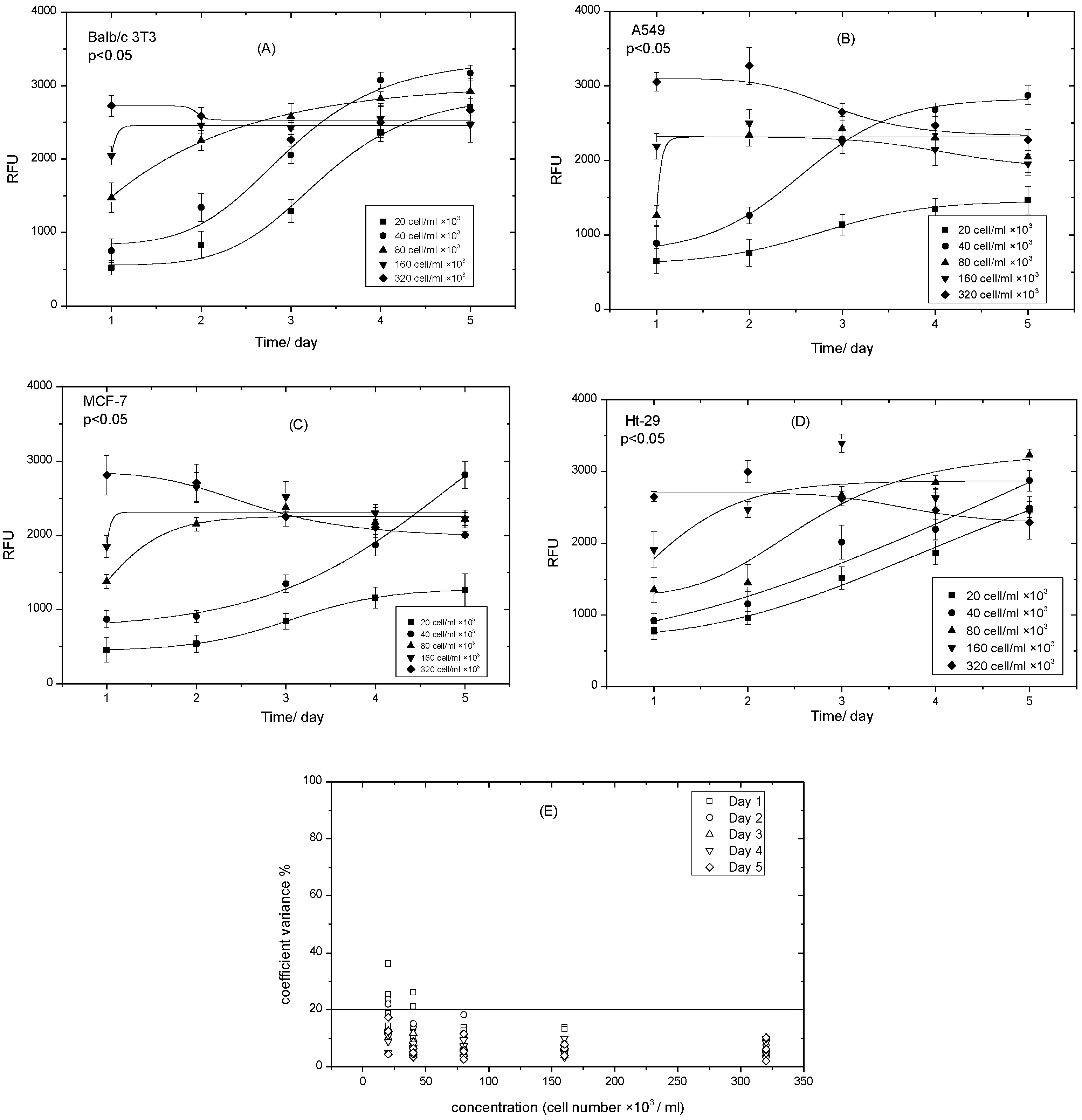
3.4. Determining the Optimal Density of Cell Lines
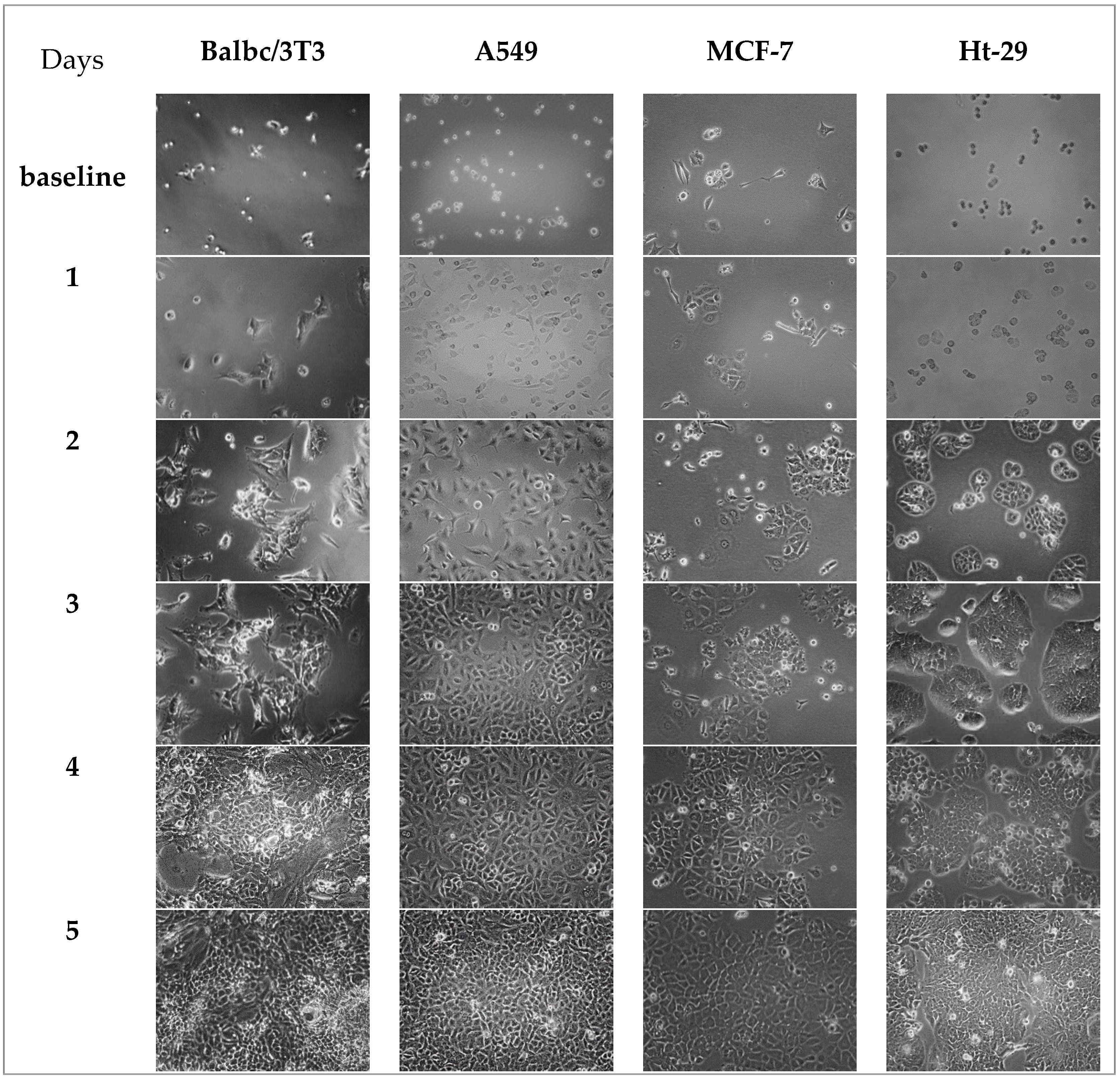
3.5. Microscope Images
3.6. Determining Cell Number and Comparison to Hemocytometer Assay
3.7. Cell Viability at Post-Confluence Culture
3.8. Alkaline Phosphatase Release from Cells at Post-Confluence Culture
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khalili, A.A.; Ahmad, M.R. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef] [Green Version]
- Koev, S.T.; Dykstra, P.H.; Luo, X.; Rubloff, G.W.; Bentley, W.E.; Payne, G.F.; Ghodssi, R. Chitosan: An integrative biomaterial for lab-on-a-chip devices. Lab Chip 2010, 1, 326–342. [Google Scholar] [CrossRef] [PubMed]
- Lukić, M.; Šimec, N.; Zatezalo, V.; Jurenec, S.; Radić-Krišto, D. Exclusion of Trypan blue exclusion test for CD34+cell viability determination: P007. Bone Marrow Transplant. 2017, 52, S126–S127. [Google Scholar]
- Menyhárt, O.; Harami-Papp, H.; Sukumar, S.; Schäfer, R.; Magnani, L.; de Barrios, O.; Győrffy, B. Guidelines for the selection of functional assays to evaluate the hallmarks of cancer. BBA Rev. Cancer 2016, 1866, 300–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzarski, J.S.; Bijonowski, B.M.; Wang, B.; Ward, H.H.; Wandinger-Ness, A.; Miller, W.M.; Wertheim, J.A. Dual-Purpose Bioreactors to Monitor Noninvasive Physical and Biochemical Markers of Kidney and Liver Scaffold Recellularization. Tissue Eng. Part C Methods 2015, 21, 132–1043. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Tapias, L.F.; Jank, B.J.; Mathisen, D.J.; Lanuti, M.; Ott, H.C. Ex vivo non-invasive assessment of cell viability and proliferation in bio-engineered whole organ constructs. Biomaterials 2015, 52, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Tapias, L.F.; Gilpin, S.E.; Ren, X.; Wei, L.; Fuchs, B.C.; Tanabe, K.K.; Lanuti, M.; Ott, H.C. Assessment of Proliferation and Cytotoxicity in a Biomimetic Three-Dimensional Model of Lung Cancer. Ann. Thorac. Surg. 2015, 100, 414–421. [Google Scholar] [CrossRef]
- Ansar Ahmed, S.; Gogal, R.M.; Walsh, J.E. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes: An alternative to [3H] thymidine incorporation assay. J. Immunol. Methods 1994, 170, 211–224. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Shum, D.; Radu, C.; Kim, E.; Cajuste, M.; Shao, Y.; Seshan, V.E.; Djaballah, H. A high density assay format for the detection of novel cytotoxic agents in large chemical libraries. J. Enzym. Inhib. Med. Chem. 2008, 23, 931–945. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Jang, S. Optimization of a resazurin-based microplate assay for large-scale compound screenings against Klebsiella pneumoniae. 3 Biotech 2018, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, F.; Rigole, P.; Brackman, G.; Coenye, T. Optimization of resazurin-based viability staining for quantification of microbial biofilms. J. Microbiol. Methods 2014, 98, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Chadha, S.; Kale, S. Simple fluorescence-based high throughput cell viability assay for filamentous fungi. Lett. Appl. Microbiol. 2015, 61, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Goughenour, K.D.; Balada-Llasat, J.-M.; Rappleye, C.A. Quantitative microplate-based growth assay for determination of antifungal susceptibility of histoplasma capsulatum yeasts. J. Clin. Microbiol. 2015, 53, 3286–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, K.; Surand, J.; Le Dantec, A.; Mosrin-Huaman, C.; Legrand, A.; Rahmouni, A. Recombinant yeast and human cells as screening tools to search for antibacterial agents targeting the transcription termination factor Rho. J. Antibiot. 2018, 71, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowling, T.; Mercer, L.; Don, R.; Jacobs, R.; Nare, B. Application of a resazurin-based high-throughput screening assay for the identification and progression of new treatments for human African trypanosomiasis. Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Rolón, M.; Vega, C.; Escario, J.A.; Gómez-Barrio, A. Development of resazurin microtiter assay for drug sensibility testing of Trypanosoma cruzi epimastigotes. Parasitol. Res. 2006, 99, 103–107. [Google Scholar] [CrossRef]
- Uzarski, J.S.; DiVito, M.D.; Wertheim, J.A.; Miller, W.M. Essential design considerations for the resazurin reduction assay to noninvasively quantify cell expansion within perfused extracellular matrix scaffolds. Biomaterials 2017, 129, 163–175. [Google Scholar] [CrossRef]
- Latner, A.L.; Skillen, A.W. Isoenzymes in Biology and Medicine; Academic Press Inc.: London, UK, 1968. [Google Scholar]
- Sharma, U.; Pal, D.; Prasad, R. Alkaline Phosphatase: An Overview. Indian J. Clin. Biochem. 2014, 29, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Acton, Q.A. Transforming Growth Factors—Advances in Research and Application; Scholarly Editions: 2013. Available online: https://books.google.td/books?id=dARP4d1Io3wC (accessed on 12 April 2019).
- Herz, F.; Halwer, A.S.M.; Bogart, L.H. Alkaline phosphatase in HT-29, a human colon cancer cell line: Influence of sodium butyrate and hyperosmolality. Arch. Biochem. Biophys. 1981, 210, 581–591. [Google Scholar] [CrossRef]
- Tsai, L.-C.; Hung, M.-W.; Chen, Y.-H.; Su, W.-C.; Chang, G.-G.; Chang, T.-C. Expression and regulation of alkaline phosphatases in human breast cancer MCF-7 cells. Eur. J. Biochem. 2000, 267, 1330–1339. [Google Scholar] [CrossRef] [PubMed]
- Hui, M.; Hu, M.; Tenenbaum, H.C. Changes in cell adhesion and cell proliferation are associated with expression of tissue non-specific alkaline phosphatase. Cell Tissue Res. 1993, 274, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, S.; Monteiro, R.; Martins, M.J.; Calhau, C.; Azevedo, I.; Soares, R. Distinct modulation of alkaline phosphatase isoenzymes by 17beta-estradiol and xanthohumol in breast cancer MCF-7 cells. Clin. Biochem. 2007, 40, 268. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Lewis, J.A. Molecular Biology of the Cell, 4th ed.; Studying Gene Expression and Function; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Yang, T.-T.; Sinai, P.; Kain, S.R. An Acid Phosphatase Assay for Quantifying the Growth of Adherent and Nonadherent Cells. Anal. Biochem. 1996, 241, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Eder, W.; Castaneda, J.; Doss, M.; Huber, E.; Ebner, R.; Kunz-Schughart, L.A. A reliable tool to determine cell viability in complex 3-D culture: The acid phosphatase assay. J. Biomol. Screen. 2007, 12, 925–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, D.P.; Parker, T.L.; Walker, D.A.; Alexander, C.; Ashford, M.B.; Gellert, P.R.; Garnett, M.C. Multiplexing spheroid volume, resazurin and acid phosphatase viability assays for high-throughput screening of tumour spheroids and stem cell neurospheres. PLoS ONE 2014, 9, e103817. [Google Scholar] [CrossRef] [Green Version]
- Joanne, K.; Itano, J.B.; Conde, F.; Saria, M. Core Curriculum for Oncology Nursing, 5th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Pace, R.T.; Burg, K.J.L. Toxic effects of resazurin on cell cultures. Cytotechnology 2015, 67, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Hamalainen-Laanaya, H.K.; Orloff, M.S. Analysis of cell viability using time-dependent increase in fluorescence intensity. Anal. Biochem. 2012, 429, 32–38. [Google Scholar] [CrossRef]
- Zheng, F.; Wang, S.; Hou, W.; Xiao, Y.; Liu, P.; Shi, X.; Shen, M. Comparative study of resazurin reduction and MTT assays for cytocompatibility evaluation of nanofibrous materials. Anal. Methods 2019, 11, 483–489. [Google Scholar] [CrossRef]
- Adams, D.L.; Zhu, P.; Makarova, O.V.; Martin, S.S.; Charpentier, M.; Chumsri, S.; Li, S.; Amstutz, P.; Tang, C.-M. The systematic study of circulating tumor cell isolation using lithographic microfilters. Rsc Adv. 2014, 4, 4334–4342. [Google Scholar] [CrossRef]
- Majeska, R.J.; Rodan, G.A. The Effect of 1,25(OH)2D3 on Alkaline-Phosphatase In Osteoblastic Osteo-Sarcoma Cells. J. Biol. Chem. 1982, 257, 3362–3365. [Google Scholar] [PubMed]
- Castrén, E.; Sillat, T.; Oja, S.; Noro, A.; Laitinen, A.; Konttinen, Y.T.; Lehenkari, P.; Hukkanen, M.; Korhonen, M. Osteogenic differentiation of mesenchymal stromal cells in two-dimensional and three-dimensional cultures without animal serum. Stem Cell Res. Ther. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Zhang, Y.; Wang, J.; Yu, W.; Wang, W.; Ma, X. Monitoring of Cell Viability and Proliferation in Hydrogel-Encapsulated System by Resazurin Assay. Appl. Biochem. Biotechnol. 2010, 162, 1996–2007. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Martins, A.M. The strong correlation between alkaline phosphatase activity and cell viability. Cell Biol. Int. 2008, 32, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Akcakaya, H.; Aroymak, A.; Gokce, S. A quantitative colorimetric method of measuring alkaline phosphatase activity in eukaryotic cell membranes. Cell Biol. Int. 2007, 31, 186–190. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Balb/c 3T3 | A549 | MCF-7 | Ht-29 | |
|---|---|---|---|---|---|
| Complete media | Media | DMEM | DMEM | MEME | McCoy’s 5A |
| 10 % Serum | NBCS | FBS | |||
| Balb/c 3T3 | A549 | MCF-7 | Ht-29 | |
|---|---|---|---|---|
| Day 4 | 3091.61 ± 28.74 | 2784.46 ± 43.35 | (1887.34 ± 5.36) | (2980.70 ± 44.51) |
| Day 5 | 2513.44 ± 7.77 *§ | (1952.29 ± 0.82) *§ | (1787.48 ± 35.99) | (2655.06 ± 34.07) * |
| Day 6 | 1872.74 ± 6.72 *§ | (1415.29 ± 44.35) *§ | (1407.01 ± 66.13) *§ | (2512.21 ± 48.32) * |
| VC | (412.6 ± 9.9) *§ | (597.6 ± 23.3) *§ | (237.8 ± 3.1) *§ | (623.8 ± 4.5) *§ |
| RC | (97.2 ± 6.3) *§ | (78.7 ± 0.9) *§ | (80.5 ± 0.3) *§ | (82.7 ± 0.7) *§ |
| Quantity of ALP Release (U/L) in Each Day | |||||
|---|---|---|---|---|---|
| Day 4 | Day 5 | Day 6 | VC | Blank | |
| Balb/c 3T3 | * | * | * | * | |
| A549 | * | * | * | * | |
| MCF-7 | * | * | * | * | |
| Ht-29 | * | * | * | * | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balbaied, T.; Moore, E. Resazurin-Based Assay for Quantifying Living Cells during Alkaline Phosphatase (ALP) Release. Appl. Sci. 2020, 10, 3840. https://doi.org/10.3390/app10113840
Balbaied T, Moore E. Resazurin-Based Assay for Quantifying Living Cells during Alkaline Phosphatase (ALP) Release. Applied Sciences. 2020; 10(11):3840. https://doi.org/10.3390/app10113840
Chicago/Turabian StyleBalbaied, Thanih, and Eric Moore. 2020. "Resazurin-Based Assay for Quantifying Living Cells during Alkaline Phosphatase (ALP) Release" Applied Sciences 10, no. 11: 3840. https://doi.org/10.3390/app10113840
APA StyleBalbaied, T., & Moore, E. (2020). Resazurin-Based Assay for Quantifying Living Cells during Alkaline Phosphatase (ALP) Release. Applied Sciences, 10(11), 3840. https://doi.org/10.3390/app10113840