1. Introduction
Microalgae are a diverse group of microorganisms that combine properties of higher plants with those of some prokaryotes [
1,
2]. They are ubiquitous, living either in aquatic (fresh and seawater) and terrestrial environments [
3]. Microalgae occur as unicellular or simple colonial organisms, usually displaying rapid growth rates [
4] and photosynthetic efficiencies between 9–10%, in general significantly higher than those of terrestrial plants [
5,
6,
7]. They are key members of the biosphere, as they play an important role in the global carbon and nitrogen cycles. By using light energy to produce organic matter, algal photosynthesis contributes to around 50% of the total planetary primary production [
8,
9]. Cyanobacteria, previously called “blue-green algae”, are even able to fix atmospheric nitrogen (N
2) into organic nitrogen and are the major contributors to global biological nitrogen fixation [
10,
11].
Being a biodiverse resource, microalgae can produce a wide range of value-added molecules such as proteins, lipids, carbohydrates, pigments and other compounds for various product applications [
12,
13]. Of these, carotenoids, phycobiliproteins, and polyunsaturated fatty acids (PUFA) such as eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids are the most widely explored. Microalgae can be applied to food, feed, biofertilizer, and cosmetic industries [
14]. They can also play a major role in the pharmaceutical industry as sources of novel biochemicals for the development of new drug leads to overcome the growing problem of antibiotic resistance in many bacterial pathogens [
15], since some microalgae produce bioactive compounds with antibacterial activity [
16]. Besides antimicrobial compounds, microalgae produce also antiprotozoal agents and anti-inflammatory, antioxidant and anticancer compounds [
17]. The biomass of oleaginous microalgae is a sustainable, renewable and eco-friendly feedstock which can be a source for the production of biofuels and bioenergy [
14,
18]. Microalgae can even be used in wastewater treatment and its photosynthetic cultivation is one of the most promising carbon dioxide (CO
2) mitigation technologies [
19].
The focus on research into the genus
Nannochloropsis has increased over the years [
20]. These microalgae are considered to be oleaginous models because of their high lipid productivity and photosynthetic efficiency, having also a well-established genetic toolbox [
21,
22,
23]. Several species of the genus
Nannochloropsis have been successfully cultivated autotrophically at large scale due to the relatively mature technology for outdoor large-scale cultivation of these eustigmatophytes [
22].
Despite their biodiversity, microalgae are still an underexploited biological resource. There are only a few long-standing companies that have been able to scale up their production of microalgal biomass in industrial facilities [
12,
23,
24]. The economics of large scale facilities are often not favorable to short-term investment, because operational and capital costs are high and the payback is usually longer than expected [
25].
Microalgae can be cultivated either in open or closed systems. Despite having several drawbacks, open systems are the most widely applied bioreactors in industrial microalgal processes, since they require lower capital and operational costs than closed systems [
26]. Microalgal open production systems include circular and raceways ponds as well as cascade systems [
27]. Among them, raceway ponds are the most commonly used open photobioreactor for commercial microalgal production [
28]. These are shallow, hairpin-shaped, closed-loop flow channels, usually 0.15–0.30 m deep, internally lined with cement, polyvinyl chloride (PVC) or clay, with mixing and aeration provided through circulation by a rotating paddlewheel [
29,
30,
31,
32]. A straight channel velocity of at least 0.2 m s
−1 is needed to ensure a velocity of 0.1 m s
−1, the minimum required to prevent sedimentation of microalgal biomass in the whole raceway [
33]. A 0.3 m s
−1 flow velocity is the most commonly used flow velocity for biomass production [
33]. The surface-to-volume ratio in raceways is always 1/pond depth. A lower depth increases the surface-to-volume ratio, improving light penetration [
34]. Thus, for autotrophic growth, shallower ponds are more productive than deeper ponds [
34]. However, at water depths lower than 0.15 m, a severe decrease in flow and turbulence occurs in large-scale raceways [
35]. This long light path results in large areal volumes (150 L m
−2) and cell concentrations of less than 0.6 g L
−1, which promote the occurrence of contaminants (i.e., grazers and competing microalgae) [
35]. Another problem of open systems is the low productivities obtained, due to temperature fluctuations, inefficient mixing and CO
2 dispersion, light limitations and evaporation losses [
29]. Typically, well-managed raceway ponds may achieve 20–25 g m
−2 d
−1 for short periods, while long-term productivity in large commercial raceways rarely exceeds 12-13 g m
−2 d
−1 [
35]. López et al. [
36] point out that 20 g m
−2 d
−1 is the threshold for the average annual productivity, which is rarely exceeded in large-scale outdoor production.
Several studies have been carried out in order to study the design of the raceway pond, with the aim of reducing the energy costs and increasing the growing efficiency of microalgae. Most of the energy losses occur at the hairpin bends. Liffman et al. [
37] used computational fluid dynamics (CFD) to model different bend configurations. These authors showed that, when compared with the traditional constant-width/constant-depth bend configuration, some of the new bend configurations not only minimize the energy consumption, but also improve the mixing of the raceway ponds by depletion of low speed and stagnant regions [
37]. Another CFD-based approach demonstrated that a raceway configuration with a minimum of three semicircular deflector baffles and an “island” at the end of the central divider, apart from being more energy-efficient than the standard configuration, can completely prevent the development of dead zones [
32].
New design studies rely mostly on modeling and very few in actual experiments. Despite the evolution of the models and the associated technology, errors are always encountered when compared to experimental data [
38,
39]. In this context, this work aimed to compare the performance of two raceway ponds with different configurations based on three criteria: areal productivity, energy consumption, and the biochemical composition of biomass produced using
Nannochloropsis oceanica as model microalga.
2. Materials and Methods
All experiments were performed between February 4th and July 28th of 2019 at Allmicroalgae’s facilities at Pataias, Portugal. The microalga strain, N. oceanica CCAP 849/10, was obtained ‘from Culture Collection of Algae and Protozoa and is kept at Allmicroalgae’s culture collection.
2.1. Raceways Design, Commissioning and Construction
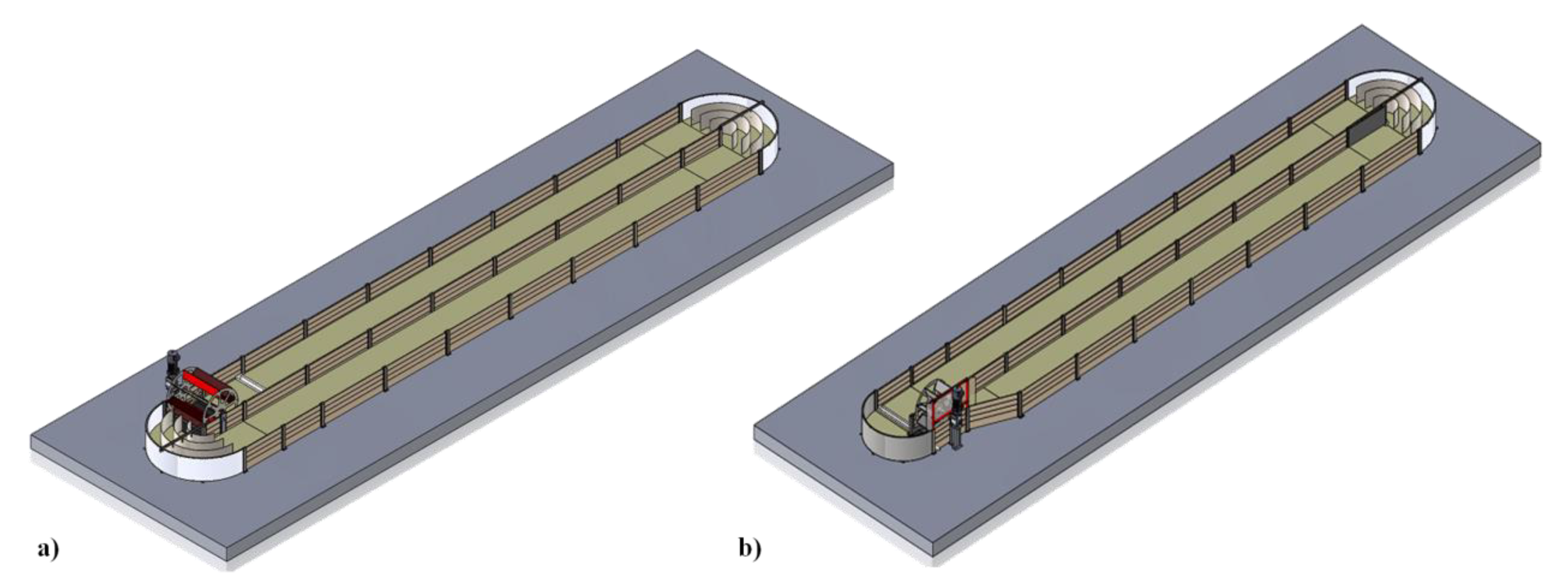
Based on the literature review and Allmicroalgae industrial experience in the growth of microalgae, two raceway pond prototypes were designed, as shown in
Figure 1: one standard and one with a modified configuration. The standard raceway included the most common dimensions, configuration and construction materials reported, while the modified version considered several aspects previously reviewed from the literature and indicated as having a positive influence in the production of microalgal biomass.
The starting point was a typical raceway with two bends, as ponds with more than two bends have higher energy consumption [
32]. To avoid the formation of dead zones as much as possible, three baffles were included in the circular endings of the ponds. Raceway ponds with around 28 m
2 were developed. They were 15 m long, 2 m wide and 1 m channel width. The depth was fixed at 0.375 m and other structural dimensions are listed in
Table 1. The mixer consisted of a single eight-flat-bladed paddlewheel installed right after one of the semicircular ends at the beginning of one of the channels.
The modified raceway model included a loop-shaped end of the central divider opposite to the paddlewheel. The ‘loop’ was intended to prevent the development of dead zones in the vicinity of this end of the divider. Besides, the paddlewheel hypothesized for the modified model had a width of 0.475 m, as compared to the standard model paddlewheel which measured 0.870 m. A picture of the raceways can be seen in
Figure 2.
Figure 3 shows a detailed view of the modified raceway.
The raw materials used for the construction of both raceways were the same. Polyethylene film and PVC waterproofing geomembrane were used in the base of the raceways to waterproof them. The structure was supported by stone powder and stainless steel and external walls were made of treated wood.
2.2. Trials
Each trial lasted 13 days and was performed simultaneously in both raceways.
N. oceanica was inoculated at a 0.3 g L
−1 concentration in Allmicroalgae’s base medium, based on Guillard’s F/2 medium [
40], with 30 g L
−1 NaCl and 0.31 g L
−1 NO
3−. In all experiments, a fresh inoculum sequentially scaled up, was used. Initially, the cultures were grown in 5 L flasks during 7 to 15 days. When the culture reached 1.2 g L
-1, 20 × 5 L flasks of culture (100 L) were used to inoculate a single flat panel (FP), which was later divided into the other three FPs. These were used to inoculate the two raceway ponds. Throughout the assay, the culture was closely monitored. The pH was kept between 8.0–8.2, through the injection of pure CO
2 to the culture. The NO
3− concentration was always kept above 0.12 g L
−1 and water refilled whenever it was necessary. Regular microscopic observations were performed to monitor contaminations. At the end of the assay the culture was harvested and centrifuged at 4000×
g for 30 min, and the resulting biomass and supernatant were biochemically analyzed. All the assays were made in duplicate.
2.2.1. Water Depth
Two different water depths were tested, 0.20 and 0.13 m, with a flow velocity of 0.30 m s−1.
2.2.2. Flow Velocity
The flow velocity optimization trial was performed using the standard raceway. A velocity of 0.15 m s−1 was used with a 0.13 m water column.
2.3. Growth Assessment
Samples (50 mL) were taken on a daily basis and microscopic observation was performed in order to monitor contaminations, which were never above 2% of cell counts.
Optical density (OD) and dry weight (DW) were measured to follow microalgal growth. OD was performed at 540 nm using a spectrophotometer (4251/50, Zuzi, Spain). DW was determined using a 0.7 µm glass fiber filter (VWR, Portugal) in a moisture analyzer (MA 50.R, RADWAG, Poland). In order to neutralize the salt weight, the culture was washed with an equivalent volume of ammonium formate solution at 35 g L−1.
Areal biomass productivity,
P (g L
−1 d
−1), was determined by Equation (1):
where
X2 (g L
−1) and
X1 (g L
−1) represented the DW times
t1 (s) and
t2 (s), respectively,
V (m
3) was the raceway volume and
A (m
2) the ground area occupied by the bioreactor.
Specific growth rate,
µ (d
−1), was expressed as a ratio between the difference final and initial logarithmic biomass concentrations (
X2 and
X1, respectively) and the associated time range (
t2 and
t1, respectively), as seen in Equation (2).
Global productivity and growth rate refer to the whole assay (13 days). Maximum productivity and growth rate were calculated based on the highest values obtained daily.
The lipid production/energy consumption ratio (
L/
kW) was calculated through the following equation:
where
L (%) was the lipid content, 13 days the total extension of the assay, and
p the power consumption.
The photosynthetic efficiency (PE) was determined by equation 4, as previously described by Quelhas et al. (2019) [
41].
where rad (kW m
−2) was the radiation that reaches the reactor,
t (s) was the time between solar radiation measurement and HHV was the higher heating value, calculated based on elemental (CHN) analysis by Equation (5), as described by Callejón-Ferre et al. [
42]:
A spectrophotometric analysis (Genesys 10S UV-VIS spectrophotometer (Thermo Scientific, Massachusetts, USA)) was used to measure the nitrate concentration of the medium. Samples were centrifuged for 10 min at 2000 g (MiniStar Silverline, VWR, Portugal). The OD of the sample was calculated from the difference at 220 nm and twice the value at 275 nm and compared with a calibration curve with sodium nitrate [
43]. The blank solution was made with 0.1 mol L
−1 of HCl.
2.4. Proximate Composition
The ash content was determined in a furnace (J. P. Selecta, Sel horn R9-L) at 550 °C for 8 h.
Total lipids were determined through a gravimetric analysis, using a previously modified method of Bligh and Dyer (1959) [
44], and reported by Pereira et al. [
45]. A biomass weight ranging 5–10 mg was solubilized in 0.8 mL of distilled water. A mixture of methanol (2 mL), chloroform (2 mL) and distilled water (1 mL) was used to extract the lipid fraction from the biomass [
44]. After centrifugation for 10 min at 2500 g, 0.7 mL of the chloroform fraction was transferred to a pre-weighed tube. A heat treatment at 60 °C (QBD2, Grant Instruments, UK) to evaporate all the chloroform was performed and after 3 h in the desiccator, the final weight of the tube was measured in the precision balance (M5P, Sartorius, Germany).
CHN analysis was performed using a Vario el III (Vario EL, Elementar Analyser system, Germany), according to Quelhas et al. [
41]. Total proteins were calculated by multiplying the nitrogen content by the standard conversion factor 6.25 [
46] after CHN analysis. Carbohydrates were considered the remaining mass.
2.5. Fatty Acid Profile
Fatty acids were determined by means of its conversion to fatty acid methyl esters (FAME), according to a modified method of Lepage and Roy [
47] described by Pereira et al. [
48]. FAME were analyzed in a gas chromatography-mass spectrometer (GC-MS) (Bruker SCION 456/GC, SCION TQ MS). The gas chromatograph was equipped with a 30 m length column with an internal diameter of 0.25 mm and a film thickness of 0.25 µm (ZB-5MS, Phenomenex, Torrance, California, US). Helium was the carrier gas. The temperature program was 60 °C (1 min), 30 °C min
−1 to 120 °C, 5 °C min
−1 to 250 °C, and 20 °C min
−1 to 300 °C (2 min). The total running time was 31.5 min. Injection temperature was 300 °C. Supelco
® 37 Component FAME Mix (Sigma-Aldrich, Sintra, Portugal) were used as standards. After identified and quantified, the FAME profile was expressed as a relative percentage of the total fatty acids content present in the biomass.


 ,
, 





{kind=link}
{kind=link}
{kind=link}