Long-Term Exposure to Azo Dyes from Textile Wastewater Causes the Abundance of Saccharibacteria Population

 , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
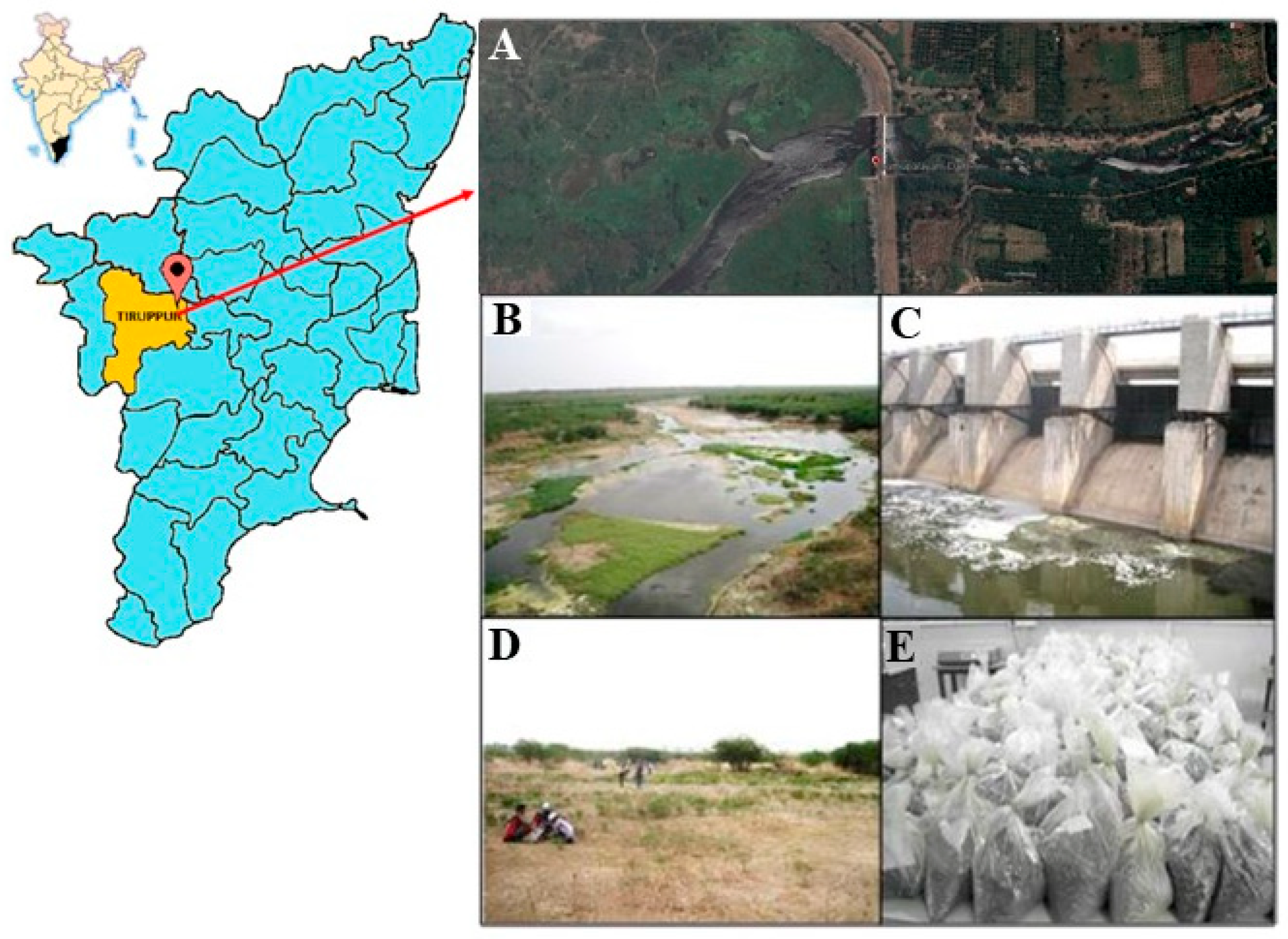
2.1. Collection of Soil Sample
2.2. Soil Enzyme Activities
2.3. Soil DNA Extraction
2.4. PCR Amplification and Phylogenetic Marker Library Preparation
2.5. Pyrosequencing and Pre-Processing of Sequence Reads
2.6. Data Analysis
3. Results and Discussion
3.1. Soil Enzyme Activities
3.2. Diversity Indices
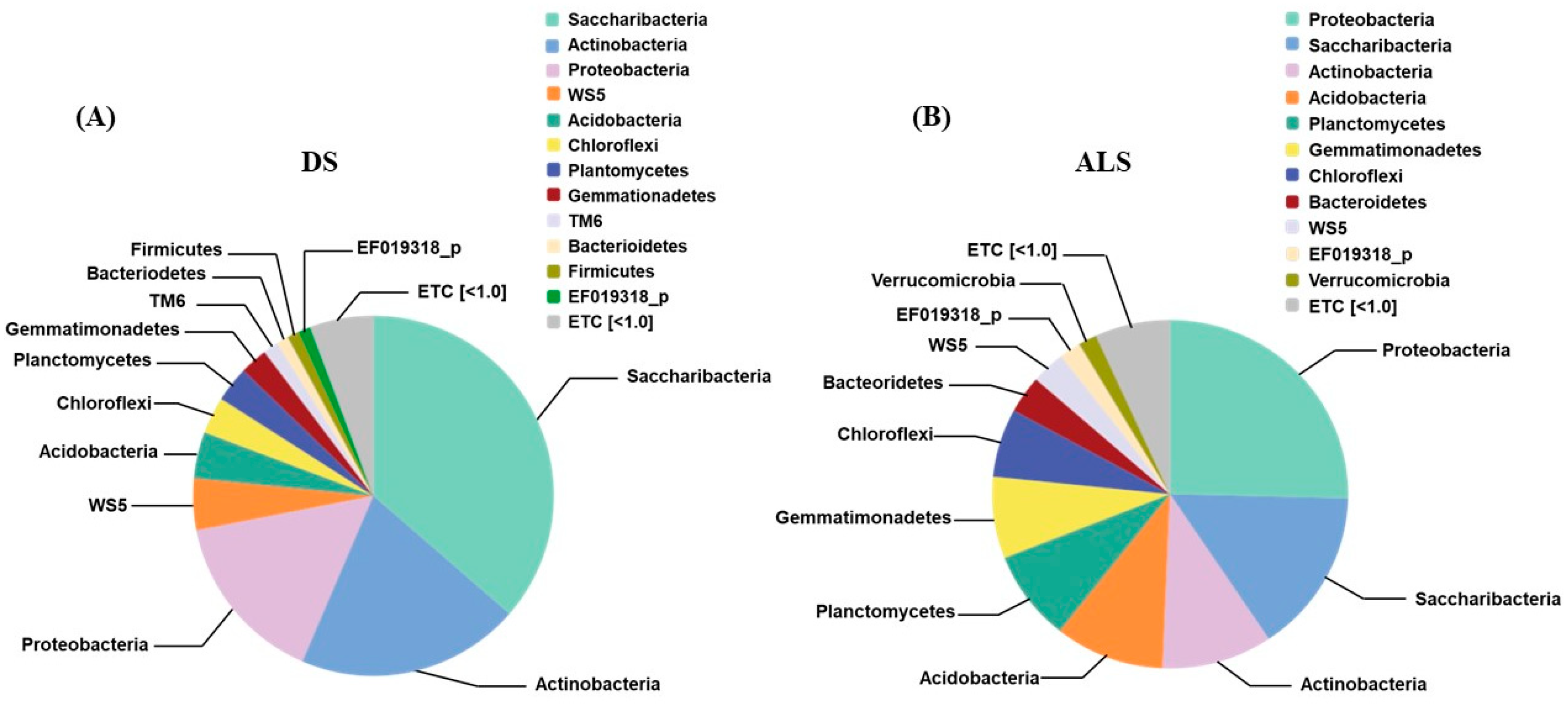
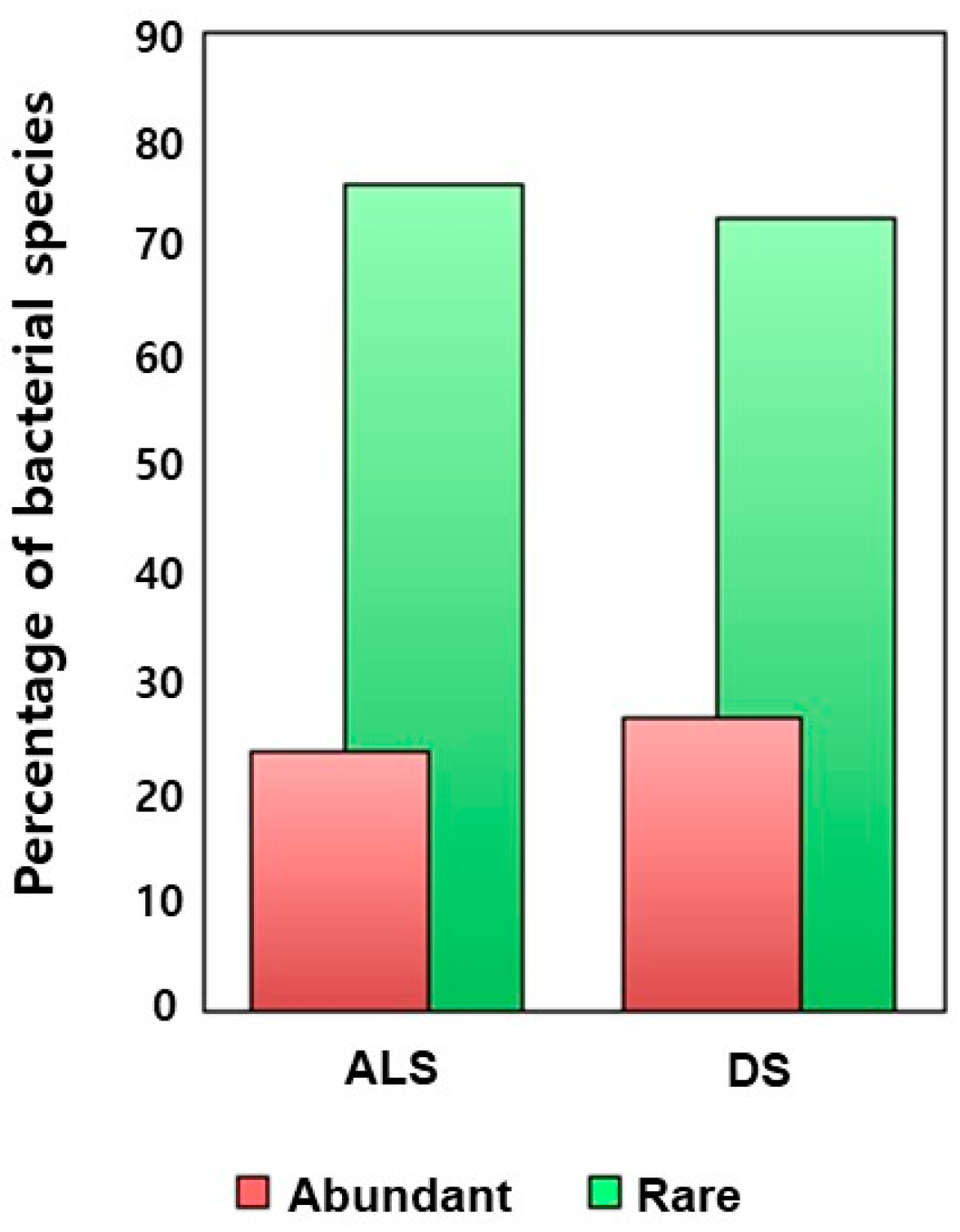
3.3. Bacterial Diversity and Community Profiles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuppusamy, S.; Thavamani, P.; Megharaj, M.; Venkateswarlu, K.; Lee, Y.B.; Naidu, R. Pyrosequencing analysis of bacterial diversity in soils contaminated long-term with PAHs and heavy metals: Implications to bioremediation. J. Hazard. Mater. 2016, 317, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Philippot, L.; Park, W. Metagenomic and functional analyses of the consequences of reduction of bacterial diversity on soil functions and bioremediation in diesel-contaminated microcosms. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Zhang, J.; Ren, L.; Zhou, Y.; Gao, J.; Luo, L.; Yang, Y.; Peng, Q.; Huang, H.; Chen, A. Diagnosis of soil contamination using microbiological indices: A review on heavy metal pollution. J. Environ. Manag. 2019, 242, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.I.; Biswas, B.; Smith, E.; Naidu, R.; Megharaj, M. Toxicity assessment of fresh and weathered petroleum hydrocarbons in contaminated soil-a review. Chemosphere 2018, 212, 755–767. [Google Scholar] [CrossRef]
- Imran, M.; Shaharoona, B.; Crowley, D.E.; Khalid, A.; Hussain, S.; Arshad, M. The stability of textile azo dyes in soil and their impact on microbial phospholipid fatty acid profiles. Ecotox. Environ. Safe. 2015, 120, 163–168. [Google Scholar] [CrossRef]
- Singh, L.; Singh, V.P. Textile Dyes Degradation: A Micrbial Approach for Biodegradation of Pollutants. In Microbial Degradation of Synthetic Dyes in Wastewaters, Environmental Science and Engineering; Singh, S.N., Ed.; Springer: Cham, Switzerland, 2015; pp. 187–204. [Google Scholar]
- Phugare, S.S.; Kalyani, D.C.; Surwase, S.N.; Jadhav, J.P. Ecofriendly degradation, decolorization and detoxification of textile effluent by a developed bacterial consortium. Ecotox. Environ. Safe. 2011, 74, 1288–1296. [Google Scholar] [CrossRef]
- Liu, N.; Xie, X.; Yang, B.; Zhang, Q.; Yu, C.; Zheng, X.; Xu, L.; Li, R.; Liu, J. Performance and microbial community structures of hydrolysis acidification process treating azo and anthraquinone dyes in different stages. Environ. Sci. Pollut. Res. 2017, 24, 252–263. [Google Scholar] [CrossRef]
- Rehman, K.; Shahzad, T.; Sahar, A.; Hussain, S.; Mahmood, F.; Siddique, M.H.; Siddique, M.A.; Rashid, M.I. Effect of reactive black 5 azo dye on soil processes related to C and N cycling. PeerJ. 2018, 6, e4802. [Google Scholar] [CrossRef]
- Zhou, Q. Chemical pollution and transport of organic dyes in water–soil–crop systems of the Chinese Coast. Bull. Environ. Contam. Toxicol. 2001, 66, 784–793. [Google Scholar]
- Lee, D.W.; Lee, H.; Kwon, B.O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.J. Biosurfactant assisted bioremediation of crude oil by indigenous bacteria isolated from taean beach sediment. Environ. Pollut. 2018, 241, 254–264. [Google Scholar] [CrossRef]
- Li, Q.; Li, J.; Jiang, L.; Sun, Y.; Luo, C.; Zhang, G. Diversity and structure of phenanthrene degrading bacterial communities associated with fungal bioremediation in petroleum contaminated soil. J. Hazard. Mater. 2020, 403, 123895. [Google Scholar] [CrossRef] [PubMed]
- Punzi, M.; Anbalagan, A.; Börner, R.A.; Svensson, B.M.; Jonstrup, M.; Mattiasson, B. Degradation of a textile azo dye using biological treatment followed by photo-Fenton oxidation: Evaluation of toxicity and microbial community structure. Chem. Eng. J. 2015, 270, 290–299. [Google Scholar] [CrossRef]
- Xu, H.; Yang, B.; Liu, Y.; Li, F.; Song, X.; Cao, X.; Sand, W. Evolution of microbial populations and impacts of microbial activity in the anaerobic-oxic-settling-anaerobic process for simultaneous sludge reduction and dyeing wastewater treatment. J. Clean Prod. 2020. [Google Scholar] [CrossRef]
- Cui, D.; Li, G.; Zhao, D.; Gu, X.; Wang, C.; Zhao, M. Microbial community structures in mixed bacterial consortia for azo dye treatment under aerobic and anaerobic conditions. J. Hazard. Mater. 2012, 221, 185–192. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Jose, P.A.; Ranjith, M.; Anandham, R.; Suganya, K.; Prabhakaran, J.; Thiyageshwari, S.; Johnson, J.; Gopal, N.O.; Kumutha, K.; et al. Decolourisation and degradation of azo dyes by mixed fungal culture consisted of Dichotomomyces cejpii MRCH 1-2 and Phoma tropica MRCH 1–3. J. Environ. Chem. Eng. 2018, 6, 588–595. [Google Scholar] [CrossRef]
- Kaczynski, P.; Lozowicka, B.; Hrynko, I.; Wolejko, E. Behaviour of mesotrione in maize and soil system and its influence on soil dehydrogenase activity. Sci. Total Environ. 2016, 571, 1079–1088. [Google Scholar] [CrossRef]
- Samaddar, S.; Truu, J.; Chatterjee, P.; Truu, M.; Kim, K.; Kim, S.; Seshadri, S.; Sa, T. Long-term silicate fertilization increases the abundance of Actinobacterial population in paddy soils. Biol. Fertil. Soils 2019, 55, 109–120. [Google Scholar] [CrossRef]
- Deng, S.P.; Tabatabai, M.A. Cellulase activity of soils. Soil Biol. Biochem. 1994, 26, 1347–1354. [Google Scholar] [CrossRef]
- Colvan, S.; Syers, J.; O’Donnell, A. Effect of long-term fertiliser use on acid and alkaline phosphomonoesterase and phosphodiesterase activities in managed grassland. Biol. Fertil. Soils 2001, 34, 258–263. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reation-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Bartram, A.K.; Lynch, M.D.; Steams, J.C.; Moreno-Hagelsieb, G.; Neufield, J.D. Generation of multimillion-sequence 16S rRNA gene libraries from complex microbial communities by assembling paired-end Illumina reads. Appl. Environ. Microbiol. 2011, 77, 3846–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteley, A.S.; Jenkins, S.; Waite, I.; Kresoje, N.; Payne, H.; Mullan, B.; Allcock, R.; O’Donnell, A. Microbial 16S rRNA Ion Tag and community metagenome sequencing using the Ion Torrent (PGM) Platform. J. Microbiol. Methods 2012, 91, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Fu, L.; Sun, S.; Li, W. Aritificial and natural duplicates in pyrosequencing reads of metagenomic data. BMC Bioinform. 2010, 11, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server–a public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Shen, T.J. Species Prediction and Diversity Estimation (SPADE) [Software and User’s Guide]. Available online: https://chao.shinyapps.io/SpadeR/http://www.chao.stat.nthu.edu.tw/softwareCE.html (accessed on 18 November 2009).
- Badiane, N.N.Y.; Chotte, J.L.; Pate, E.; Masse, D.; Rouland, C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol. 2001, 18, 229–238. [Google Scholar] [CrossRef]
- Kaczyńska, G.; Borowik, A.; Wyszkowska, J. Soil dehydrogenases as an indicator of contamination of the environment with petroleum products. Water Air Soil Pollut. 2015, 226, 372. [Google Scholar] [CrossRef] [Green Version]
- Lipińska, A.; Kucharski, J.; Wyszkowska, J. The effect of polycyclic aromatic hydrocarbons on the structure of organotrophic bacteria and dehydrogenase activity in soil. Polycycl. Aromat. Compd. 2014, 34, 35–53. [Google Scholar] [CrossRef]
- Yang, J.K.; Zhang, J.J.; Yu, H.Y.; Cheng, J.W.; Miao, L.H. Community composition and cellulase activity of cellulolytic bacteria from forest soils planted with broad-leaved deciduous and evergreen trees. Appl. Microbiol. Biotechnol. 2014, 98, 1449–1458. [Google Scholar] [CrossRef]
- Sandaa, R.A.; Torsvik, V.; Enger, Ø. Influence of long-term heavy-metal contamination on microbial communities in soil. Soil Biol. Biochem. 2001, 33, 287–295. [Google Scholar] [CrossRef]
- Sengupta, A.; Dick, W.A. Bacterial community diversity in soil under two tillage practices as determined by pyrosequencing. Microb. Ecol. 2015, 70, 853–859. [Google Scholar] [CrossRef]
- Feng, G.; Xie, T.; Wang, X.; Bai, J.; Tang, L.; Zhao, H.; Wei, W.; Wang, M.; Zhao, Y. Metagenomic analysis of microbial community and function involved in cd-contaminated soil. BMC Microbiol. 2018, 18, 11. [Google Scholar] [CrossRef] [PubMed]
- Prabha, S.; Gogoi, A.; Mazumder, P.; Ramanathan, A.L.; Kumar, M. Assessment of the impact of textile effluents on microbial diversity in Tirupur district, Tamil Nadu. Appl. Water Sci. 2017, 7, 2267–2277. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.N.; Wang, X.N.; Zhang, F.; Wu, J.; Zhang, W.; Lu, Y.Z.; Fu, L.; Lau, T.C.; Zeng, R.J. High-rate anaerobic decolorization of methyl orange from synthetic azo dye wastewater in a methane-based hollow fiber membrane bioreactor. J. Hazard. Mater. 2020, 388, 121753. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Shi, J.; Xu, C.; Ma, W.; Zhang, Z.; Zhu, H.; Han, H. Ecological and functional research into microbiomes for targeted phenolic removal in anoxic carbon-based fluidized bed reactor (CBFBR) treating coal pyrolysis wastewater (CPW). Bioresour. Technol. 2020, 308, 123308. [Google Scholar] [CrossRef] [PubMed]
- Priyaragini, S.; Veena, S.; Swetha, D.; Karthik, L.; Kumar, G.; Rao, K.B. Evaluating the effectiveness of marine actinobacterial extract and its mediated titanium dioxide nanoparticles in the degradation of azo dyes. J. Environ. Sci. 2014, 26, 775–782. [Google Scholar] [CrossRef]
- Chittal, V.; Gracias, M.; Anu, A.; Saha, P.; Rao, K.B. Biodecolorization and Biodegradation of azo dye reactive orange-16 by marine nocardiopsis sp. Iran. J. Biotechnol. 2019, 17, e1551. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Su, W.; Chen, H.; Barberán, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of actinobacteria and proteobacteria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Samples | Dehydrogenase (ng TPF g−1 Soil h−1) | Urease (mg NH4+-N g−1 Soil h−1) | Cellulase (μg Glucose g−1 Soil h−1) | Acid Phosphomonoesterase (μg Glucose g−1 Soil h−1) |
|---|---|---|---|---|
| DS | 15.37 ± 1.96 | 3.36 ± 0.47 a | 27.28 ± 1.41 | 54.92 ± 9.16 a |
| ALS | 22.35 ± 2.68 | 5.11 ± 0.63 a | 40.75 ± 4.55 | 55.71 ± 11.35 a |
| LSD (p ≤ 0.05) | 9.17 | 1.99 | 10.56 | 37.53 |
| Soil Samples | Number of OTUs | Chao 1 (Richness) | Shanon (Diversity) | Inv-Simpson (Diversity) |
|---|---|---|---|---|
| DS | 8302 | 1303.71 | 5.54 | 51.82 |
| ALS | 12162 | 1472.93 | 6.25 | 158.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnamoorthy, R.; Roy Choudhury, A.; Arul Jose, P.; Suganya, K.; Senthilkumar, M.; Prabhakaran, J.; Gopal, N.O.; Choi, J.; Kim, K.; Anandham, R.; et al. Long-Term Exposure to Azo Dyes from Textile Wastewater Causes the Abundance of Saccharibacteria Population. Appl. Sci. 2021, 11, 379. https://doi.org/10.3390/app11010379
Krishnamoorthy R, Roy Choudhury A, Arul Jose P, Suganya K, Senthilkumar M, Prabhakaran J, Gopal NO, Choi J, Kim K, Anandham R, et al. Long-Term Exposure to Azo Dyes from Textile Wastewater Causes the Abundance of Saccharibacteria Population. Applied Sciences. 2021; 11(1):379. https://doi.org/10.3390/app11010379
Chicago/Turabian StyleKrishnamoorthy, Ramasamy, Aritra Roy Choudhury, Polpass Arul Jose, Kathirvel Suganya, Murugaiyan Senthilkumar, James Prabhakaran, Nellaiappan Olaganathan Gopal, Jeongyun Choi, Kiyoon Kim, Rangasamy Anandham, and et al. 2021. "Long-Term Exposure to Azo Dyes from Textile Wastewater Causes the Abundance of Saccharibacteria Population" Applied Sciences 11, no. 1: 379. https://doi.org/10.3390/app11010379
APA StyleKrishnamoorthy, R., Roy Choudhury, A., Arul Jose, P., Suganya, K., Senthilkumar, M., Prabhakaran, J., Gopal, N. O., Choi, J., Kim, K., Anandham, R., & Sa, T. (2021). Long-Term Exposure to Azo Dyes from Textile Wastewater Causes the Abundance of Saccharibacteria Population. Applied Sciences, 11(1), 379. https://doi.org/10.3390/app11010379