
Not Only Antimicrobial: Metronidazole Mitigates the Virulence of Proteus mirabilis Isolated from Macerated Diabetic Foot Ulcer


 , , , , , ,
, , , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Materials
2.2. Minimum Inhibitory Concentration (MIC) Determination
2.3. Effect of Metronidazole in Sub-MIC on P. mirabilis Growth
2.4. Protease Assay
2.5. Hemolysis Assay
2.6. Urease Assay
2.7. Assay of P. mirabilis Motilities
2.8. Adhesion Assay
2.8.1. Adhesion to Epithelial Cells
2.8.2. Adhesion to Abiotic Surfaces
2.9. Biofilm Formation Assay
2.9.1. Assessment of Biofilm Formation
2.9.2. Biofilm Production Assay
2.9.3. Minimum Biofilm Eradication Concentration (MBEC) Determination
2.10. Combination of Metronidazole with Antibiotics
2.11. Molecular Docking of Metronidazole onto P. mirabilis Proteins
2.12. Statistical Analysis
3. Results
3.1. Determination of MIC and MBEC
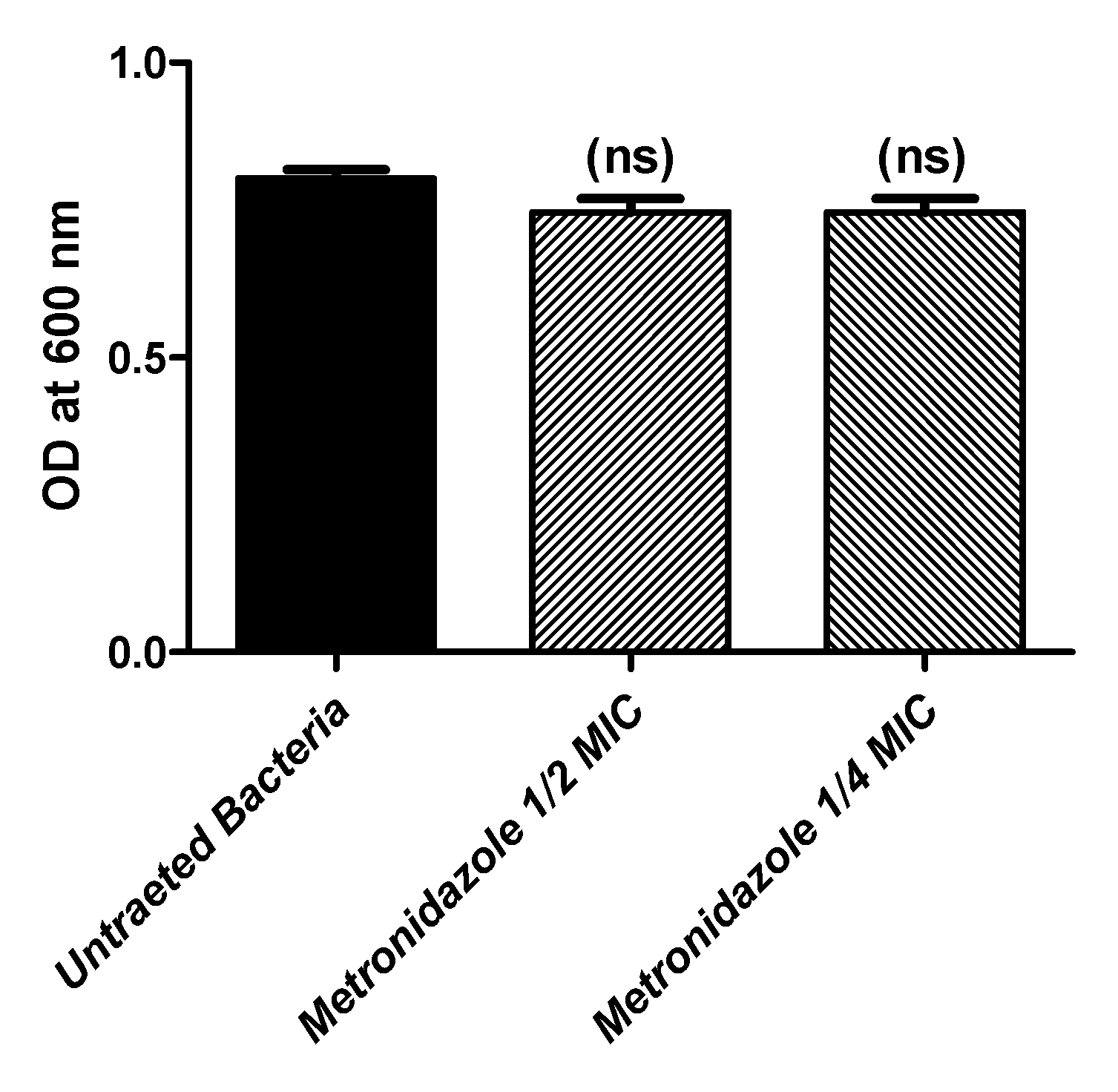
3.2. Effect of Metronidazole in Sub-MIC on Bacterial Growth
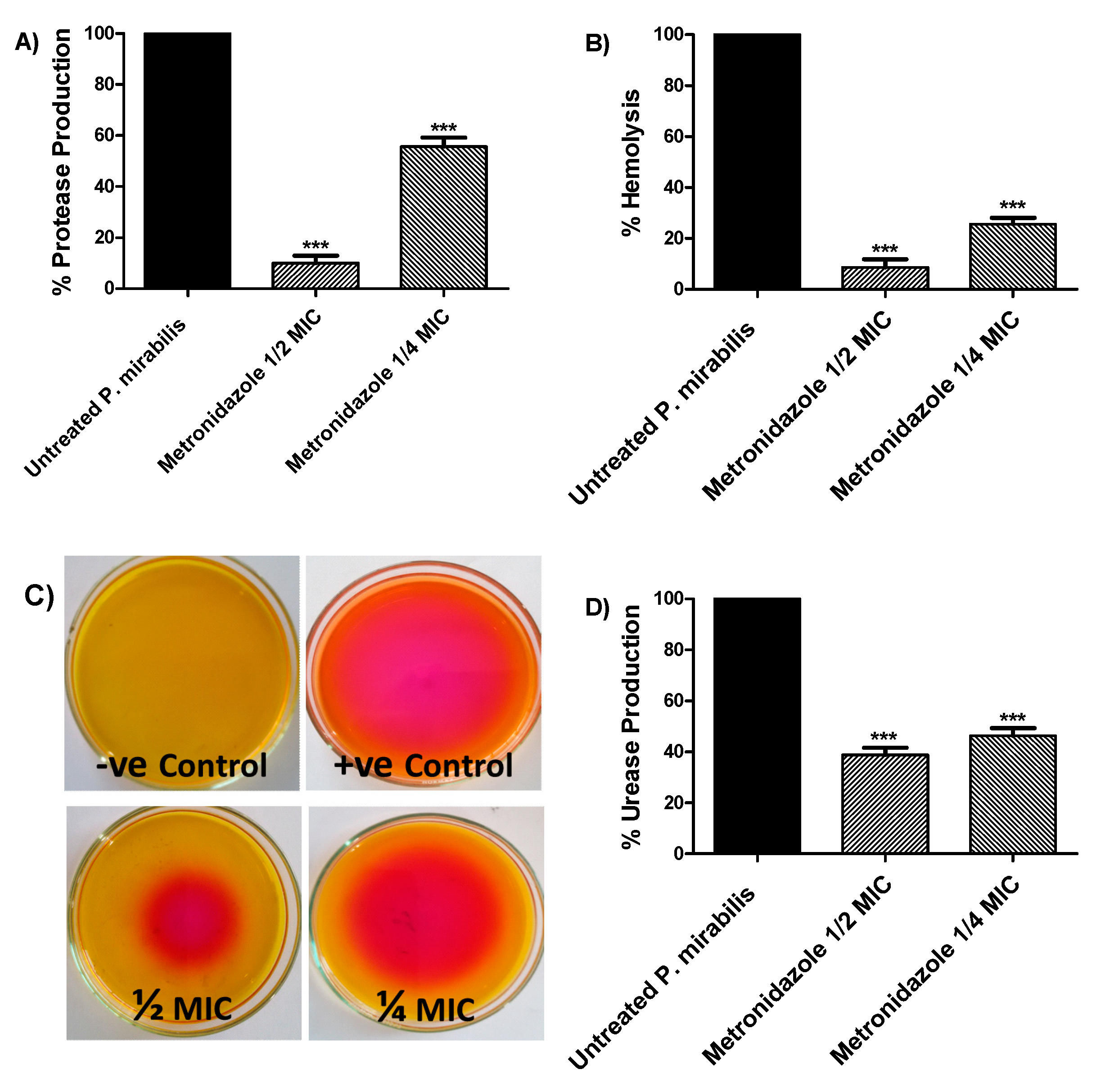
3.3. Effect of Metronidazole in Sub-MIC on Production of Bacterial Virulence Enzymes
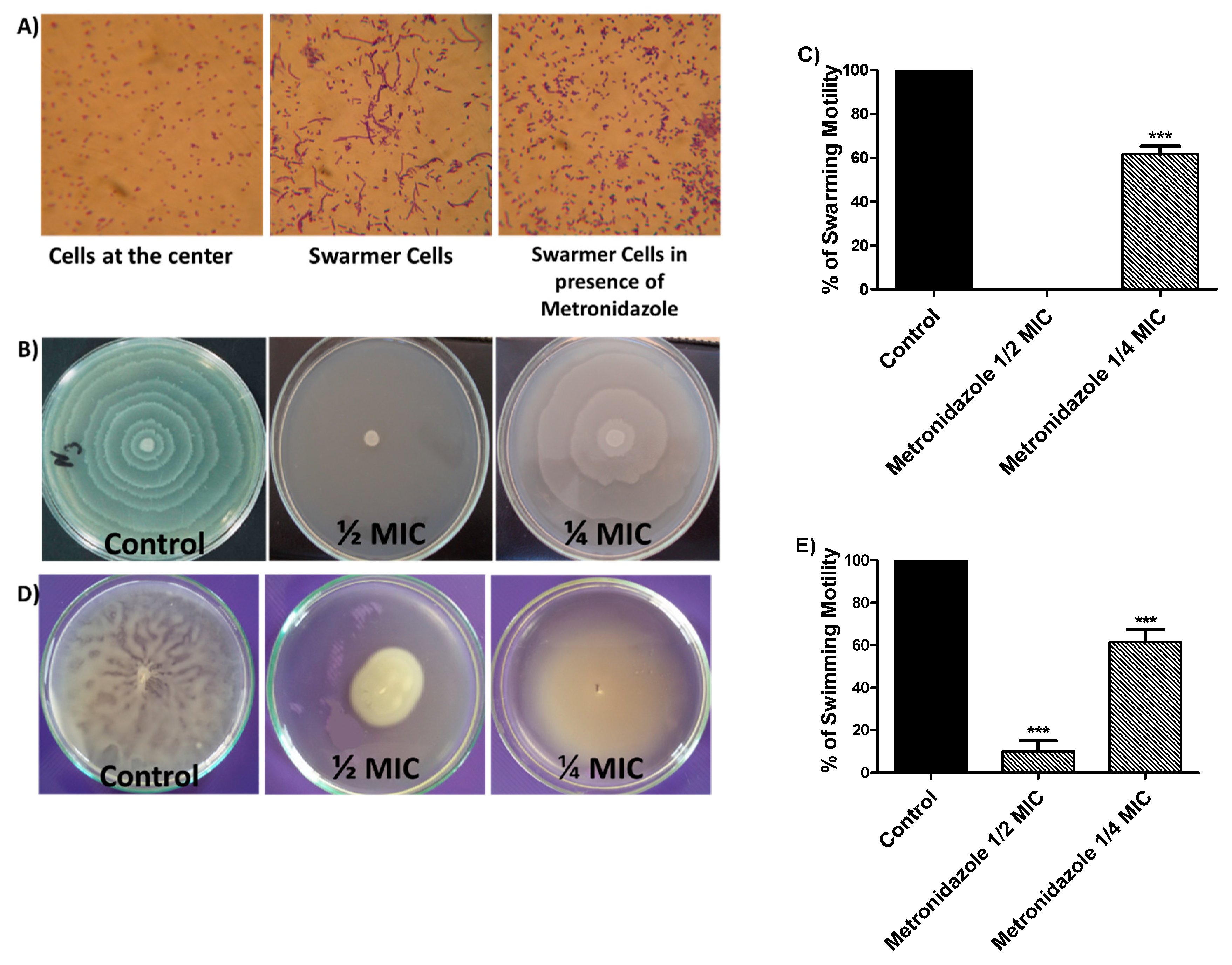
3.4. Effect of Metronidazole in sub-MIC on Bacterial Motilities
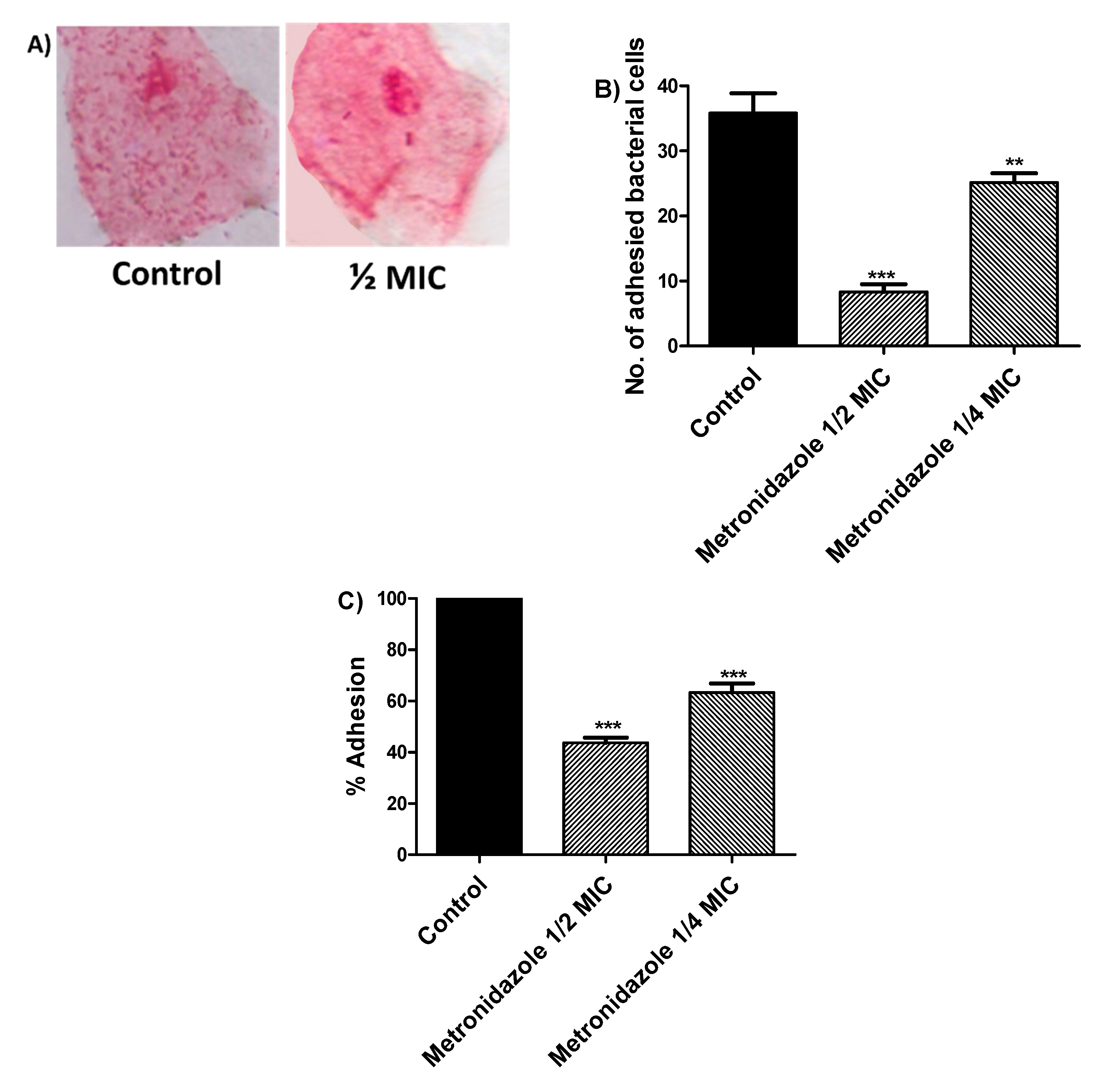
3.5. Effect of Metronidazole in Sub-MIC on Bacterial Adhesion
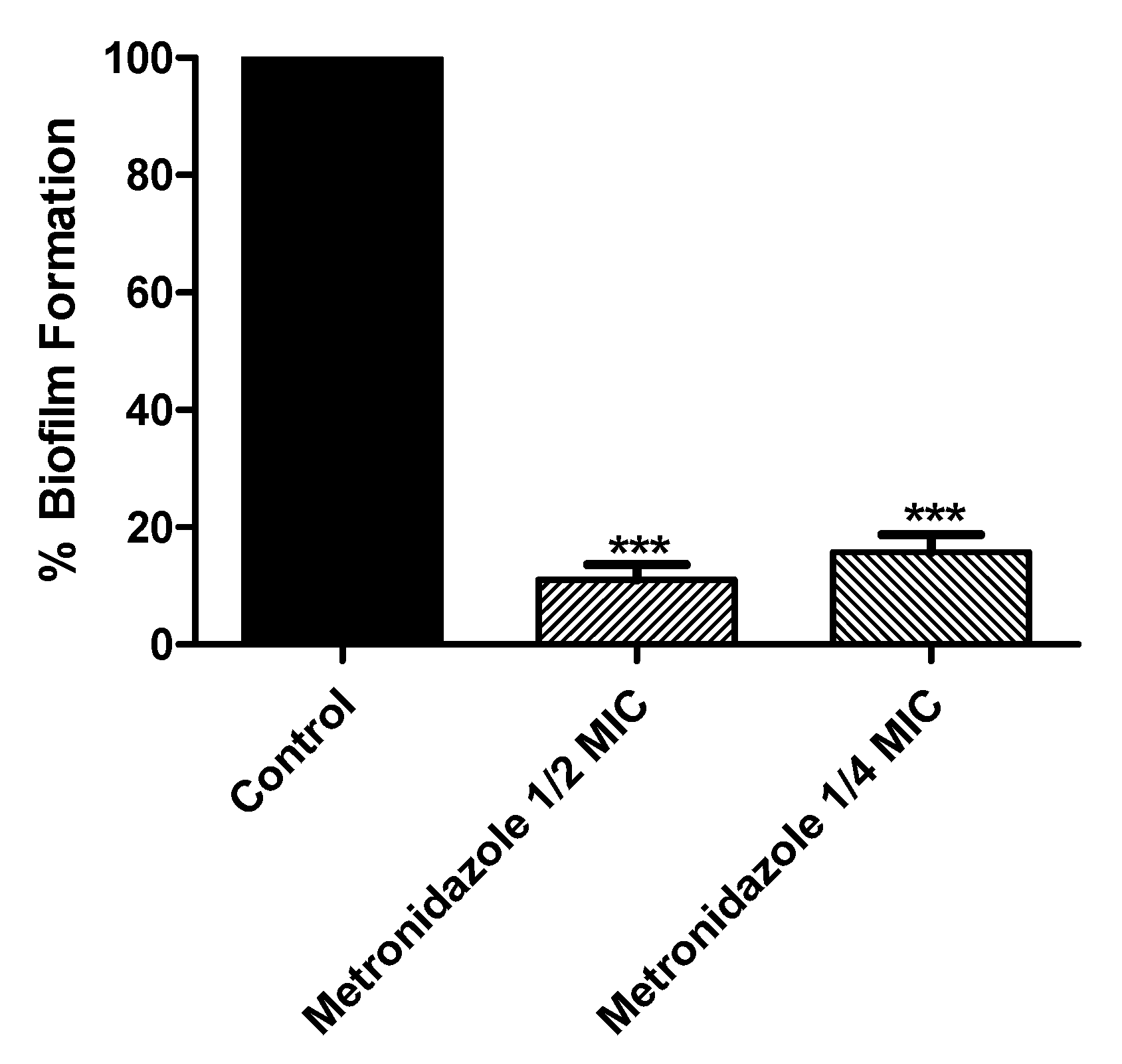
3.6. Effect of Metronidazole in Sub-MIC on Biofilm Formation
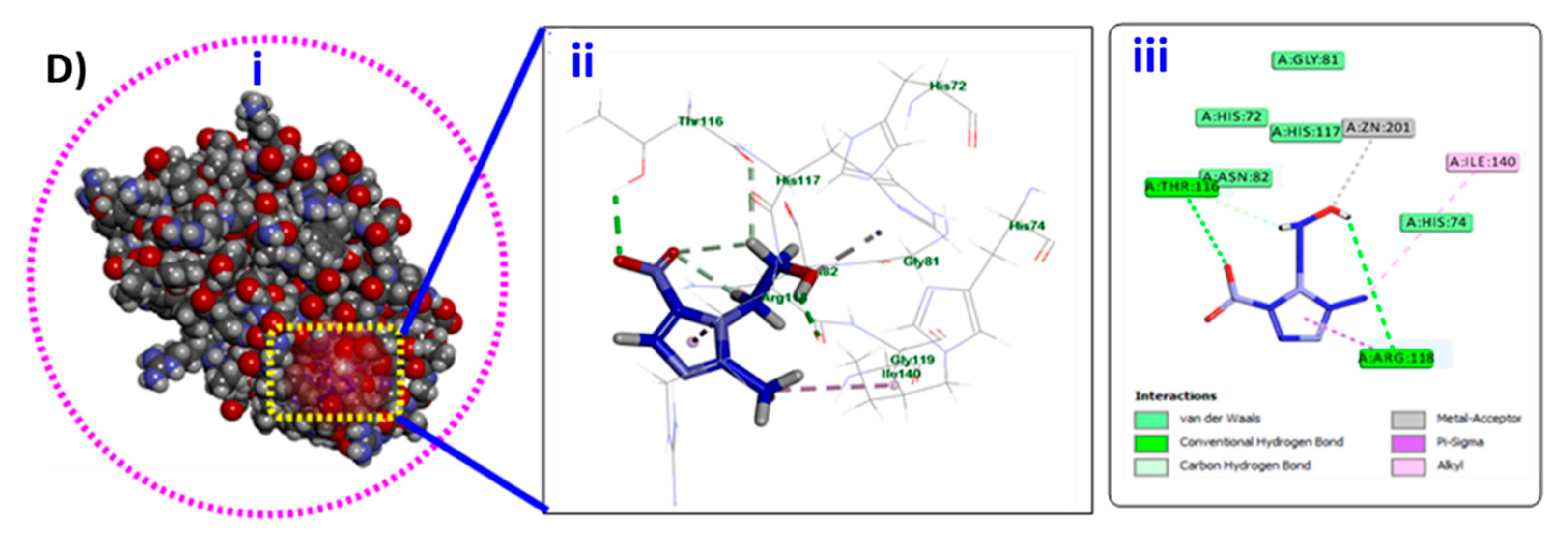
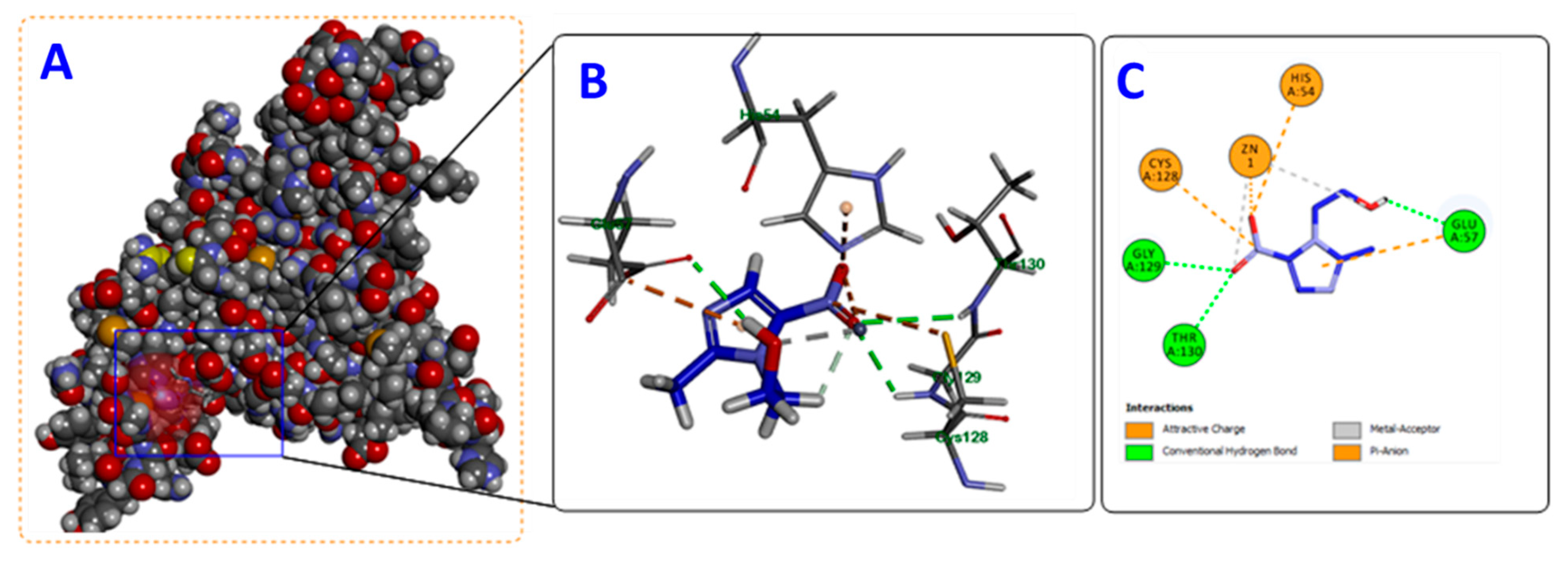
3.7. In-Silico Docking of Metronidazole into P. mirabilis QS Eseential Proteins
3.8. MICs and MBECs of Tested Antibiotics in Precence of Metronidazole in Sub-MIC
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Van Battum, P.; Schaper, N.; Prompers, L.; Apelqvist, J.; Jude, E.; Piaggesi, A.; Bakker, K.; Edmonds, M.; Holstein, P.; Jirkovska, A.; et al. Differences in minor amputation rate in diabetic foot disease throughout Europe are in part explained by differences in disease severity at presentation. Diabet. Med. 2011, 28, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Volmer-Thole, M.; Lobmann, R. Neuropathy and Diabetic Foot Syndrome. Int. J. Mol. Sci. 2016, 17, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulton, A.J. The pathway to foot ulceration in diabetes. Med. Clin. N. Am. 2013, 97, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Hartemann-Heurtier, A.; Senneville, E. Diabetic foot osteomyelitis. Diabetes Metab. 2008, 34, 87–95. [Google Scholar] [CrossRef]
- Hegazy, W.A.H. Hepatitis C virus pathogenesis: Serum IL-33 level indicates liver damage. Afr. J. Microbiol. Res. 2015, 9, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, C.E.; Mobley, H.L. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar] [CrossRef]
- Jacobsen, S.M.; Stickler, D.J.; Mobley, H.L.; Shirtliff, M.E. Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin. Microbiol. Rev. 2008, 21, 26–59. [Google Scholar] [CrossRef] [Green Version]
- Sekhar, S.; Vyas, N.; Unnikrishnan, M.; Rodrigues, G.; Mukhopadhyay, C. Antimicrobial susceptibility pattern in diabetic foot ulcer: A pilot study. Ann. Med. Health Sci. Res. 2014, 4, 742–745. [Google Scholar] [CrossRef] [Green Version]
- Perim, M.C.; Borges Jda, C.; Celeste, S.R.; Orsolin Ede, F.; Mendes, R.R.; Mendes, G.O.; Ferreira, R.L.; Carreiro, S.C.; Pranchevicius, M.C. Aerobic bacterial profile and antibiotic resistance in patients with diabetic foot infections. Rev. Soc. Bras. Med. Trop. 2015, 48, 546–554. [Google Scholar] [CrossRef] [Green Version]
- El-Tahawy, A.T. Bacteriology of diabetic foot. Saudi Med. J. 2000, 21, 344–347. [Google Scholar]
- Flannery, E.L.; Mody, L.; Mobley, H.L. Identification of a modular pathogenicity island that is widespread among urease-producing uropathogens and shares features with a diverse group of mobile elements. Infect. Immun. 2009, 77, 4887–4894. [Google Scholar] [CrossRef] [Green Version]
- Wasfi, R.; Hamed, S.M.; Amer, M.A.; Fahmy, L.I. Proteus mirabilis Biofilm: Development and Therapeutic Strategies. Front. Cell Infect. Microbiol. 2020, 10, 414. [Google Scholar] [CrossRef]
- Tansarli, G.S.; Athanasiou, S.; Falagas, M.E. Evaluation of antimicrobial susceptibility of Enterobacteriaceae causing urinary tract infections in Africa. Antimicrob. Agents Chemother. 2013, 57, 3628–3639. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of CTX-M extended-spectrum beta-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef]
- Abbas, H.A.; Hegazy, W.A.H. Targeting the virulence factors of Serratia marcescens by ambroxol. Roum. Arch. Microbiol. Immunol. 2017, 76, 27–32. [Google Scholar]
- Abbas, H.A.; Hegazy, W.A.H. Repurposing anti-diabetic drug “Sitagliptin” as a novel virulence attenuating agent in Serratia marcescens. PLoS ONE 2020, 15, e0231625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hamid, A.; Marwa, I.; Y El-Naenaeey, E.S.; Hegazy, W.A.; Mosbah, R.A.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Al Saqr, A.; Khafagy, E.S.; Alalaiwe, A.; Aldawsari, M.F.; Alshahrani, S.M.; Anwer, M.K.; Khan, S.; Lila, A.S.A.; Arab, H.H.; Hegazy, W.A.H. Synthesis of Gold Nanoparticles by Using Green Machinery: Characterization and In Vitro Toxicity. Nanomaterials 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Askoura, M.; Hegazy, W.A.H. Ciprofloxacin interferes with Salmonella Typhimurium intracellular survival and host virulence through repression of Salmonella pathogenicity island-2 (SPI-2) genes expression. Pathog. Dis. 2020, 78. [Google Scholar] [CrossRef]
- Hegazy, W.A.H. Diclofenac inhibits virulence of Proteus mirabilis isolated from diabetic foot ulcer. Afr. J. Microbiol. Res. 2016, 10, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Hegazy, W.A.H.; Al Mamari, R.; Almazroui, K.; Al Habsi, A.; Kamona, A.; AlHarthi, H.; Al Lawati, A.I.; AlHusaini, A.H. Retrospective Study of Bone-TB in Oman: 2002-2019. J. Epidemiol. Glob. Health 2021, 11, 238–245. [Google Scholar] [CrossRef]
- Bendary, M.M.; Ibrahim, D.; Mosbah, R.A.; Mosallam, F.; Hegazy, W.A.H.; Awad, N.F.S.; Alshareef, W.A.; Alomar, S.Y.; Zaitone, S.A.; Abd El-Hamid, M.I. Thymol Nanoemulsion: A New Therapeutic Option for Extensively Drug Resistant Foodborne Pathogens. Antibiotics 2020, 10, 25. [Google Scholar] [CrossRef]
- Hegazy, W.A.H.; Khayat, M.T.; Ibrahim, T.S.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Repurposing Anti-diabetic Drugs to Cripple Quorum Sensing in Pseudomonas aeruginosa. Microorganisms 2020, 8, 1285. [Google Scholar] [CrossRef]
- Hegazy, W.A.H.; Khayat, M.T.; Ibrahim, T.S.; Youns, M.; Mosbah, R.; Soliman, W.E. Repurposing of antidiabetics as Serratia marcescens virulence inhibitors. Braz. J. Microbiol. 2021, 52, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Khayyat, A.N.; Hegazy, W.A.H.; Shaldam, M.A.; Mosbah, R.; Almalki, A.J.; Ibrahim, T.S.; Khayat, M.T.; Khafagy, E.S.; Soliman, W.E.; Abbas, H.A. Xylitol Inhibits Growth and Blocks Virulence in Serratia marcescens. Microorganisms 2021, 9, 1083. [Google Scholar] [CrossRef] [PubMed]
- Vishwa, B.; Moin, A.; Gowda, D.V.; Rizvi, S.M.D.; Hegazy, W.A.H.; Abu Lila, A.S.; Khafagy, E.S.; Allam, A.N. Pulmonary Targeting of Inhalable Moxifloxacin Microspheres for Effective Management of Tuberculosis. Pharmaceutics 2021, 13, 79. [Google Scholar] [CrossRef]
- Aldawsari, M.F.; Alalaiwe, A.; Khafagy, E.S.; Al Saqr, A.; Alshahrani, S.M.; Alsulays, B.B.; Alshehri, S.; Abu Lila, A.S.; Danish Rizvi, S.M.; Hegazy, W.A.H. Efficacy of SPG-ODN 1826 Nanovehicles in Inducing M1 Phenotype through TLR-9 Activation in Murine Alveolar J774A.1 Cells: Plausible Nano-Immunotherapy for Lung Carcinoma. Int. J. Mol. Sci. 2021, 22, 6833. [Google Scholar] [CrossRef]
- Freeman, C.D.; Klutman, N.E.; Lamp, K.C. Metronidazole. A therapeutic review and update. Drugs 1997, 54, 679–708. [Google Scholar] [CrossRef] [PubMed]
- Dingsdag, S.A.; Hunter, N. Metronidazole: An update on metabolism, structure-cytotoxicity and resistance mechanisms. J. Antimicrob. Chemother. 2018, 73, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Pearson, M.M. Culture Methods for Proteus mirabilis. Methods Mol. Biol. 2019, 2021, 5–13. [Google Scholar] [CrossRef]
- Nalca, Y.; Jansch, L.; Bredenbruch, F.; Geffers, R.; Buer, J.; Haussler, S. Quorum-sensing antagonistic activities of azithromycin in Pseudomonas aeruginosa PAO1: A global approach. Antimicrob. Agents Chemother. 2006, 50, 1680–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Shuai, Y.; Tao, H.; Li, C.; He, L. Novel Method for the Quantitative Analysis of Protease Activity: The Casein Plate Method and Its Applications. ACS Omega 2021, 6, 3675–3680. [Google Scholar] [CrossRef] [PubMed]
- Vesterlund, S.; Paltta, J.; Karp, M.; Ouwehand, A.C. Measurement of bacterial adhesion-in vitro evaluation of different methods. J. Microbiol. Methods 2005, 60, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Cernohorska, L.; Votava, M. Antibiotic synergy against biofilm-forming Pseudomonas aeruginosa. Folia Microbiol. 2008, 53, 57–60. [Google Scholar] [CrossRef]
- Mackay, M.L.; Milne, K.; Gould, I.M. Comparison of methods for assessing synergic antibiotic interactions. Int. J. Antimicrob. Agents 2000, 15, 125–129. [Google Scholar] [CrossRef]
- Abou-Zied, H.A.; Youssif, B.G.M.; Mohamed, M.F.A.; Hayallah, A.M.; Abdel-Aziz, M. EGFR inhibitors and apoptotic inducers: Design, synthesis, anticancer activity and docking studies of novel xanthine derivatives carrying chalcone moiety as hybrid molecules. Bioorg. Chem. 2019, 89, 102997. [Google Scholar] [CrossRef]
- Al-Sanea, M.M.; Gotina, L.; Mohamed, M.F.; Grace Thomas Parambi, D.; Gomaa, H.A.M.; Mathew, B.; Youssif, B.G.M.; Alharbi, K.S.; Elsayed, Z.M.; Abdelgawad, M.A.; et al. Design, Synthesis and Biological Evaluation of New HDAC1 and HDAC2 Inhibitors Endowed with Ligustrazine as a Novel Cap Moiety. Drug Des. Dev. 2020, 14, 497–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssif, B.G.M.; Mohamed, M.F.A.; Al-Sanea, M.M.; Moustafa, A.H.; Abdelhamid, A.A.; Gomaa, H.A.M. Novel aryl carboximidamide and 3-aryl-1,2,4-oxadiazole analogues of naproxen as dual selective COX-2/15-LOX inhibitors: Design, synthesis and docking studies. Bioorg. Chem. 2019, 85, 577–584. [Google Scholar] [CrossRef]
- Morgenstein, R.M.; Szostek, B.; Rather, P.N. Regulation of gene expression during swarmer cell differentiation in Proteus mirabilis. FEMS Microbiol. Rev. 2010, 34, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, H.A. Inhibition of Virulence of Pseudomonas aeruginosa: A Novel Role of Metronidazole Against Aerobic Bacteria. Res. J. Pharm. Technol. 2015, 8, 1640–1644. [Google Scholar] [CrossRef]
- Saleh, M.M.; Abbas, H.A.; Askoura, M.M. Repositioning secnidazole as a novel virulence factors attenuating agent in Pseudomonas aeruginosa. Microb. Pathog. 2019, 127, 31–38. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus mirabilis Infection. EcoSal Plus 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozalski, A.; Sidorczyk, Z.; Kotelko, K. Potential virulence factors of Proteus bacilli. Microbiol. Mol. Biol. Rev. 1997, 61, 65–89. [Google Scholar] [CrossRef] [PubMed]
- Mobley, H.L.; Island, M.D.; Hausinger, R.P. Molecular biology of microbial ureases. Microbiol. Rev. 1995, 59, 451–480. [Google Scholar] [CrossRef] [PubMed]
- Senior, B.W. The production of HlyA toxin by Proteus penneri strains. J. Med. Microbiol. 1993, 39, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Pearson, M.M.; Sebaihia, M.; Churcher, C.; Quail, M.A.; Seshasayee, A.S.; Luscombe, N.M.; Abdellah, Z.; Arrosmith, C.; Atkin, B.; Chillingworth, T.; et al. Complete genome sequence of uropathogenic Proteus mirabilis, a master of both adherence and motility. J. Bacteriol. 2008, 190, 4027–4037. [Google Scholar] [CrossRef] [Green Version]
- Rather, P.N. Swarmer cell differentiation in Proteus mirabilis. Environ. Microbiol. 2005, 7, 1065–1073. [Google Scholar] [CrossRef]
- Allison, C.; Emody, L.; Coleman, N.; Hughes, C. The role of swarm cell differentiation and multicellular migration in the uropathogenicity of Proteus mirabilis. J. Infect. Dis. 1994, 169, 1155–1158. [Google Scholar] [CrossRef]
- De Kievit, T.R.; Parkins, M.D.; Gillis, R.J.; Srikumar, R.; Ceri, H.; Poole, K.; Iglewski, B.H.; Storey, D.G. Multidrug efflux pumps: Expression patterns and contribution to antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2001, 45, 1761–1770. [Google Scholar] [CrossRef] [Green Version]
- Askoura, M.; Youns, M.; Halim Hegazy, W.A. Investigating the influence of iron on Campylobacter jejuni transcriptome in response to acid stress. Microb. Pathog. 2020, 138, 103777. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Suarez, C.; Pasma, J.; van der Borden, A.J.; Wingender, J.; Flemming, H.C.; Busscher, H.J.; van der Mei, H.C. Influence of extracellular polymeric substances on deposition and redeposition of Pseudomonas aeruginosa to surfaces. Microbiology 2002, 148, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Belas, R.; Suvanasuthi, R. The ability of Proteus mirabilis to sense surfaces and regulate virulence gene expression involves FliL, a flagellar basal body protein. J. Bacteriol. 2005, 187, 6789–6803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, W.A.H.; Abbas, H.A. Evaluation of the role of SsaV ‘Salmonella pathogenicity island-2 dependent type III secretion system components on the virulence behavior of Salmonella enterica serovar Typhimurium. Afr. J. Biotechnol. 2017, 16, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, J.N.; Norsworthy, A.N.; Sun, T.T.; Pearson, M.M. Proteus mirabilis fimbriae- and urease-dependent clusters assemble in an extracellular niche to initiate bladder stone formation. Proc. Natl. Acad. Sci. USA 2016, 113, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Ubhayasekera, W.; Breed, M.C.; Norsworthy, A.N.; Serr, N.; Mobley, H.L.T.; Pearson, M.M.; Knight, S.D. MrpH, a new class of metal-binding adhesin, requires zinc to mediate biofilm formation. PLoS Pathog. 2020, 16, e1008707. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Ashraf, M.I.; Mujawar, S.; Mishra, R.; Lahiri, C. In silico Identification of the Indispensable Quorum Sensing Proteins of Multidrug Resistant Proteus mirabilis. Front. Cell Infect. Microbiol. 2018, 8, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.C.; Chien, H.F.; Tsai, Y.L.; Liu, M.C.; Liaw, S.J. The RNA chaperone Hfq is involved in stress tolerance and virulence in uropathogenic Proteus mirabilis. PLoS ONE 2014, 9, e85626. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Agent | MIC | MBEC | Ratio MBEC/MIC |
|---|---|---|---|
| Metronidazole | 10 mg/mL | 40 mg/mL | 4 |
| Ciprofloxacin | 2 µg/mL | 128 µg/mL | 64 |
| Cefoperazone | 64 µg/mL | 1024 µg/mL | 16 |
| Amoxicillin/Clavulinic acid | 256 µg/mL | 2048 µg/mL | 8 |
| Imipenem | 4 µg/mL | 8 µg/mL | 2 |
| Gentamycin | 16 µg/mL | 512 µg/mL | 32 |
| Tetracycline | 64 µg/mL | 2048 µg/mL | 32 |
| Chloramphenicol | 64 µg/mL | 2048 µg/mL | 32 |
| Antibiotic | MIC | MICmet | FIC | MBEC | MBECmet | FIC |
|---|---|---|---|---|---|---|
| Ciprofloxacin | 2 µg/mL | 1 µg/mL | 0.5 | 128 µg/mL | 32 µg/mL | 0.25 |
| Cefoperazone | 64 µg/mL | 16 µg/mL | 0.25 | 1024 µg/mL | 256 µg/mL | 0.25 |
| Amoxicillin/Clavulinic acid | 256 µg/mL | 32 µg/mL | 0.125 | 2048 µg/mL | 512 µg/mL | 0.25 |
| Imipenem | 4 µg/mL | 2 µg/mL | 0.5 | 8 µg/mL | 4 µg/mL | 0.5 |
| Gentamycin | 16 µg/mL | 16 µg/mL | 1 | 512 µg/mL | 256 µg/mL | 0.5 |
| Tetracycline | 64 µg/mL | 64 µg/mL | 1 | 2048 µg/mL | 1024 µg/mL | 0.5 |
| Chloramphenicol | 64 µg/mL | 16 µg/mL | 0.25 | 2048 µg/mL | 256 µg/mL | 0.125 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khayyat, A.N.; Abbas, H.A.; Mohamed, M.F.A.; Asfour, H.Z.; Khayat, M.T.; Ibrahim, T.S.; Youns, M.; Khafagy, E.-S.; Abu Lila, A.S.; Safo, M.K.; et al. Not Only Antimicrobial: Metronidazole Mitigates the Virulence of Proteus mirabilis Isolated from Macerated Diabetic Foot Ulcer. Appl. Sci. 2021, 11, 6847. https://doi.org/10.3390/app11156847
Khayyat AN, Abbas HA, Mohamed MFA, Asfour HZ, Khayat MT, Ibrahim TS, Youns M, Khafagy E-S, Abu Lila AS, Safo MK, et al. Not Only Antimicrobial: Metronidazole Mitigates the Virulence of Proteus mirabilis Isolated from Macerated Diabetic Foot Ulcer. Applied Sciences. 2021; 11(15):6847. https://doi.org/10.3390/app11156847
Chicago/Turabian StyleKhayyat, Ahdab N., Hisham A. Abbas, Mamdouh F. A. Mohamed, Hani Z. Asfour, Maan T. Khayat, Tarek S. Ibrahim, Mahmoud Youns, El-Sayed Khafagy, Amr S. Abu Lila, Martin K. Safo, and et al. 2021. "Not Only Antimicrobial: Metronidazole Mitigates the Virulence of Proteus mirabilis Isolated from Macerated Diabetic Foot Ulcer" Applied Sciences 11, no. 15: 6847. https://doi.org/10.3390/app11156847
APA StyleKhayyat, A. N., Abbas, H. A., Mohamed, M. F. A., Asfour, H. Z., Khayat, M. T., Ibrahim, T. S., Youns, M., Khafagy, E. -S., Abu Lila, A. S., Safo, M. K., & Hegazy, W. A. H. (2021). Not Only Antimicrobial: Metronidazole Mitigates the Virulence of Proteus mirabilis Isolated from Macerated Diabetic Foot Ulcer. Applied Sciences, 11(15), 6847. https://doi.org/10.3390/app11156847