
Functional and Bioactive Properties of Protein Extracts Generated from Spirulina platensis and Isochrysis galbana T-Iso

Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Algae Used in This Study
2.2. Protein Extraction and Determination
2.3. Total Amino Acid (TAA) and Free Amino Acids (FAAs) Composition
2.4. Protein Size Composition and Polyacrylamide Gel Electrophoresis
2.5. Determination of the pH of Protein Extracts
2.6. Water Activity (aw)
2.7. Water-Holding Capacity (WHC) and Oil-Holding Capacity (OHC) of Protein Extracts
2.8. Solubility
2.9. Foaming Capacity and Stability
2.10. Emulsifying Activity and Stability (EAI and ESI) of Protein Extracts
2.11. ACE-I and Renin Inhibitory Activity of Extracted Soluble Proteins
2.12. Statistical Analysis
3. Results and Discussion
3.1. Protein Yield and Composition
3.2. Amino Acid Composition
3.3. Determination of pH and Water Activity of Protein Extracts
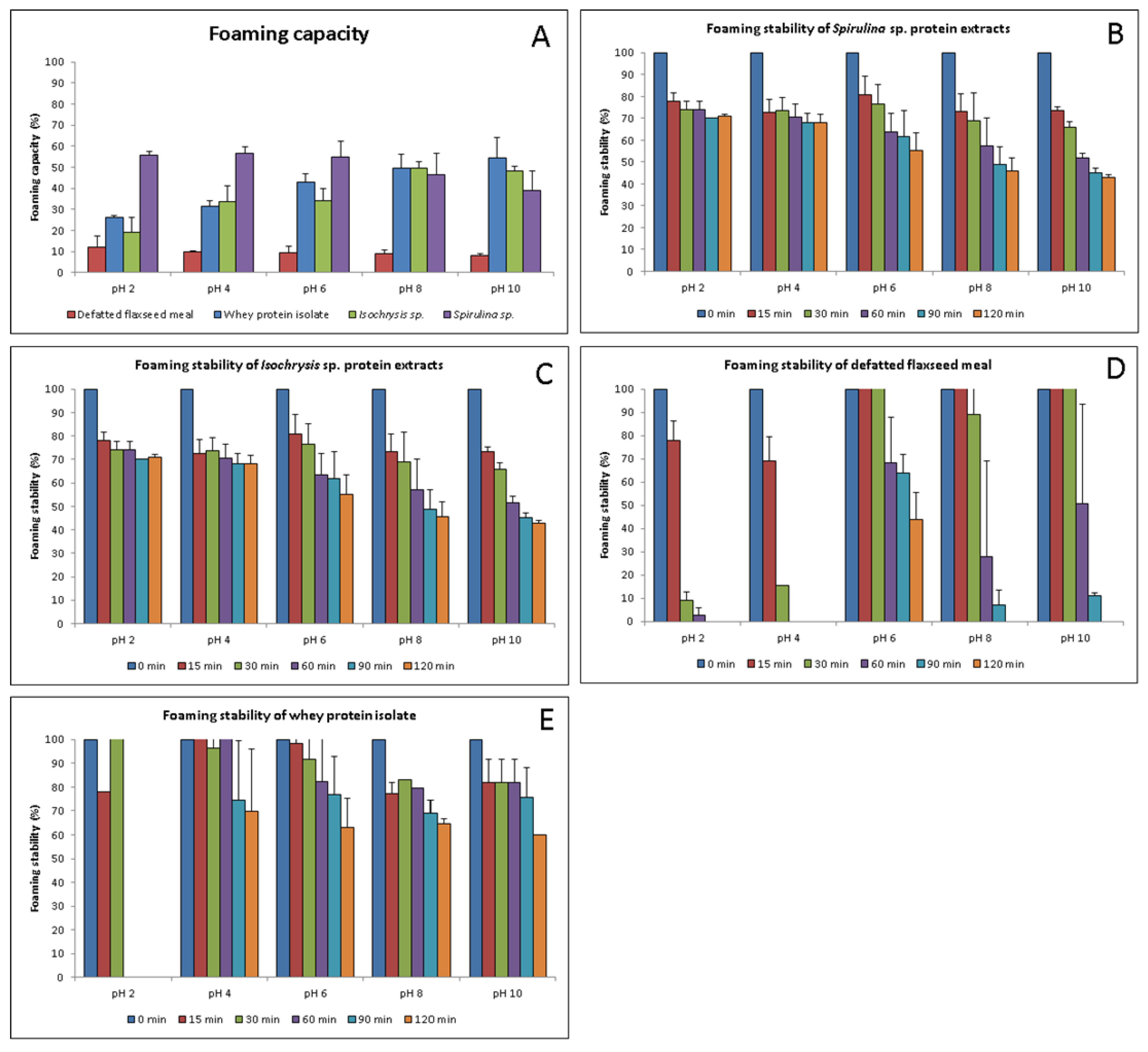
3.4. Foaming Capacity and Stability of Protein Extracts
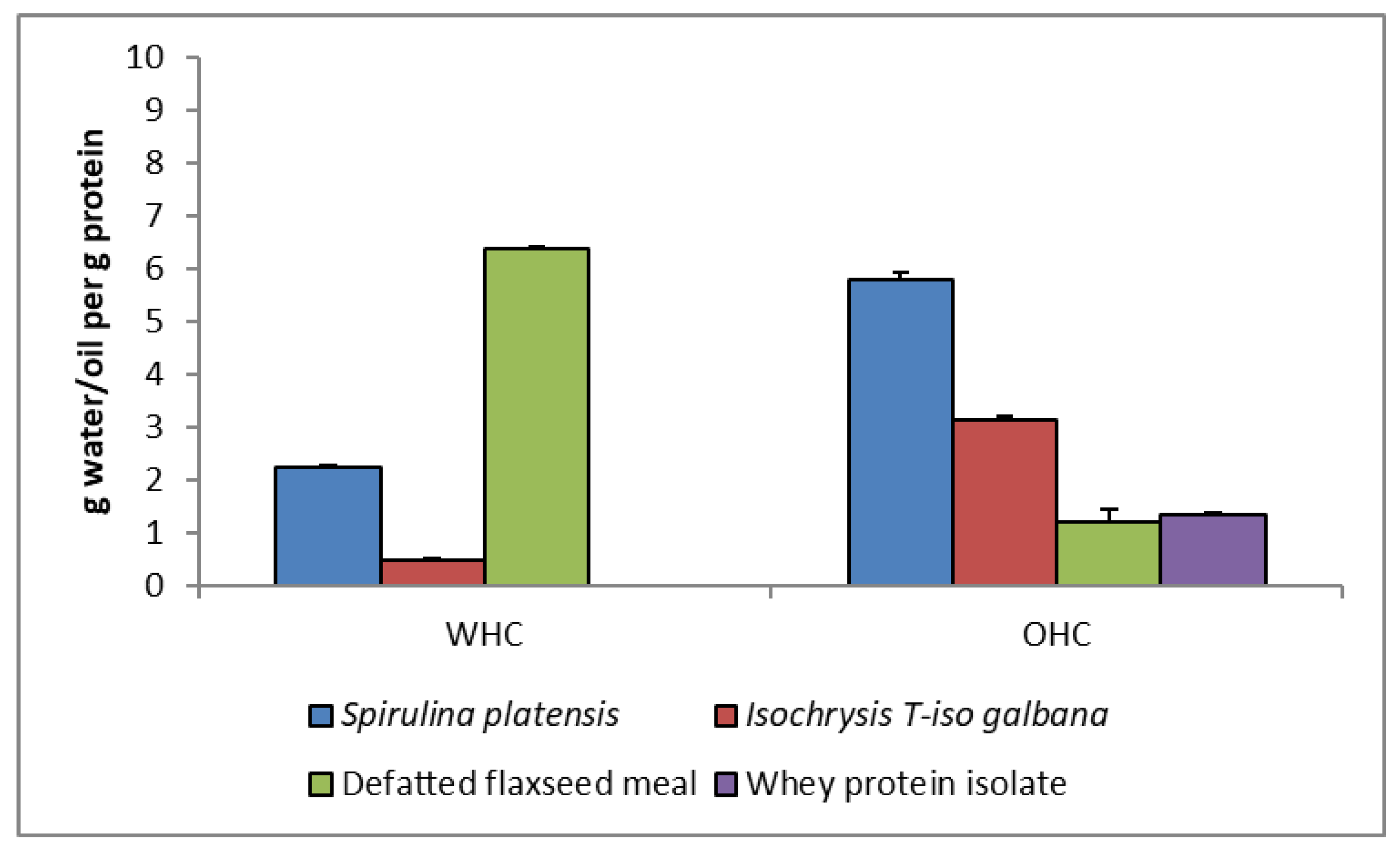
3.5. The WHC and OHC of Protein Extracts
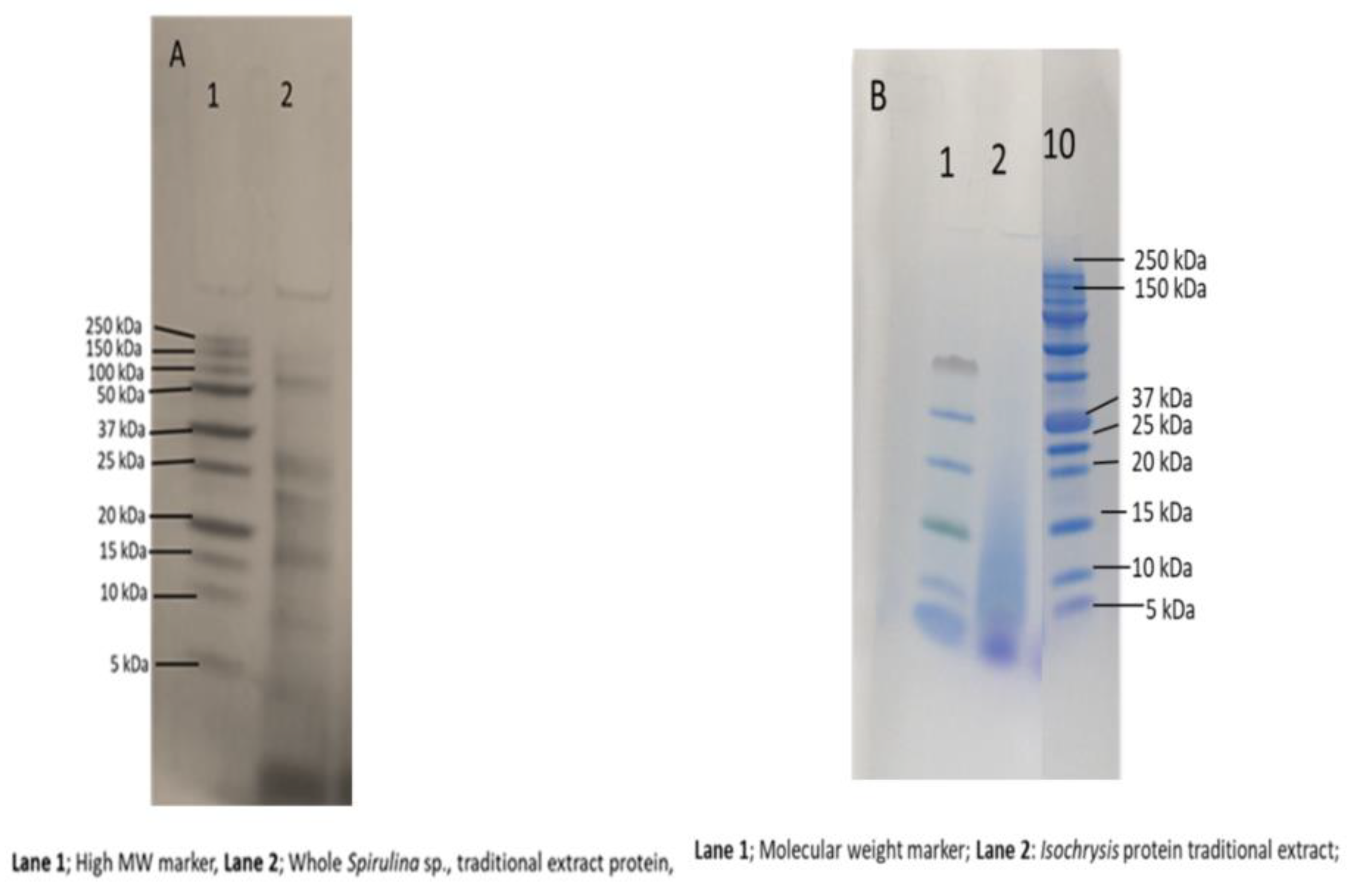
3.6. Molecular Weight Distribution Profile of Soluble Proteins Only from Spirulina sp. and Isochrysis sp.
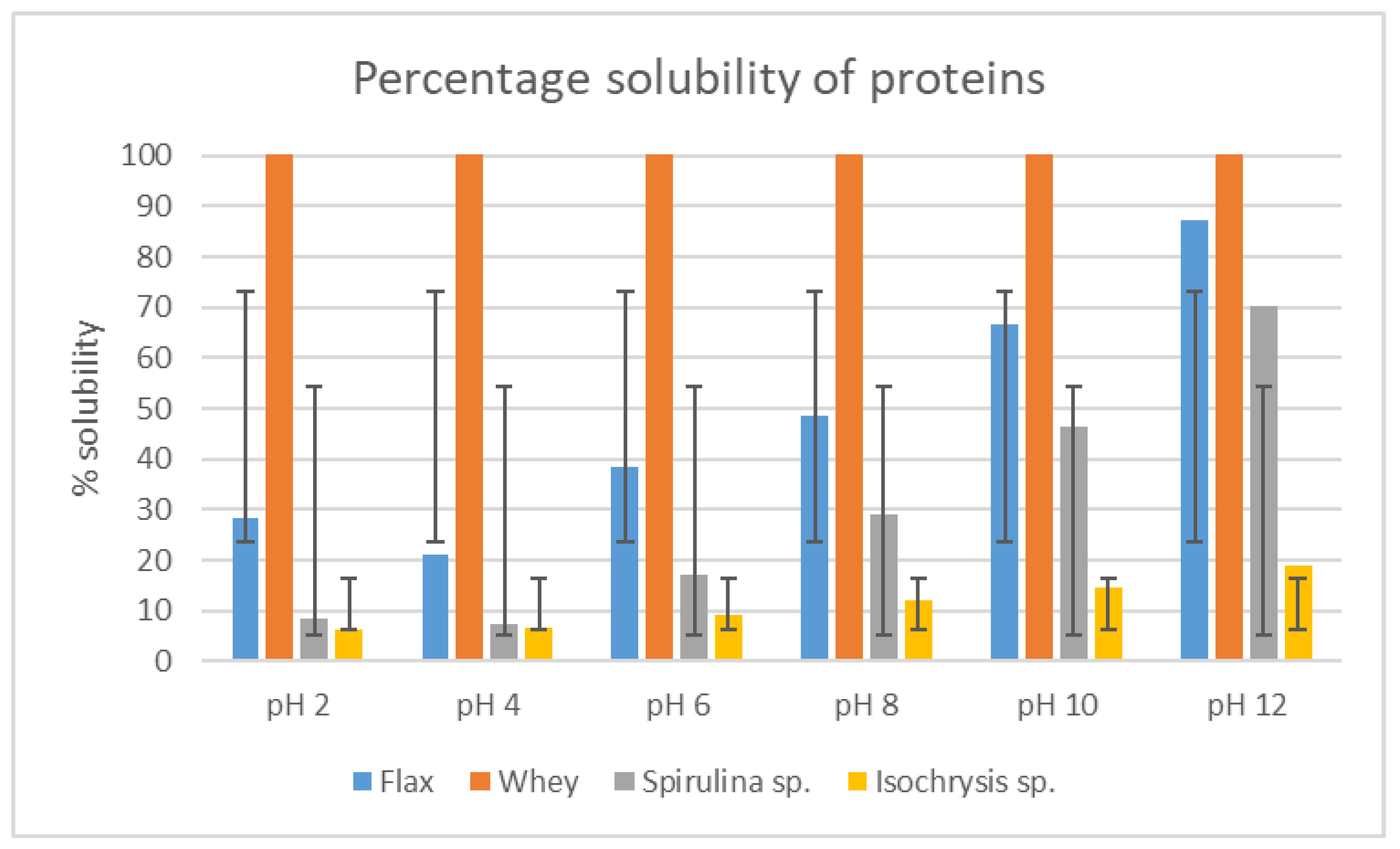
3.7. Solubility of Protein Extracts
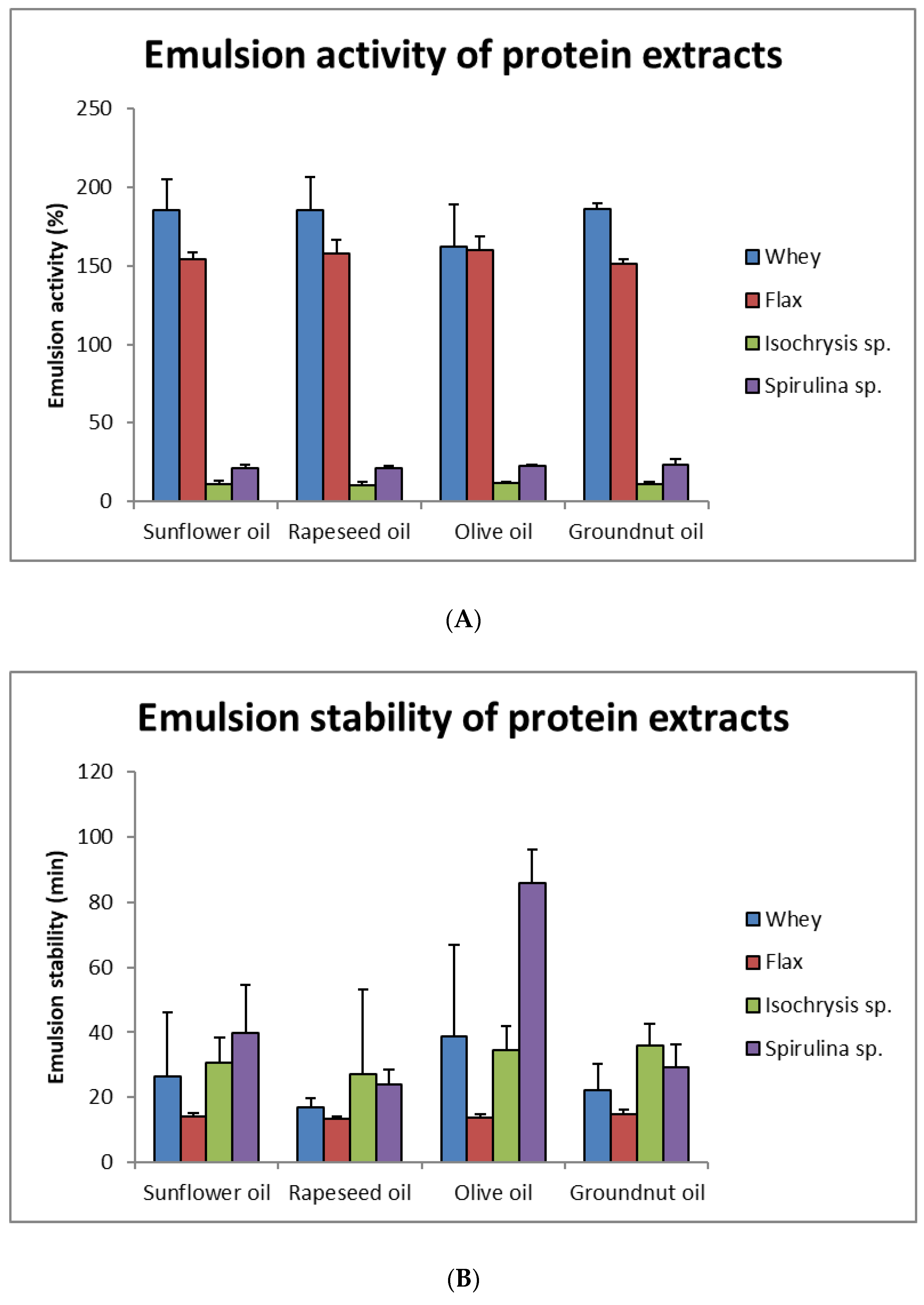
3.8. Emulsion Activity and Stability of Protein Extracts
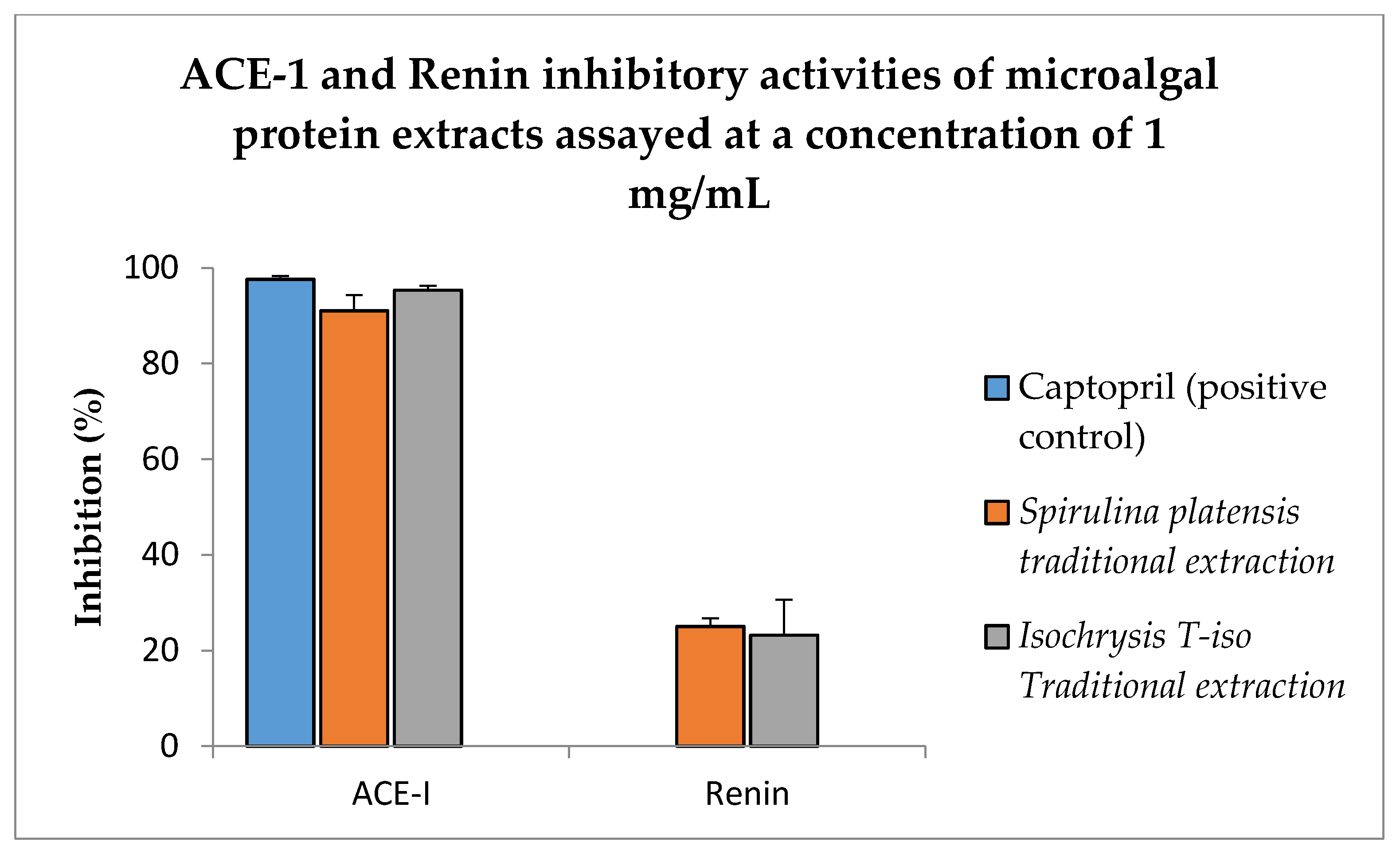
3.9. ACE-I and Renin Inhibitory Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moorehead, K.; Campelli, B. Spirulina Nature’s Superfood, 3rd ed.; Cyanotech Corporation: Kalulua-Kona, HI, USA, 2011. [Google Scholar]
- Dörner, J.; Carbonell, P.; Pino, S.; Farías, A. Variation of Fatty Acids in Isochrysis galbana (T-Iso) and Tetraselmis suecica, cultured under different Nitrate availabilities. Fish Aquac. J. 2014, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Barka, A.; Blecker, C. Microalgae as a potential source of single cell proteins. A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 427436. [Google Scholar]
- Gouveia, L.; Batista, A.P.; Sousa, I.; Raymundo, A.; Bandarra, N.M. Microalgae in novel food products. In Food Chemistry Research Developments; Papadopoulos, K.N., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2008; pp. 75–111. [Google Scholar]
- Piwowar, A.; Harasym, A. The importance and prospects of the use of algae in Agribusiness. Sustainability 2020, 12, 5669. [Google Scholar] [CrossRef]
- Sankaran, R.; Manickam, S.; Yap, Y.J.; Ling, T.C.; Chang, J.-S.; Show, P.L. Extraction of proteins from microalgae using integrated method of sugaring-out assisted liquid biphasic flotation (LBF) and ultrasound. Ultrason. Sonochem. 2018, 48, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Gerde, J.A.; Montalbo-Lomboy, M.; Yao, L.; Grewell, D.; Wang, T. Evaluation of microalgae cell disruption by ultrasonic treatment. Bioresour. Technol. 2012, 125, 175–181. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Ursu, A.V.; Laroche, C.; Zebib, B.; Pontalier, P.-Y.; Vaca-Garcia, C. Release of hydro-soluble microalgal proteins using mechanical and chemical treatments. Algal Res. 2014, 3, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vaquero, M.; Lopez-Alonso, M.; Hayes, M. Assessment of the functional properties of protein extracted from the brown seaweed Himanthalia elongata (Linnaeus) S. F. Gray. Food Res. Int. 2017, 99, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Hamed, I. The Evolution and Versatility of Microalgal Biotechnology: A Review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef] [PubMed]
- Galland-Irmouli, A.-V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.-P.; Villaume, C.; Guéant, J.-L. Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Mora, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Vila, A.; Hayes, M. Isolation and characterisation of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 10. [Google Scholar] [CrossRef]
- Dave, L.A.; Hayes, M.; Mora, L.; Montoya, C.A.; Moughan, P.J.; Rutherfurd, S.M. Gastrointestinal Endogenous Protein-Derived Bioactive Peptides: An in Vitro Study of Their Gut Modulatory Potential. Int. J. Mol. Sci. 2016, 17, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonne, A.H.; Simonne, E.H.; Eitenmiller, R.R.; Mills, H.A.; Cresman, C.P. Could the Dumas Method Replace the Kjeldahl Digestion for Nitrogen and Crude Protein Determinations in Foods? J. Sci. Food Agric. 1997, 73, 39–45. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae. Calculation of new N-P conversion factors. Eur. J. Phycol. 2002, 37, 758–811. [Google Scholar]
- Hill, R.L. Hydrolysis of proteins. Adv. Protein Chem. 1965, 20, 37. [Google Scholar]
- Bencini, M.C. Functional Properties of Drum-Dried Chickpea (Cicer arietinum L.) Flours. J. Food Sci. 1986, 51, 1518–1521. [Google Scholar] [CrossRef]
- Beuchat, L.R.; Cherry, J.P.; Quinn, M.R. Physicochemical properties of peanut flour as affected by proteolysis. J. Agric. Food Chem. 1975, 23, 616–620. [Google Scholar] [CrossRef]
- Poole, S.; West, S.I.; Walters, C.L. Protein–protein interactions: Their importance in the foaming of heterogeneous protein systems. J. Sci. Food Agric. 1984, 35, 701–711. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar] [CrossRef]
- Ufaz, S.; Galilli, G. Improving the content of essential amino acids in crop plants: Goals and opportunities. Plant Physiol. 2008, 147, 954–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein Determination—Method Matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, M. Measuring Protein Content in Food: An Overview of Methods. Foods 2020, 9, 1340. [Google Scholar] [CrossRef]
- Sablani, S.; Kasapis, S.; Rahman, M. Evaluating water activity and glass transition concepts for food stability. J. Food Eng. 2007, 78, 266–271. [Google Scholar] [CrossRef]
- Liestianty, D.; Rodianawati, I.; Arfah, R.A.; Assa, A. Nutritional analysis of Spirulina sp to promote as superfood candidate. IOP Conf. Ser. Mater. Sci. Eng. 2015, 509, 012031. [Google Scholar] [CrossRef]
- Schmidt, S.J.; Fontana, A.J. Appendix E: Water activity values of select food ingredients and products. In Water Activity in Foods; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2008; pp. 407–420. [Google Scholar]
- Zayas, J. Foaming properties of proteins. In Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 1997; pp. 132–227. [Google Scholar]
- Ngoc, T.T.B.; Len, N.T.; Lindberg, J.E. Chemical characterisation and water holding capacity of Fibre-rich feedstuffs used for pigs in Vietnam. Asian Australas. J. Anim. Sci. 2012, 25, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Sun, D.-W. Factors affecting the water holding capacity of red meat products: A review of recent research advances. Crit. Rev. Food Sci. Nutr. 2008, 48, 2. [Google Scholar] [CrossRef] [PubMed]
- De Moor, H.; Huyghebaert, A. Functional properties of dehydrated protein-rich milk products. In Physico-Chemical Aspects of Dehydrated Protein-Rich Milk Products; Statens Forsoegsmejeri: Helsingor, Denmark, 1983. [Google Scholar]
- Kumar, K.S.; Ganesan, K.; Selvaraj, K.; Rao, P.S. Studies on the functional properties of protein concentrate of Kappaphycus alvarezii (Doty) Doty—An edible seaweed. Food Chem. 2014, 153, 353–360. [Google Scholar] [CrossRef]
- Seena, S.; Sridhar, K. Physicochemical, functional and cooking properties of under explored legumes, Canavalia of the southwest coast of India. Food Res. Int. 2005, 38, 803–814. [Google Scholar] [CrossRef]
- Chandi, G.K.; Sogi, D. Functional properties of rich bran protein concentrates. J. Food Eng. 2007, 79, 592–597. [Google Scholar] [CrossRef]
- Gregarsen, S.; Pertseva, M.; Marcatili, P.; Holdt, S.L.; Jacobsen, C.; García-Moreno, P.J.; Hansen, E.B.; Overgaard, M.T. Proteomic characterization of pilot scale hot-water extracts from the industrial carrageenan red seaweed Eucheuma denticulatum. bioRxiv 2020. [Google Scholar] [CrossRef]
- Brandelli, A.; Daroit, D.J.; Corrêa, A.P.F. Whey as a source of peptides with remarkable biological activities. Food Res. Int. 2015, 73, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Hayes, M.; Tiwari, B.K. Bioactive Carbohydrates and Peptides in Foods: An Overview of Sources, Downstream Processing Steps and Associated Bioactivities. Int. J. Mol. Sci. 2015, 16, 22485–22508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Xu, N.; Sun, X.; Yu, H.; Zhou, C. Hydrolysis and purification of ACE inhibitory peptides from the marine microalga Isochrysis galbana. J. Appl. Phycol. 2014, 27, 351–361. [Google Scholar] [CrossRef]
- European Union. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, 327, 1–22. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spirulina sp. | Isochrysis sp. | ||||
|---|---|---|---|---|---|
| Spirulina platensis Whole Biomass (g/kg) | Spirulina platensis Protein Extract (g/kg) | Isochrysis T-Iso Whole Biomass (g/ kg) | Isochrysis T-Iso Protein Extract (g/kg) | ||
| Essential amino acids | Isoleucine | 3.53 | 4.61 | 2.75 | 0.3 |
| Leucine | 5.59 | 7.58 | 5.39 | 0.45 | |
| Valine | 4.54 | 5.46 | 3.64 | 0.4 | |
| Phenylanine | 2.95 | 4.38 | 3.07 | 0.3 | |
| Tyrptophan | . | . | . | . | |
| Histidine | 1.48 | 1.82 | 2.53 | 0.59 | |
| Lysine | 2.78 | 2.64 | 2.8 | 0.27 | |
| Threonine | 3.38 | 3.58 | 3.13 | 0.31 | |
| Methionine | 1.84 | 1.9 | 1.95 | 0.17 | |
| Non-essential amino acids | Alanine | 5.38 | 5 | 4.88 | 0.54 |
| Glycine | 3.28 | 4.11 | 3.46 | 0.36 | |
| Proline | 2.35 | 3.38 | 3.17 | . | |
| Tyrosine | 2.41 | 2.75 | 0.98 | 0.38 | |
| Aspartic acid and Asparagine | 6.55 | 7.18 | 5.92 | 0.72 | |
| Glutamic acid and Glutamine | 8.99 | 7.65 | 6.43 | 0.73 | |
| Arginine | 4.09 | 4.31 | 3.61 | 0.38 | |
| Serine | 3.06 | 3.63 | 2.72 | 0.27 | |
| Cysteine | 1.56 | 0.69 | 1.55 | 0.48 | |
| Total amino acids (TAA) | 63.76 | 70.67 | 57.98 | 6.65 | |
| Essential amino acids (EAA) | 26.09 | 31.97 | 25.26 | 2.79 | |
| Non-essential amino acids (NEAA) | 47.15 | 38.7 | 32.72 | 3.86 | |
| Protein | 85.5 ± 4.9% | 71.9 ± 8.6% | |||
| Ash | 2.1 ± 0.4% | 1.7 ± 1.1% | |||
| Lipid | 3.27 ± 4.9% | 2.87 ± 1.27% |
| Spirulina sp. | Isochrysis sp. | ||||
|---|---|---|---|---|---|
| Spirulina platensis Whole Biomass (g/kg) | Spirulina platensis Protein Extract (g/kg) | Isochrysis T-Iso Whole Biomass (g/kg) | Isochrysis T-Iso Protein Extract (g/kg) | ||
| Essential amino acids | Isoleucine | 10.42 | 1.84 | 12.05 | 0.45 |
| Leucine | 25.53 | 2.31 | 17.63 | 0.55 | |
| Valine | 15.2 | 1.96 | 15.86 | 1.05 | |
| Phenylalanine | 32 | 2.63 | 12.3 | 0.54 | |
| Tyrptophan | 6.73 | . | 15.79 | . | |
| Histidine | 20.44 | 1.53 | 7.12 | 0.69 | |
| Lysine | 8.98 | 0.13 | 4.54 | . | |
| Threonine | 7.16 | 0.94 | 16.91 | 0.64 | |
| Methionine | 4.92 | . | 2.51 | . | |
| Non-essential amino acids | Alanine | 24.37 | 1.33 | 50.88 | 1.84 |
| Glycine | 3.92 | 0.55 | 12.45 | 0.14 | |
| Proline | 1.14 | 0.22 | 18.32 | . | |
| Tyrosine | 29.55 | 2.81 | 12.01 | 0.45 | |
| Aspartic acid and Asparagine | 6.08 | 1.08 | 7.43 | 0.69 | |
| Glutamic acid and Glutamine | 54.79 | 2 | 11.01 | 0.89 | |
| Arginine | 15.12 | 1.79 | 20.98 | . | |
| Serine | 8.65 | 0.56 | 10.14 | 0.59 | |
| Cysteine | 17.18 | 2.51 | 6.19 | 1.16 | |
| Total free amino acids (FAA) | 292.18 | 24.19 | 254.12 | 9.68 | |
| EAA | 131.38 | 11.34 | 104.71 | 3.92 | |
| NEAA | 160.8 | 12.85 | 149.41 | 5.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleakley, S.; Hayes, M. Functional and Bioactive Properties of Protein Extracts Generated from Spirulina platensis and Isochrysis galbana T-Iso. Appl. Sci. 2021, 11, 3964. https://doi.org/10.3390/app11093964
Bleakley S, Hayes M. Functional and Bioactive Properties of Protein Extracts Generated from Spirulina platensis and Isochrysis galbana T-Iso. Applied Sciences. 2021; 11(9):3964. https://doi.org/10.3390/app11093964
Chicago/Turabian StyleBleakley, Stephen, and Maria Hayes. 2021. "Functional and Bioactive Properties of Protein Extracts Generated from Spirulina platensis and Isochrysis galbana T-Iso" Applied Sciences 11, no. 9: 3964. https://doi.org/10.3390/app11093964
APA StyleBleakley, S., & Hayes, M. (2021). Functional and Bioactive Properties of Protein Extracts Generated from Spirulina platensis and Isochrysis galbana T-Iso. Applied Sciences, 11(9), 3964. https://doi.org/10.3390/app11093964