

Structure-Function Coupling Reveals Seizure Onset Connectivity Patterns

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Data
2.2. Image Acquisition
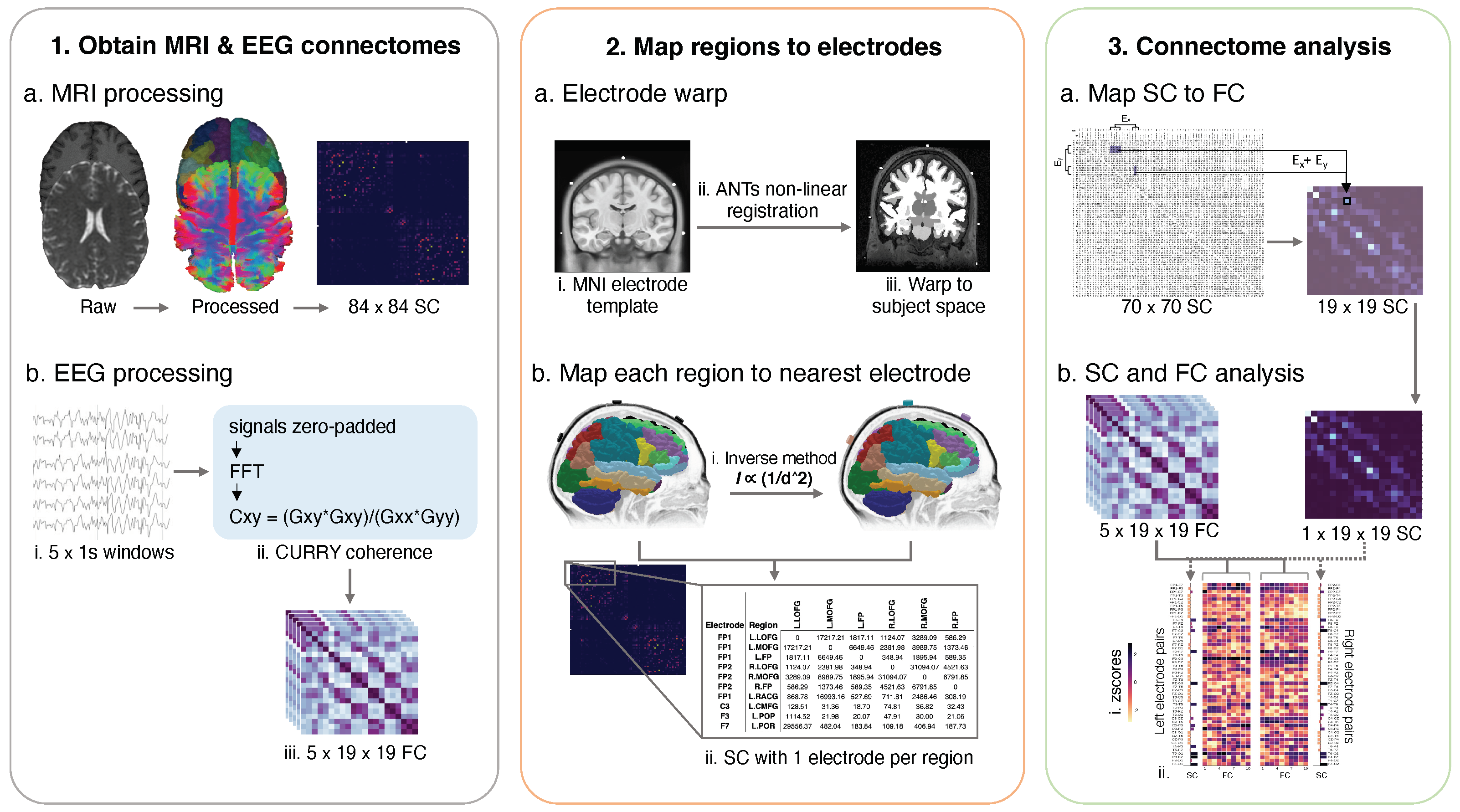
2.3. Image Processing to Obtain Structural Connectomes
2.4. EEG Acquisition
2.5. EEG Processing to Obtain Functional Connectomes
2.6. Mapping Cortical Regions to the Nearest Electrode
2.7. Mapping the Structural Connectome to the Functional Connectome
2.8. Statistical Analysis of Structure-Function Coupling
3. Results
3.1. Demographics
3.2. Electrode-Region Mapping
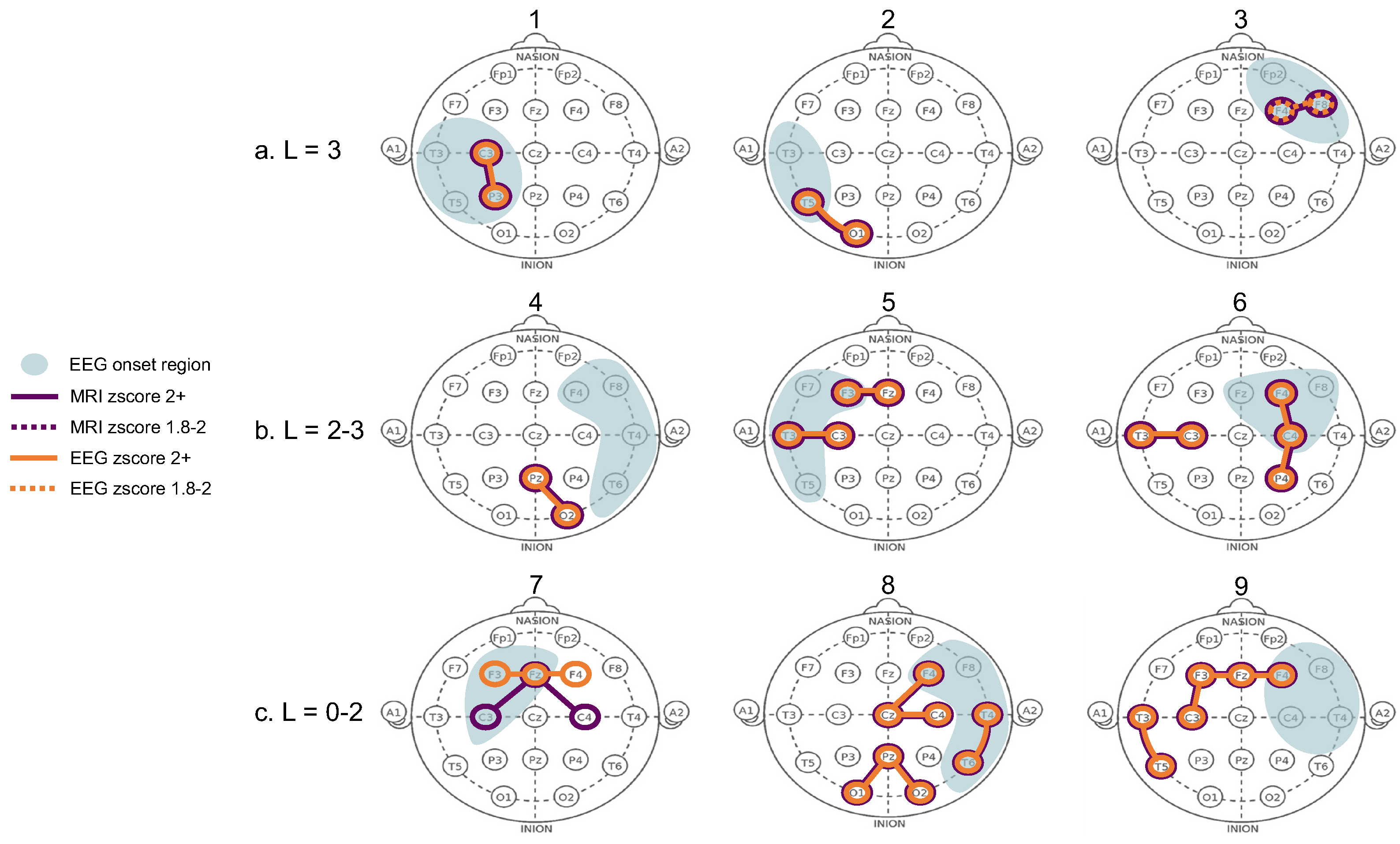
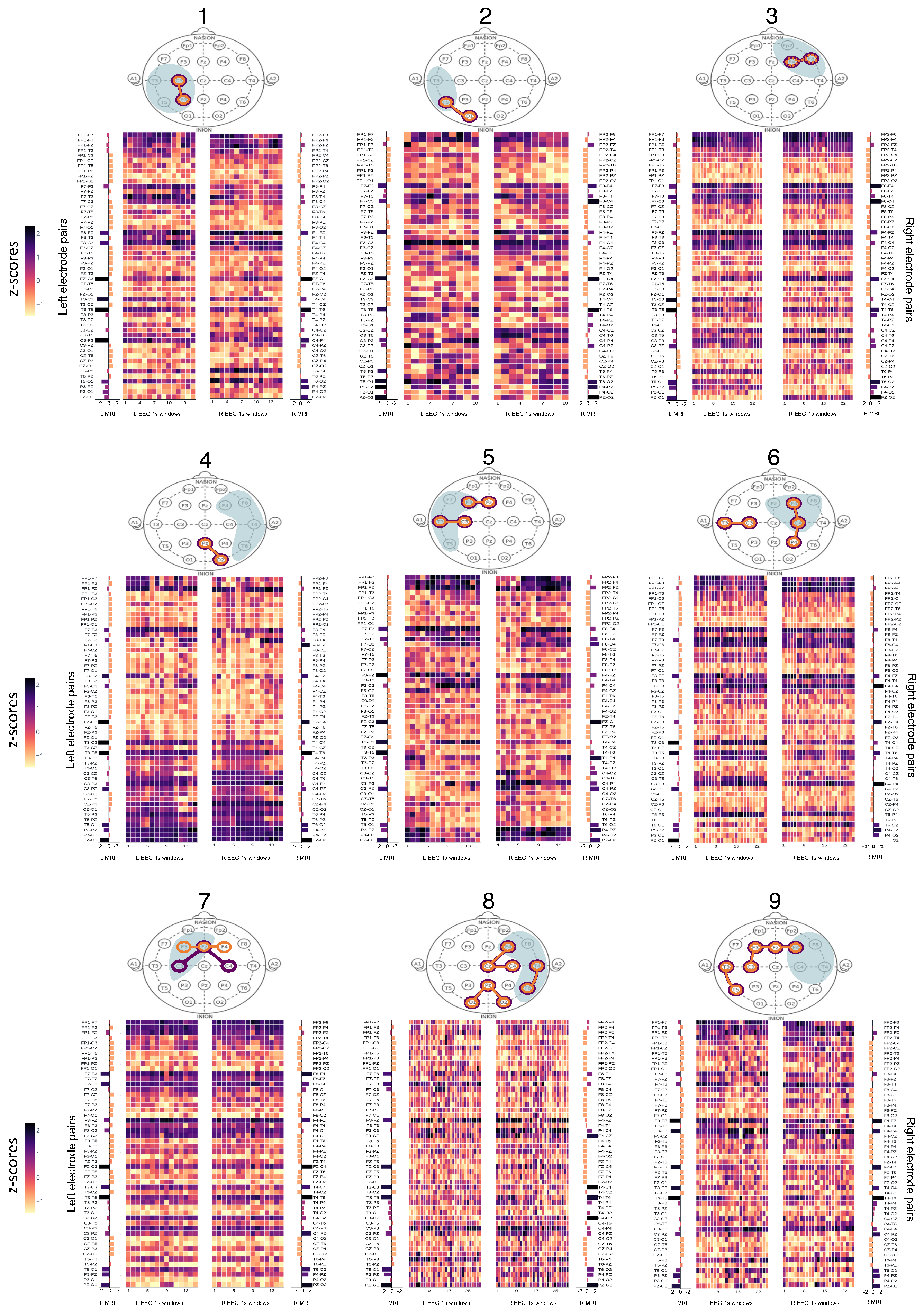
3.3. Structure-Function Coupling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Weng, Y.; Larivière, S.; Caciagli, L.; Vos de Wael, R.; Rodríguez-Cruces, R.; Royer, J.; Xu, Q.; Bernasconi, N.; Bernasconi, A.; Thomas Yeo, B.; et al. Macroscale and microcircuit dissociation of focal and generalized human epilepsies. Commun. Biol. 2020, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zijlmans, M.; Zweiphenning, W.; van Klink, N. Changing concepts in presurgical assessment for epilepsy surgery. Nat. Rev. Neurol. 2019, 15, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Babaeeghazvini, P.; Rueda-Delgado, L.M.; Gooijers, J.; Swinnen, S.P.; Daffertshofer, A. Brain Structural and Functional Connectivity: A Review of Combined Works of Diffusion Magnetic Resonance Imaging and Electro-Encephalography. Front. Hum. Neurosci. 2021, 15, 585. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting human resting-state functional connectivity from structural connectivity. Proc. Natl. Acad. Sci. USA 2009, 106, 2035–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Ashourvan, A.; Mikhail, F.; Pines, A.; Kini, L.; Oechsel, K.; Das, S.R.; Stein, J.M.; Shinohara, R.T.; Bassett, D.S.; et al. Characterizing the role of the structural connectome in seizure dynamics. Brain 2019, 142, 1955–1972. [Google Scholar] [CrossRef]
- Vattikonda, A.N.; Hashemi, M.; Sip, V.; Woodman, M.M.; Bartolomei, F.; Jirsa, V.K. Identifying spatio-temporal seizure propagation patterns in epilepsy using Bayesian inference. Commun. Biol. 2021, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lascano, A.M.; Perneger, T.; Vulliemoz, S.; Spinelli, L.; Garibotto, V.; Korff, C.M.; Vargas, M.I.; Michel, C.M.; Seeck, M. Yield of MRI, high-density electric source imaging (HD-ESI), SPECT and PET in epilepsy surgery candidates. Clin. Neurophysiol. 2016, 127, 150–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, N.; Duncan, J.S.; Diehl, B.; Chowdhury, F.A.; de Tisi, J.; Miserocchi, A.; McEvoy, A.W.; Davis, K.A.; Vos, S.B.; Winston, G.P.; et al. Intracranial EEG structure-function coupling predicts surgical outcomes in focal epilepsy. arXiv 2022, arXiv:2204.08086. [Google Scholar] [CrossRef]
- Chiang, S.; Stern, J.M.; Engel, J., Jr.; Haneef, Z. Structural–functional coupling changes in temporal lobe epilepsy. Brain Res. 2015, 1616, 45–57. [Google Scholar] [CrossRef]
- O’Muircheartaigh, J.; Keller, S.S.; Barker, G.J.; Richardson, M.P. White matter connectivity of the thalamus delineates the functional architecture of competing thalamocortical systems. Cereb. Cortex 2015, 25, 4477–4489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, V.L.; Sainburg, L.E.; Johnson, G.W.; Janson, A.; Levine, K.K.; Rogers, B.P.; Chang, C.; Englot, D.J. Presurgical temporal lobe epilepsy connectome fingerprint for seizure outcome prediction. Brain Commun. 2022, 4, fcac128. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Luo, C.; Chang, X.; Zhang, R.; Klugah-Brown, B.; Guo, L.; Xu, P.; Yao, D. White matter connectivity pattern associate with characteristics of scalp EEG signals. Brain Topogr. 2017, 30, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Deslauriers-Gauthier, S.; Lina, J.M.; Butler, R.; Whittingstall, K.; Gilbert, G.; Bernier, P.M.; Deriche, R.; Descoteaux, M. White matter information flow mapping from diffusion MRI and EEG. NeuroImage 2019, 201, 116017. [Google Scholar] [CrossRef] [Green Version]
- Sadaghiani, S.; Brookes, M.J.; Baillet, S. Connectomics of human electrophysiology. NeuroImage 2022, 247, 118788. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, S.; Buzori, S.; Trebuchon, A.; Carron, R.; Scavarda, D.; Milh, M.; McGonigal, A.; Bartolomei, F. The repertoire of seizure onset patterns in human focal epilepsies: Determinants and prognostic values. Epilepsia 2019, 60, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, Z.; Morita-Sherman, M.; Hogue, O.; Joseph, B.; Alvim, M.K.; Yasuda, C.L.; Vegh, D.; Nair, D.; Burgess, R.; Bingaman, W.; et al. Improving the prediction of epilepsy surgery outcomes using basic scalp EEG findings. Epilepsia 2021, 62, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.J.; Tanaka, N.; Diaz, J.; Edlow, B.L.; Wu, O.; Hämäläinen, M.; Stufflebeam, S.; Cash, S.S.; Kramer, M.A. EEG functional connectivity is partially predicted by underlying white matter connectivity. Neuroimage 2015, 108, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cserpan, D.; Gennari, A.; Gaito, L.; Lo Biundo, S.P.; Tuura, R.; Sarnthein, J.; Ramantani, G. Scalp HFO rates are higher for larger lesions. Epilepsia Open 2022, 7, 496–503. [Google Scholar] [CrossRef]
- Samadzadehaghdam, N.; Makkiabadi, B.; Masjoodi, S.; Mohammadi, M.; Mohagheghian, F. A new linearly constrained minimum variance beamformer for reconstructing EEG sparse sources. Int. J. Imaging Syst. Technol. 2019, 29, 686–700. [Google Scholar] [CrossRef]
- Rubega, M.; Carboni, M.; Seeber, M.; Pascucci, D.; Tourbier, S.; Toscano, G.; Van Mierlo, P.; Hagmann, P.; Plomp, G.; Vulliemoz, S.; et al. Estimating EEG source dipole orientation based on singular-value decomposition for connectivity analysis. Brain Topogr. 2019, 32, 704–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neugebauer, F.; Antonakakis, M.; Unnwongse, K.; Parpaley, Y.; Wellmer, J.; Rampp, S.; Wolters, C.H. Validating EEG, MEG and combined MEG and EEG beamforming for an estimation of the epileptogenic zone in focal cortical dysplasia. Brain Sci. 2022, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Zagorchev, L.; Brueck, M.; Fläschner, N.; Wenzel, F.; Hyde, D.; Ewald, A.; Peters, J. Patient-specific sensor registration for electrical source imaging using a deformable head model. IEEE Trans. Biomed. Eng. 2020, 68, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Proix, T.; Bartolomei, F.; Guye, M.; Jirsa, V.K. Individual brain structure and modelling predict seizure propagation. Brain 2017, 140, 641–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jirsa, V.K.; Proix, T.; Perdikis, D.; Woodman, M.M.; Wang, H.; Gonzalez-Martinez, J.; Bernard, C.; Bénar, C.; Guye, M.; Chauvel, P.; et al. The virtual epileptic patient: Individualized whole-brain models of epilepsy spread. Neuroimage 2017, 145, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Varatharajah, Y.; Berry, B.; Joseph, B.; Balzekas, I.; Pal Attia, T.; Kremen, V.; Brinkmann, B.; Iyer, R.; Worrell, G. Characterizing the electrophysiological abnormalities in visually reviewed normal EEGs of drug-resistant focal epilepsy patients. Brain Commun. 2021, 3, fcab102. [Google Scholar] [CrossRef] [PubMed]
- Asadzadeh, S.; Rezaii, T.Y.; Beheshti, S.; Delpak, A.; Meshgini, S. A systematic review of EEG source localization techniques and their applications on diagnosis of brain abnormalities. J. Neurosci. Methods 2020, 339, 108740. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Seeck, M.; Beniczky, S. Accuracy of interictal and ictal electric and magnetic source imaging: A systematic review and meta-analysis. Front. Neurol. 2019, 10, 1250. [Google Scholar] [CrossRef] [Green Version]
- Birot, G.; Spinelli, L.; Vulliémoz, S.; Mégevand, P.; Brunet, D.; Seeck, M.; Michel, C.M. Head model and electrical source imaging: A study of 38 epileptic patients. NeuroImage Clin. 2014, 5, 77–83. [Google Scholar] [CrossRef]
- Maher, C.; D’Souza, A.; Zeng, R.; Wang, D.; Barnett, M.; Kavehei, O.; Armin, N.; Wang, C. Automated method to map cortical brain regions to the nearest scalp electroencephalography electrode. Epilepsia 2021, 62, 238–239. [Google Scholar]
- Rados, M.; Mouthaan, B.; Barsi, P.; Carmichael, D.; Heckemann, R.A.; Kelemen, A.; Kobulashvili, T.; Kuchukhidze, G.; Marusic, P.; Minkin, K.; et al. Diagnostic value of MRI in the presurgical evaluation of patients with epilepsy: Influence of field strength and sequence selection: A systematic review and meta-analysis from the E-PILEPSY Consortium. Epileptic Disord. 2022, 24, 323–342. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker,, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.; D’Souza, A.; Zeng, R.; Barnett, M.; Kavehei, O.F.; Nikpour, A.; Wang, C. White matter alterations in focal to bilateral tonic-clonic seizures. Front. Neurol. 2022, 13, 972590. [Google Scholar] [CrossRef]
- Andersson, J.L.; Sotiropoulos, S.N. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage 2016, 125, 1063–1078. [Google Scholar] [CrossRef] [Green Version]
- Fischl, B. Freesurfer. Neuroimage 2012, 62, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.; Skoch, A.; Bajada, C.J.; Caspers, S.; Connelly, A. Hybrid surface-volume segmentation for improved anatomically-constrained tractography. In Proceedings of the OHBM Annual Meeting, Virtual, 23 June–3 July 2020; pp. 1–5. [Google Scholar]
- Tournier, J.D.; Smith, R.; Raffelt, D.; Tabbara, R.; Dhollander, T.; Pietsch, M.; Christiaens, D.; Jeurissen, B.; Yeh, C.H.; Connelly, A. MRtrix3: A fast, flexible and open software framework for medical image processing and visualisation. NeuroImage 2019, 202, 116137. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 2004, 23, S208–S219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 bias correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef] [Green Version]
- Dhollander, T.; Raffelt, D.; Connelly, A. Unsupervised 3-tissue response function estimation from single-shell or multi-shell diffusion MR data without a co-registered T1 image. In Proceedings of the ISMRM Workshop on Breaking the Barriers of Diffusion MRI. ISMRM, Lisbon, Portugal, 12–16 September 2016; Volume 5. [Google Scholar]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. Anatomically-constrained tractography: Improved diffusion MRI streamlines tractography through effective use of anatomical information. NeuroImage 2012, 62, 1924–1938. [Google Scholar] [CrossRef]
- Tournier, J.D.; Calamante, F.; Connelly, A. Improved probabilistic streamlines tractography by 2nd order integration over fibre orientation distributions. In Proceedings of the International Society for Magnetic Resonance in Medicine, Stockholm, Sweden, 1–7 May 2010; p. 1670. [Google Scholar]
- Smith, R.E.; Tournier, J.D.; Calamante, F.; Connelly, A. SIFT2: Enabling dense quantitative assessment of brain white matter connectivity using streamlines tractography. NeuroImage 2015, 119, 338–351. [Google Scholar] [CrossRef]
- Justesen, A.B.; Foged, M.T.; Fabricius, M.; Skaarup, C.; Hamrouni, N.; Martens, T.; Paulson, O.B.; Pinborg, L.H.; Beniczky, S. Diagnostic yield of high-density versus low-density eeg: The effect of spatial sampling, timing and duration of recording. Clin. Neurophysiol. 2019, 130, 2060–2064. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.A.; Devries, S.P.; Krieger, A.; Mihaylova, T.; Minecan, D.; Litt, B.; Wagenaar, J.B.; Stacey, W.C. The effect of increased intracranial EEG sampling rates in clinical practice. Clin. Neurophysiol. 2018, 129, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, G.M.; Chowdhury, F.A.; Cook, M.J.; Diehl, B.; Duncan, J.S.; Karoly, P.J.; Taylor, P.N.; Wang, Y. Multiple mechanisms shape the relationship between pathway and duration of focal seizures. Brain Commun. 2022, 4, fcac173. [Google Scholar] [CrossRef] [PubMed]
- Suárez, L.E.; Markello, R.D.; Betzel, R.F.; Misic, B. Linking structure and function in macroscale brain networks. Trends Cogn. Sci. 2020, 24, 302–315. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Classification | MRI Diagnosis | Onset Age | Age at MRI | Duration | Drug Res. | Handedness |
|---|---|---|---|---|---|---|---|---|
| 1 | F | Left fronto-temporal | Normal | 49 | 53 | 4 | Y | R |
| 2 | M | Left fronto-temporal | HS | 21 | 49 | 28 | Y | L |
| 3 | F | Right frontal | Normal | 38 | 48 | 10 | N | R |
| 4 | F | Right temporal | Normal | 16 | 29 | 13 | Y | U |
| 5 | M | Left fronto-temporal | Normal | 16 | 31 | 15 | Y | R |
| 6 | F | Left occipital | Normal | 12 | 47 | 35 | Y | R |
| 7 | F | Left fronto-temporal | Normal | 35 | 48 | 13 | N | R |
| 8 | M | Right fronto-temporal | Normal | 15 | 33 | 18 | Y | R |
| 9 | M | Right temporal | Normal | 22 | 29 | 7 | Y | R |
| Subject No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Region Name | |||||||||
| L. rostralanteriorcingulate | FP1 | FP1 | FP1 | FZ | F3 | FP1 | FZ | FP1 | FP1 |
| R. rostralanteriorcingulate | FP2 | FP2 | FP2 | FZ | FP2 | FP2 | FZ | F4 | FP2 |
| L. parsopercularis | F3 | F7 | F7 | F3 | F7 | F7 | T3 | F7 | F3 |
| R. parsopercularis | F8 | F8 | F8 | F8 | F8 | F8 | F4 | T4 | F4 |
| L. insula | T3 | F7 | T3 | F7 | T3 | T3 | T3 | T3 | T3 |
| R. insula | T4 | F8 | F8 | F8 | F8 | T4 | C4 | T4 | T4 |
| L. inferiortemporal | T5 | T5 | T5 | T5 | T3 | T5 | T5 | T3 | T5 |
| R. inferiortemporal | T6 | T4 | T6 | T6 | T4 | T6 | T6 | T4 | T6 |
| L. lateralorbitofrontal | FP1 | F7 | F7 | F7 | F7 | F7 | F7 | F7 | F7 |
| R. lateralorbitofrontal | FP2 | F8 | F8 | F8 | F8 | F8 | F4 | F8 | FP2 |
| L. cuneus | O1 | O1 | O1 | O1 | O1 | O1 | PZ | O1 | O1 |
| R. cuneus | O2 | O2 | O2 | PZ | O2 | O2 | PZ | O2 | O2 |
| L. transversetemporal | T3 | T3 | T3 | C3 | T3 | T3 | T3 | T3 | T3 |
| R. transversetemporal | T4 | T4 | T4 | C4 | T4 | T4 | T4 | T4 | T4 |
| L. caudalanteriorcingulate | FZ | FZ | FZ | FZ | F3 | FZ | FZ | F3 | FZ |
| R. caudalanteriorcingulate | FZ | FZ | FZ | FZ | FZ | FZ | FZ | F4 | FZ |
| L. isthmuscingulate | PZ | PZ | PZ | CZ | PZ | PZ | PZ | PZ | PZ |
| R. isthmuscingulate | PZ | PZ | PZ | CZ | PZ | PZ | PZ | PZ | PZ |
| R. bankssts | T6 | T6 | T4 | T6 | T4 | T6 | T6 | T6 | T6 |
| R. superiorfrontal | FZ | FZ | FZ | FZ | FZ | FZ | FZ | CZ | FZ |
| R. caudalmiddlefrontal | C4 | C4 | C4 | F4 | C4 | C4 | C4 | C4 | C4 |
| R. temporalpole | F8 | F8 | F8 | F8 | F8 | F8 | F8 | T4 | F8 |
| L. supramarginal | C3 | P3 | C3 | C3 | C3 | C3 | C3 | C3 | C3 |
| L. superiorparietal | P3 | PZ | PZ | PZ | PZ | PZ | PZ | PZ | PZ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maher, C.; D’Souza, A.; Barnett, M.; Kavehei, O.; Wang, C.; Nikpour, A. Structure-Function Coupling Reveals Seizure Onset Connectivity Patterns. Appl. Sci. 2022, 12, 10487. https://doi.org/10.3390/app122010487
Maher C, D’Souza A, Barnett M, Kavehei O, Wang C, Nikpour A. Structure-Function Coupling Reveals Seizure Onset Connectivity Patterns. Applied Sciences. 2022; 12(20):10487. https://doi.org/10.3390/app122010487
Chicago/Turabian StyleMaher, Christina, Arkiev D’Souza, Michael Barnett, Omid Kavehei, Chenyu Wang, and Armin Nikpour. 2022. "Structure-Function Coupling Reveals Seizure Onset Connectivity Patterns" Applied Sciences 12, no. 20: 10487. https://doi.org/10.3390/app122010487
APA StyleMaher, C., D’Souza, A., Barnett, M., Kavehei, O., Wang, C., & Nikpour, A. (2022). Structure-Function Coupling Reveals Seizure Onset Connectivity Patterns. Applied Sciences, 12(20), 10487. https://doi.org/10.3390/app122010487