
Composition of the Wild Rabbit Nest and Its Implication for Domestic Rabbit Breeding


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Experiment 1
2.3. Experiment 2
2.4. Experiment 3
2.5. Statistical Analyses
3. Results
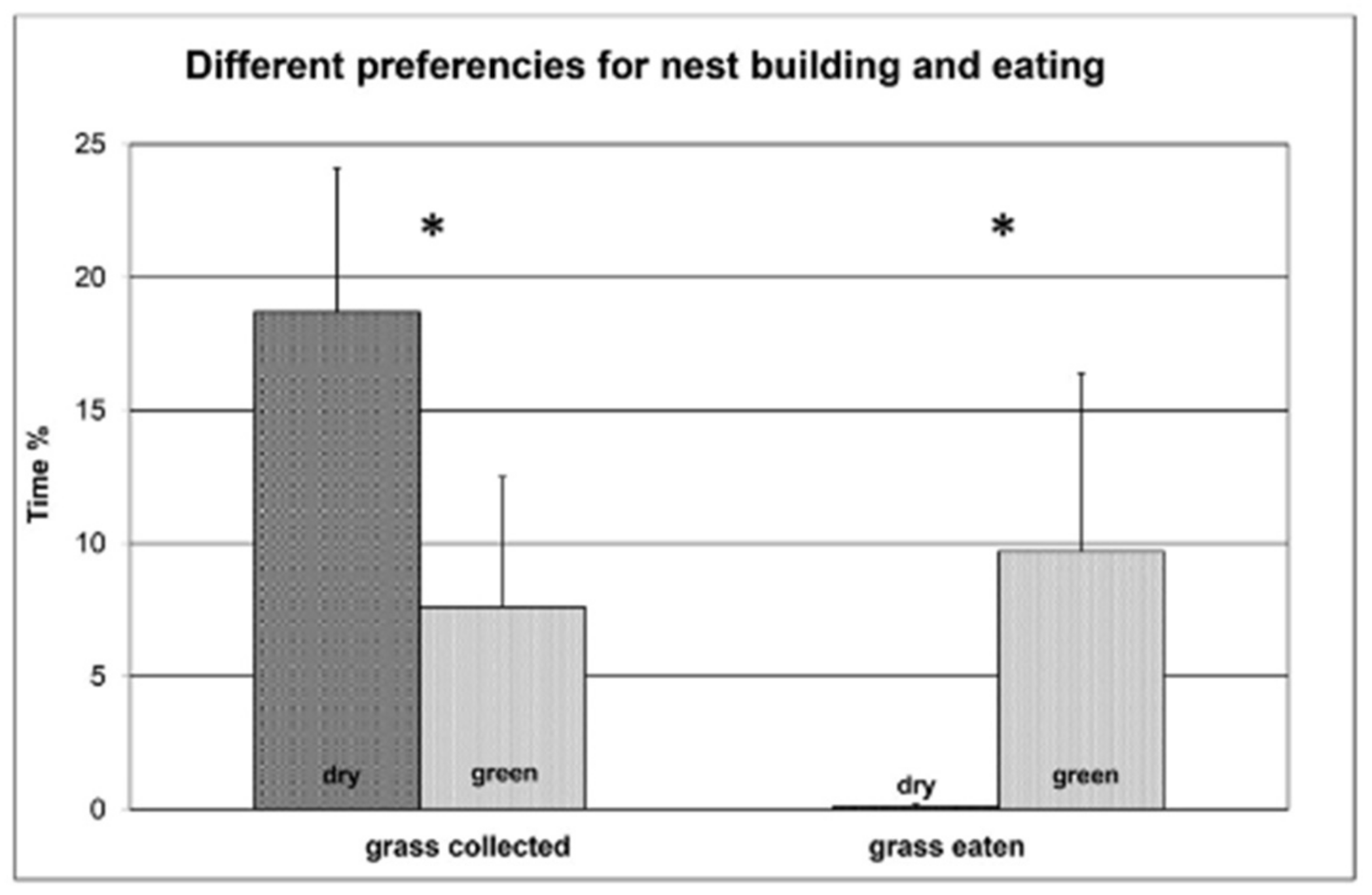
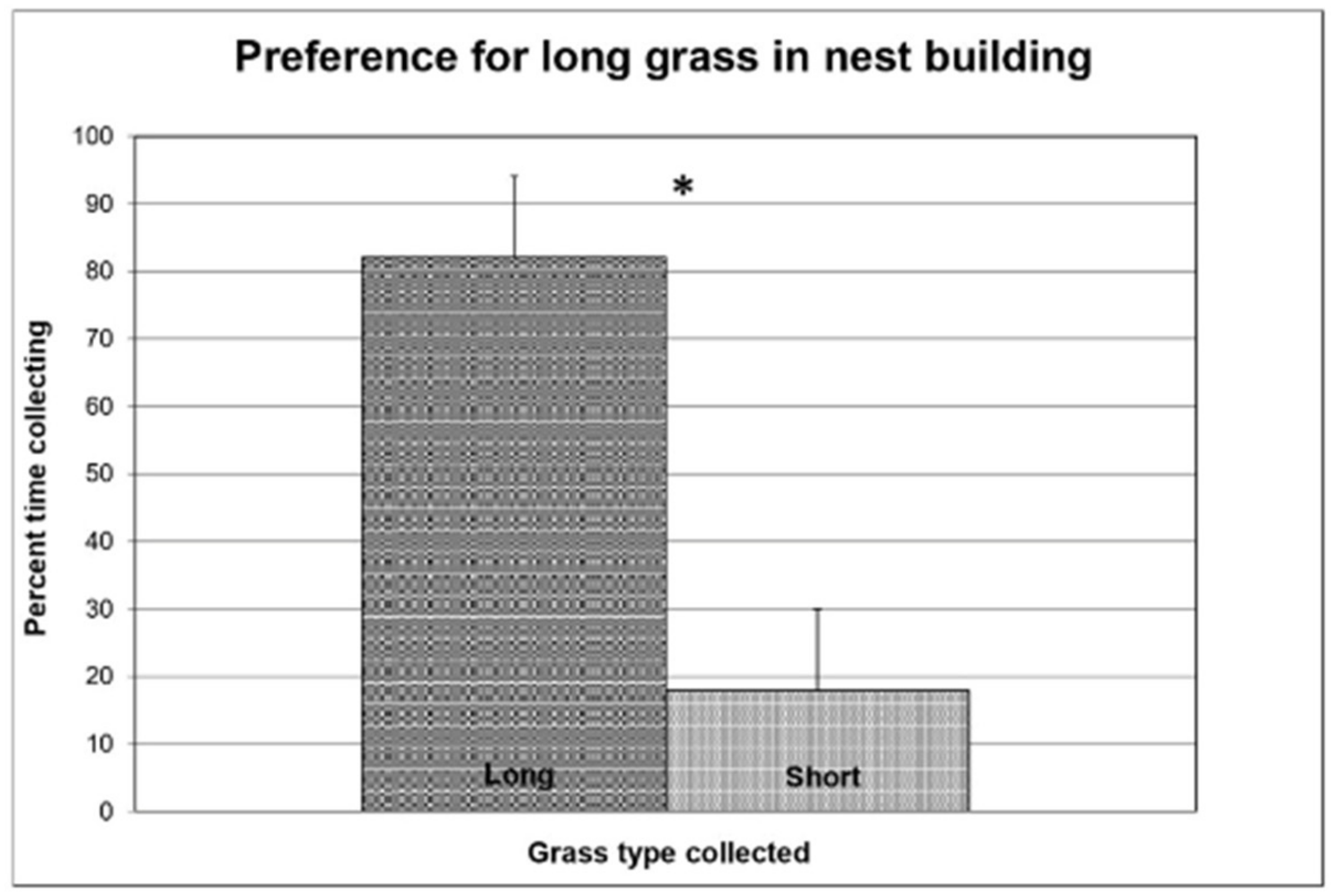
3.1. Experiment 1
3.2. Experiment 2
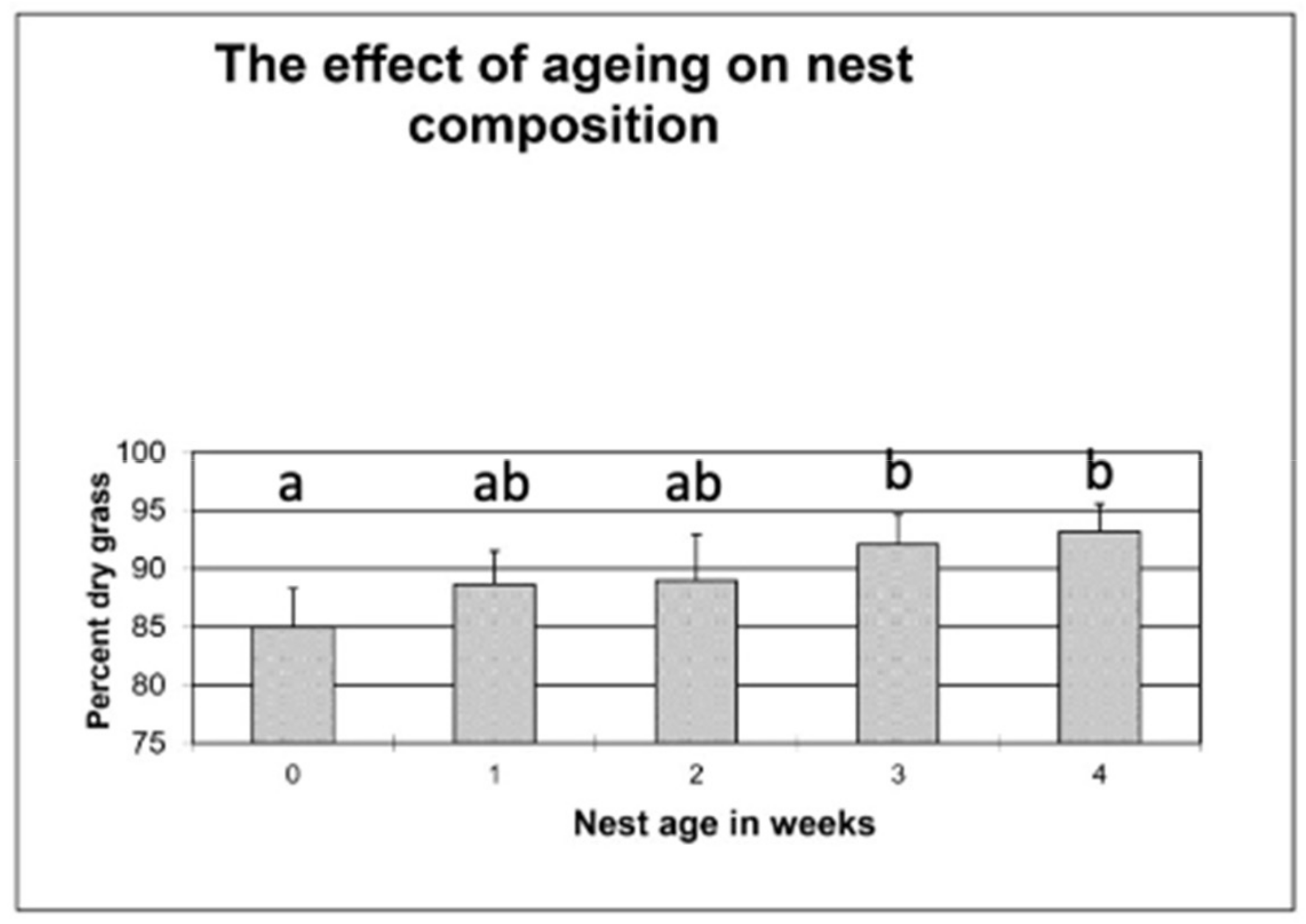
3.3. Experiment 3
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hansell, M.H. Animal Architecture & Building Behaviour; Longman: London, UK, 1984. [Google Scholar]
- Casey, T.M. Nest insulation: Energy savings to brown lemmings using a winter nest. Oecologia 1981, 50, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaskill, B.N.; Gordon, C.J.; Pajor, E.A.; Lucas, J.R.; Davis, J.K.; Garner, J.P. Impact of nesting material on mouse body temperature and physiology. Physiol. Behav. 2013, 110, 87–95. [Google Scholar] [CrossRef] [PubMed]
- De-Oliveira, M.C.; Lima, S.C.O.; Mesquita, S.A.; Silva, J.A.; Gomes, Y.S.; Attia, Y.A.; Oliveira, H.C. Nesting materials for does: Effect on nest building and performance at first parturition. Rev. Colomb. Cienc. Pecu. 2017, 30, 308–315. [Google Scholar] [CrossRef]
- Bethge, P.; Munks, S.; Otley, H.; Nicol, S. Platypus burrow temperatures at a subalpine Tasmanian lake. Proc. Linn. Soc. N. S. W. 2004, 125, 273–276. [Google Scholar]
- Geiser, F. Reduction of metabolism during hibernation and daily torpor in mammals and birds—Temperature effect or physiological inhibition? J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1988, 158, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Houston, A.I.; McNamara, J.M. A theoretical investigation of the fat reserves and mortality levels of small birds in winter. Ornis Scand. 1993, 24, 205–219. [Google Scholar] [CrossRef]
- Lovegrove, B.G.; Raman, J.; Perrin, M.R. Heterothermy in elephant shrews, Elephantulus spp. (Macroscelidea): Daily torpor or hibernation? J. Comp. Physiol. B Biochem. Syst. Envrion. Physiol. 2001, 171, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, D.J.; Moncrieff, J.B.; Taylor, I.R. Winter microclimate of field voles (Microtus agrestis) in SW Scotland. J. Therm. Biol. 2003, 28, 397–401. [Google Scholar] [CrossRef]
- Redman, P.; Selman, C.; Speakman, J.R. Male short-tailed field voles (Microtus agrestis) build better insulated nests than females. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1999, 169, 581–587. [Google Scholar] [CrossRef]
- Barclay, R.; Lausen, C.L.; Hollis, L. What’s hot and what’s not: Defining torpor in free-ranging birds and mammals. Can. J. Zool. 2001, 79, 1885–1890. [Google Scholar] [CrossRef]
- Pinowski, J.; Haman, A.; Jerzak, L.; Pinowska, B.; Barkowska, M.; Grodzki, A.; Haman, K. The thermal properties of some nests of the Eurasian tree sparrow, Passer montanus. J. Therm. Biol. 2006, 31, 573–581. [Google Scholar] [CrossRef]
- Lamprecht, I.; Schmolz, E. Thermal investigations of some bird nests. Thermochim. Acta 2006, 415, 141–148. [Google Scholar] [CrossRef]
- Zarrow, M.X.; Farooq, A.; Denenberg, V.H.; Sawin, P.B.; Ross, S. Maternal behaviour in the rabbit: Endocrine control of maternal nest building. J. Reprod. Fertil. 1963, 6, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, A.; Drummond, H.; Martínez-Gómez, M.; Hudson, R. Thermal benefit of sibling presence in the newborn rabbit. Dev. Psychobiol. 2003, 43, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Swihart, R.K. Body size, breeding season length, and life history tactics of Lagomorphs. Oikos 1984, 43, 282–290. [Google Scholar] [CrossRef]
- Broekhuizen, S.; Mulder, J. Differences and similarities in nursing behaviour of hares and rabbits. Acta Zool. Fenn. 1983, 174, 61–63. [Google Scholar]
- Mykytowycz, R. Social behaviour of an experimental colony of wild rabbits, Oryctolagus cuniculus (L.). CS.IRO Wildl. Res. 1958, 3, 7–25. [Google Scholar] [CrossRef]
- Henderson, B.A. Regulation of the size of the breeding population of the European rabbit, Oryctolagus cuniculus, by social behaviour. J. Appl. Ecol. 1979, 16, 383–392. [Google Scholar] [CrossRef]
- Lloyd, M.; McCowan, D. Some observations on the breeding burrows of the wild rabbit Oryctolagus cuniculus on the island of Skokholm. J. Zool. 1968, 156, 540–549. [Google Scholar] [CrossRef]
- Delibes, M.; Hiraldo, F. The rabbit as prey in the Iberian Mediterranean ecosystem. In Proceedings of the World Lagomorph Conference, Guelph, ON, Canada, 12-16 August 1979; Myers, K., MacInnes, C.D., Eds.; University of Guelph: Guelph, ON, Canada, 1979; pp. 614–622. [Google Scholar]
- Jaksic, F.M.; Soriguer, R. Predation upon the European rabbit Oryctolagus cuniculus in Mediterranean habitats of Chile and Spain: A comparative analysis. J. Anim. Ecol. 1981, 50, 269–281. [Google Scholar] [CrossRef]
- Rödel, H.G.; Starkloff, A.; Seltmann, M.W.; Prager, G.; von Holst, D. Causes and predictors of nest mortality in a European rabbit population. Mammal. Biol. 2009, 74, 198–209. [Google Scholar] [CrossRef]
- Künkele, J. Infanticide in wild rabbits (Oryctolagus cuniculus). J. Mammal. 1991, 73, 317–320. [Google Scholar] [CrossRef]
- Hudson, R.; Distel, H. The pattern of behaviour of rabbit pups in the nest. Behaviour 1982, 79, 255–271. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.H. The demography of a rabbit population in an arid region of New South Wales Australia. J. Anim. Ecol. 1980, 49, 55–79. [Google Scholar] [CrossRef]
- Parer, I.; Libke, J.A. Distribution of rabbit, Oryctolagus cuniculus, warrens in relation to soil type. Aust. Wildl. Res. 1985, 12, 387–405. [Google Scholar] [CrossRef]
- Jilge, B. The ontogeny of circadian rhythms in the rabbit. J. Biol. Rhythm. 1993, 8, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, R.; Bilkó, A.; Altbäcker, V. Nursing, weaning and the development of independent feeding in the rabbit. Z. Saeugetierkunde 1996, 61, 39–48. [Google Scholar]
- Cowie, A.T. Variations in the yield and composition of the milk during lactation in the rabbit and the galactopoietic effect of the prolactin. J. Endocrinol. 1969, 44, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, H.H.; Lukefahr, S.D.; McNitt, J.I. Maternal nest quality and its influence on litter survival and weaning performance in commercial rabbits. J. Anim. Sci. 1997, 75, 926–933. [Google Scholar] [CrossRef]
- Denenberg, V.H.; Huff, R.L.; Ross, S.; Sawin, P.B.; Zarrow, M.X. Maternal behaviour in the rabbit: The quantification of nest building. Anim. Behav. 1963, 11, 494–499. [Google Scholar] [CrossRef]
- Ross, S.; Zarrow, M.X.; Sawin, P.B.; Denenberg, V.H.; Blumenfield, M. Maternal behaviour in the rabbit under seminatural conditions. Anim. Behav. 1963, 11, 283–285. [Google Scholar] [CrossRef]
- Verga, M.; Dell’Orto, V.; Carenzi, C. A general review and survey of maternal behaviour in the rabbit. Appl. Anim. Ethol. 1978, 4, 235–252. [Google Scholar] [CrossRef]
- Farkas, P.; Szendrő, Z.; Matics, Z.; Radnai, I.; Nagy, I.; Gerencsér, Z. Preference of rabbit does among different nest materials. World Rabbit Sci. 2018, 26, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Farkas, T.P.; Szendrő, Z.; Matics, Z.; Mayer, A.; Radnai, I.; Odermatt, M.; Gerencsér, Z. Effect of different nest materials on nest quality and reproduction performance of rabbit does of various breeds. Állattenyésztés Takarmányozás 2016, 65, 35–41. [Google Scholar]
- González-Redondo, P. Maternal behaviour in peripartum influences preweaning kit mortality in cage-bred wild rabbits. World Rabbit Sci. 2010, 18, 91–102. [Google Scholar] [CrossRef] [Green Version]
- González-Mariscal, G.; Caba, M.; Martínez-Gómez, M.; Bautista, A.; Hudson, R. Mothers and offspring: The rabbit as a model system in the study of mammalian maternal behavior and sibling interactions. Horm. Behav. 2016, 77, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Zarrow, M.X.; Sawin, P.B.; Ross, S.; Denenberg, V.; Crary, D.; Wilson, E.D.; Farooq, A. Maternal behaviour in the rabbit: Evidence for an endocrine basis of nest building and additional data on maternal nest building in the Dutch-belted race. J. Reprod. Fertil. 1961, 2, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.S. The Behaviour of Domestic Animals; Bailliere & Tindall: London, UK, 1969. [Google Scholar]
- Coureaud, G.; Schaal, B.; Coudert, P.; Hudson, R.; Rideaud, P.; Orgeur, P. Mimicking natural nursing conditions promotes early pup survival in domestic rabbits. Ethology 2000, 106, 207–225. [Google Scholar] [CrossRef]
- González-Mariscal, G.; Rosenblatt, J.S. Maternal behavior in rabbits. A historical and multidimensional perspective. Adv. Study Behav. 1996, 25, 333–359. [Google Scholar]
- González-Mariscal, G. Neuroendocrinology applied to rabbit breeding. World Rabbit Sci. 2021, 29, 231–238. [Google Scholar] [CrossRef]
- Baumans, V. Environmental enrichment for laboratory rodents and rabbits: Requirements of rodents, rabbits and research. Ilar J. 2005, 46, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Flux, J.E.C.; Fullagar, P.J. World distribution of the rabbit (Oryctolagus cuniculus). Acta Zool. Fenn. 1983, 174, 75–77. [Google Scholar]
- Homolka, M. A comparison of the trophic niches of Lepus europeus and Oryctolagus cuniculus. Folia Zool. 1987, 36, 307–317. [Google Scholar]
- Mátrai, K.; Altbäcker, V.; Hahn, I. Seasonal diet of rabbits and their effect on juniper in Bugac Juniper Forest (Hungary). Acta Theriol. 1998, 43, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Altbäcker, V.; Hudson, R.; Bilkó, Á. Can rabbit mother’s diet influence the pups later food choice? Ethology 1995, 99, 107–116. [Google Scholar] [CrossRef]
- Brüll, U. Nahrungsbiologische Studien am Feldhasen in Schleswig-Holstein. Ein Beitrag zu Asungsverbesserung. In Ecology and Management of European Hare Populations; Pielowski, Z., Pucek, Z., Eds.; PWRL: Warsawa, Poland, 1976; pp. 93–103. [Google Scholar]
- Deutsch, J. Nest building behaviour of domestic rabbits under seminatural conditions. Br. J. Anim. Behav. 1957, 5, 53–54. [Google Scholar] [CrossRef]
- González-Mariscal, G.; Melo, A.I.; Jiménez, P.; Beyer, C.; Rosenblatt, J.S. Estradiol, progesterone, and prolactin regulate maternal nest-building in rabbits. J. Neuroendocr. 1996, 8, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Millar, J.S.; Zwickel, F.C. Characteristics and ecological significance of haypiles of pikas. Mammalia 1972, 36, 58–68. [Google Scholar] [CrossRef]
- Gedeon, C.I.; Markó, G.; Németh, I.; Nyitrai, V.; Altbäcker, V. Nest material selection affects nest insulation quality for the European ground squirrel (Spermophilus citellus). J. Mammal. 2010, 91, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Mcgowan, A.; Sharp, S.P.; Hatchwell, B.J. The structure and function of nests of long-tailed tits, Aegithalos caudatus. Funct. Ecol. 2004, 18, 578–583. [Google Scholar] [CrossRef]
- Lanteigne, M.; Reebs, S.G. Preference for bedding material in Syrian hamsters. Lab. Anim. 2006, 40, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Casteel, D.A. Nest building, parturition and copulation in the Cottontail rabbit. Am. Midl. Nat. 1966, 75, 160–167. [Google Scholar] [CrossRef]
- Rosvold, E.M.; Newberry, R.C.; Andersen, I.L. Early mother-young interactions in domestic sows—Nest-building material increases maternal investment. Appl. Anim. Behav. Sci. 2019, 219, 104837. [Google Scholar] [CrossRef]
- Ruiz-Castellano, C.; Tomas, G.; Ruiz-Rodriguez, M.; Martin-Galvez, D.; Soler, J.J. Nest material shapes eggs bacterial environment. PLoS ONE 2016, 11, e0148894. [Google Scholar] [CrossRef] [Green Version]
- Peles, F.; Sipos, P.; Győri, Z.; Pfliegler, W.P.; Giacometti, F.; Serraino, A.; Pócsi, I. Adverse effects, transformation and channeling of aflatoxins into food raw materials in livestock. Front. Microbial. 2019, 10, 2861. [Google Scholar] [CrossRef] [Green Version]
- Schlolaut, W.; Hudson, R.; Rödel, H.G. Impact of rearing management on health in domestic rabbits: A review. World Rabbit Sci. 2013, 21, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Trocino, A.; Xiccato, G. Animal welfare in reared rabbits: A review with emphasis on housing systems. World Rabbit Sci. 2006, 14, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Verga, M.; Luzi, F.; Petracci, M.; Cavani, C. Welfare aspects in rabbit rearing and transport. Ital. J. Anim. Sci. 2009, 8 (Suppl. 1), 191–204. [Google Scholar] [CrossRef]
- Benedek, I.; Altbӓcker, V.; Zsolnai, A.; Molnár, T. Exploring the genetic background of the differences in nest-building behavior in European rabbit. Animals 2020, 10, 1579. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P.; Oester, H.; Stauffacher, M. Effects of temporary nest box removal on maternal behaviour and pup survival in caged rabbits (Oryctolagus cuniculus). Appl. Anim. Behav. Sci. 2005, 91, 167–178. [Google Scholar] [CrossRef]
- Van De Weerd, H.A.; van Loo, P.L.P.; van Zutphen, L.F.; Koolhaas, J.M.; Baumans, V. Preferences for nesting material as environmental enrichment for laboratory mice. Lab. Anim. 1997, 3, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Percent Volume (Standard Error) | |
|---|---|
| Fur | 43 (12.2) |
| Plant material | 55 (14.7) |
| Other (eg. sand, litter) | 2 (3.8) |
| Percent Plant Material (Standard Error) | ||
|---|---|---|
| Dry | ||
| Agropyron | 15.3 | |
| Calamagrostis | 67.3 | |
| Carex | 13.3 | |
| Other | 3.5 | |
| Total dry | 84.4 (14.5) | |
| Green | ||
| Colchicum | 2.5 | |
| Hypnum c. | 7.4 | |
| Polygonatum o. | 3.2 | |
| Other | 2.5 | |
| Total green | 15.6 (14.5) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilkó, Á.; Petróczi, I.; Bárdos, B.; Nagy, I.; Altbacker, V. Composition of the Wild Rabbit Nest and Its Implication for Domestic Rabbit Breeding. Appl. Sci. 2022, 12, 1915. https://doi.org/10.3390/app12041915
Bilkó Á, Petróczi I, Bárdos B, Nagy I, Altbacker V. Composition of the Wild Rabbit Nest and Its Implication for Domestic Rabbit Breeding. Applied Sciences. 2022; 12(4):1915. https://doi.org/10.3390/app12041915
Chicago/Turabian StyleBilkó, Ágnes, Imre Petróczi, Boróka Bárdos, István Nagy, and Vilmos Altbacker. 2022. "Composition of the Wild Rabbit Nest and Its Implication for Domestic Rabbit Breeding" Applied Sciences 12, no. 4: 1915. https://doi.org/10.3390/app12041915
APA StyleBilkó, Á., Petróczi, I., Bárdos, B., Nagy, I., & Altbacker, V. (2022). Composition of the Wild Rabbit Nest and Its Implication for Domestic Rabbit Breeding. Applied Sciences, 12(4), 1915. https://doi.org/10.3390/app12041915