
Cellular Effects of Silver Nanoparticle Suspensions on Lung Epithelial Cells and Macrophages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Measurement of Mitochondrial Activity
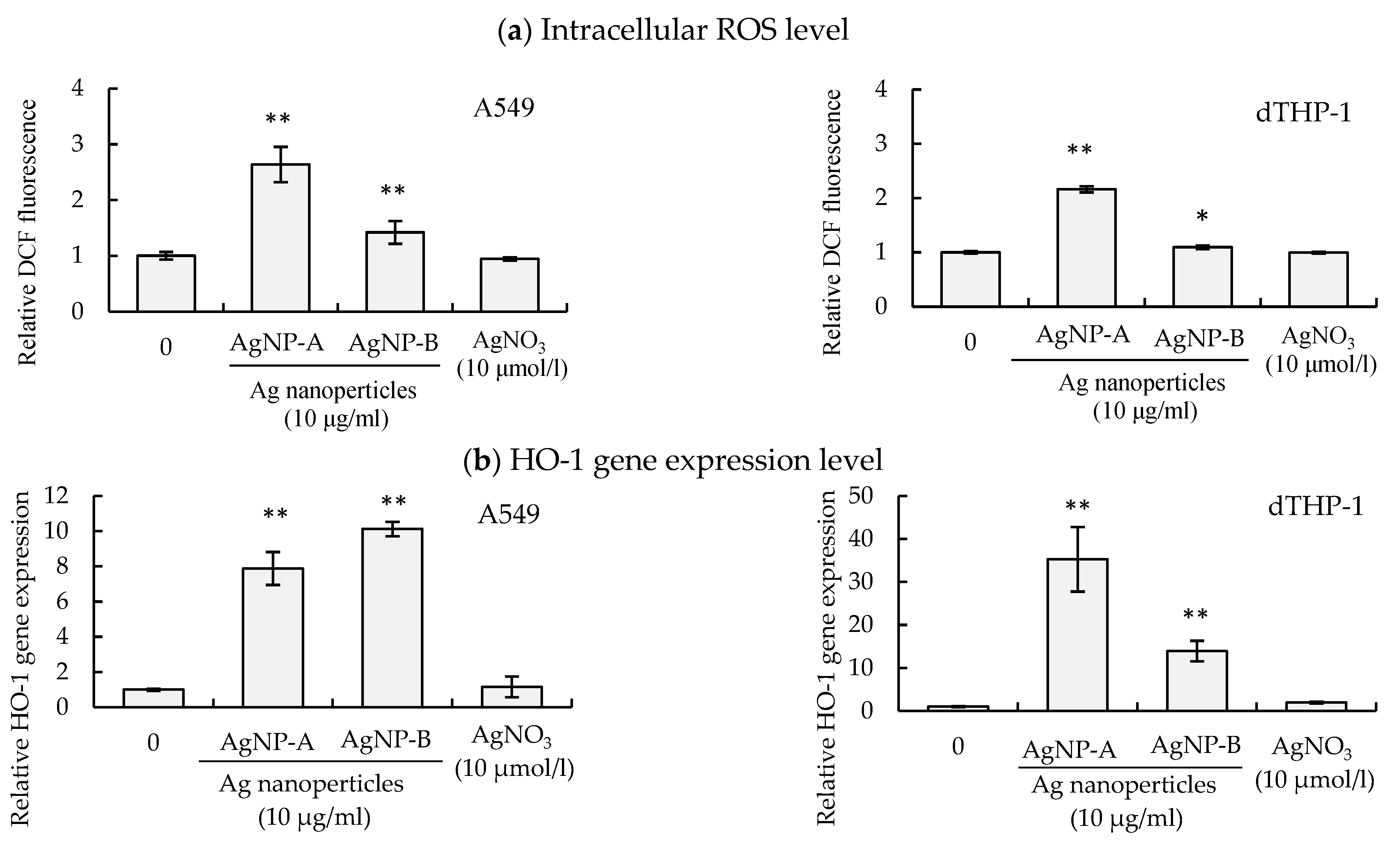
2.4. Measurement of Intracellular ROS Level
2.5. Determination of IL-8 Level in Culture Medium
2.6. Transmission Electron Microscopy (TEM)
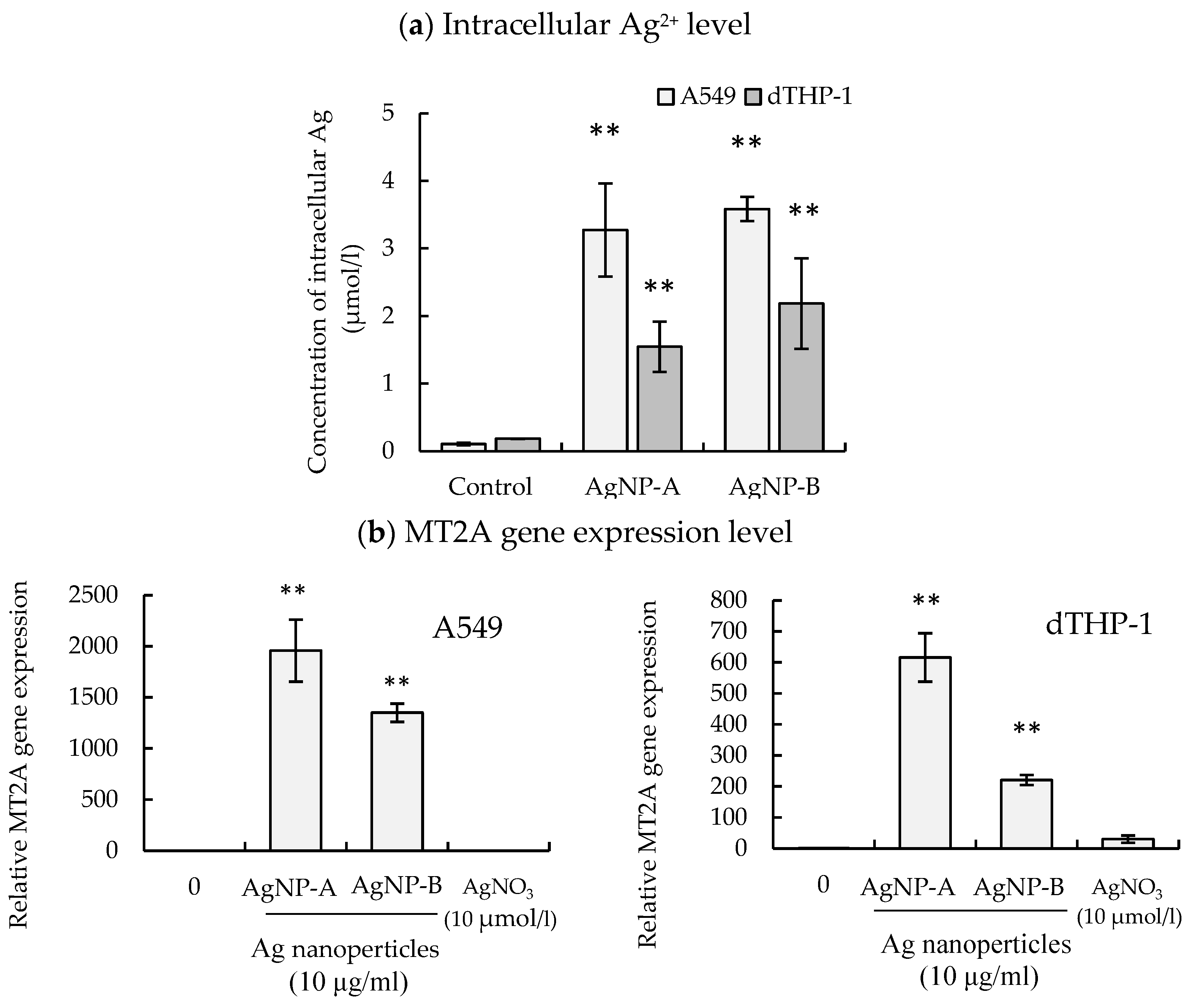
2.7. Gene Expression
2.8. Measurement of Silver Ions Released from AgNPs
2.9. Determination of Intracellular Silver Concentration
2.10. Statistical Analysis
3. Results
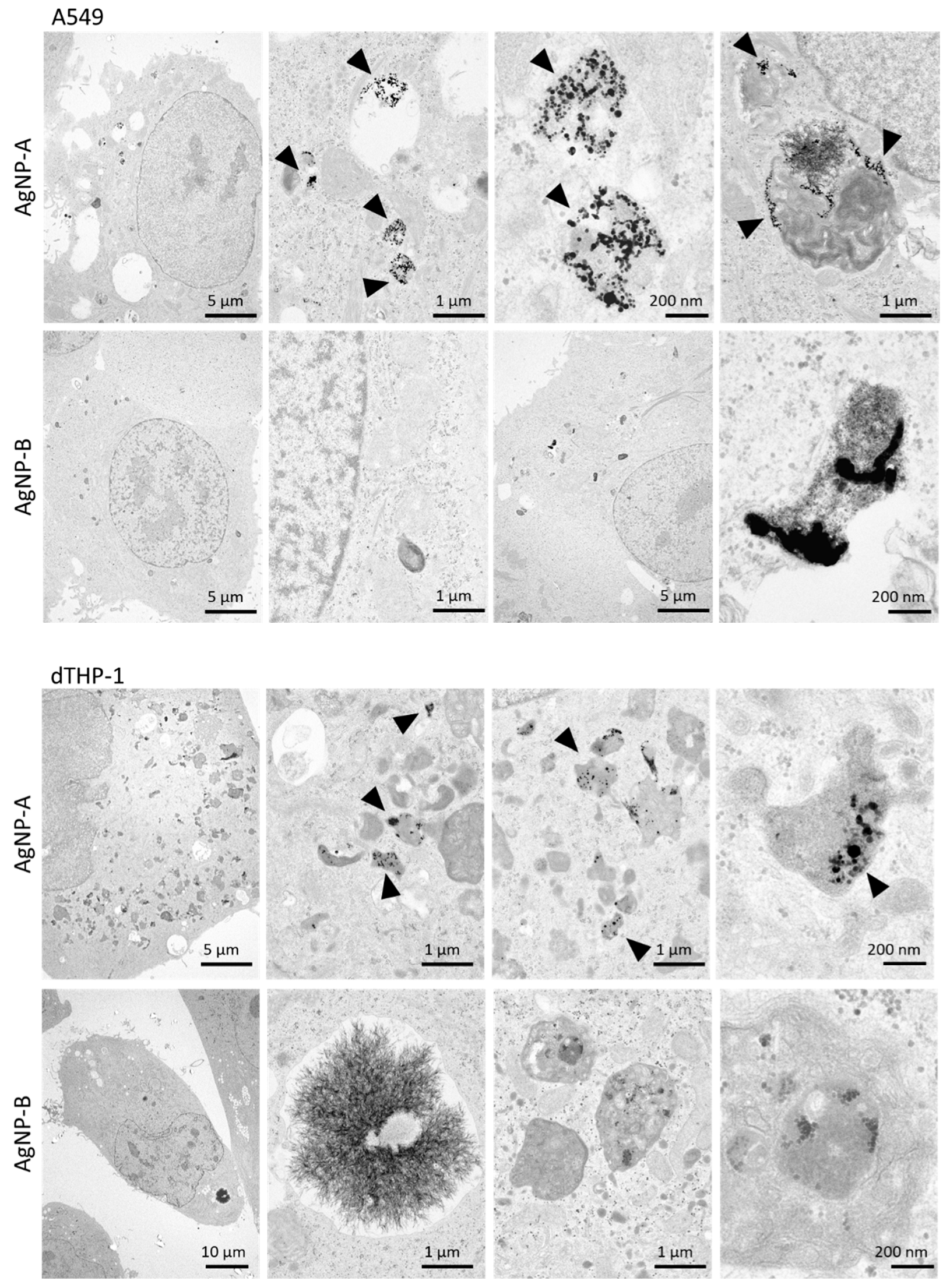
3.1. Cellular Uptake of AgNPs
3.2. Intracellular Silver Levels after Cell Exposure to a Silver Nanoparticle Suspension
3.3. Release of Silver Ions from AgNPs
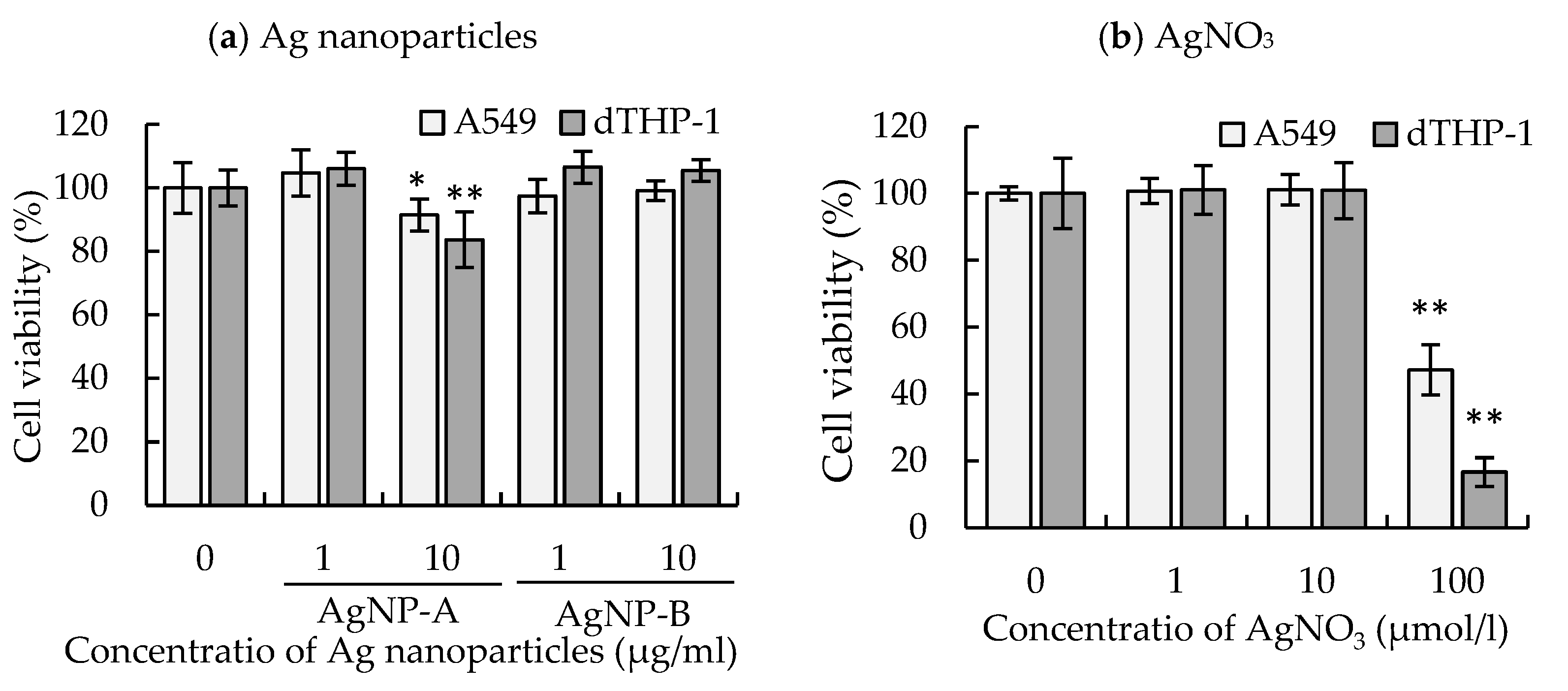
3.4. Effects of AgNP Suspension and AgNO3 Solution on Cell Viability
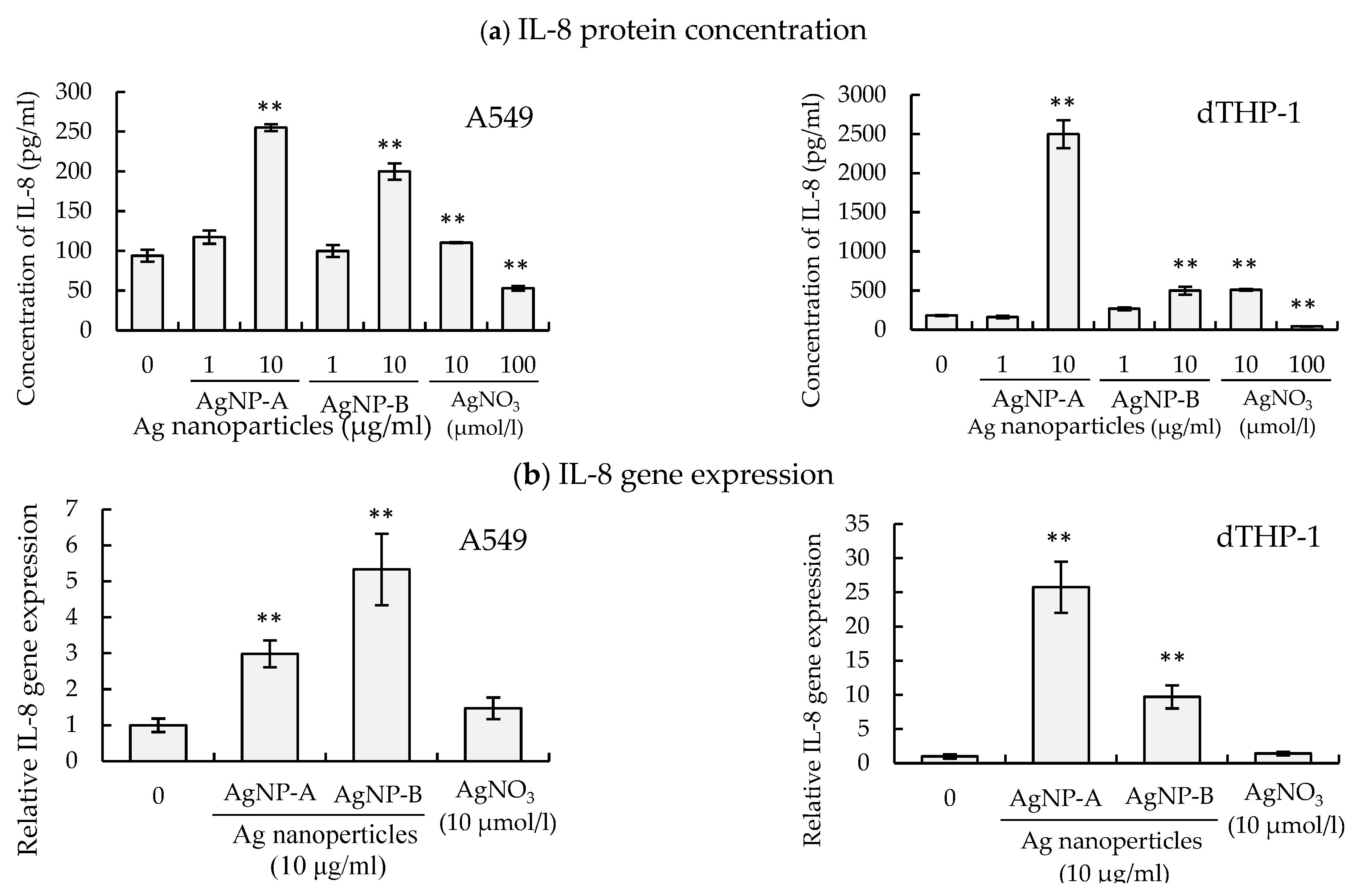
3.5. Regulation of IL-8 Expression by AgNP Suspension and AgNO3 Solution
3.6. Effects of AgNP Suspension and AgNO3 Solution on Oxidative Stress
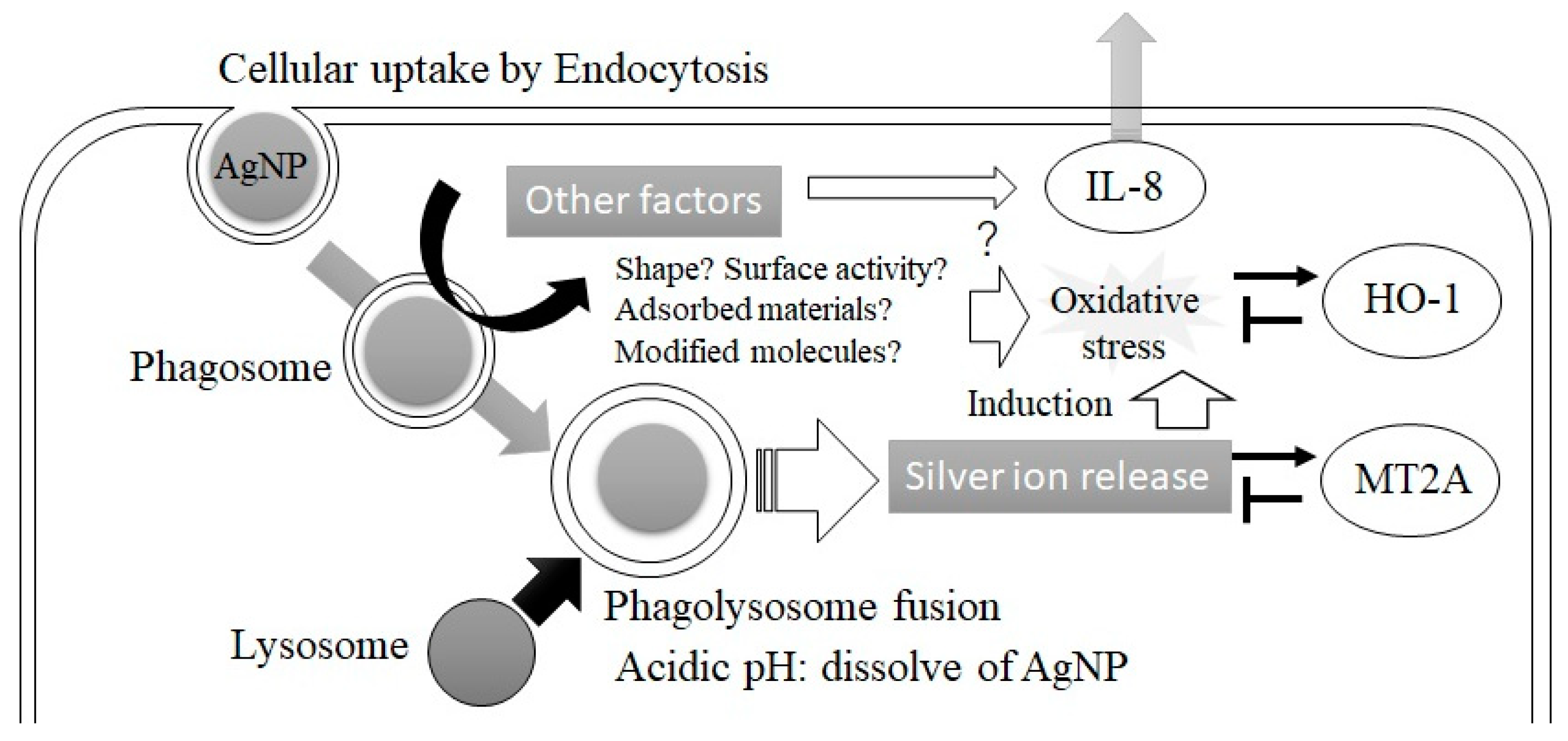
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Horie, M.; Iwahashi, H. The impact of the physiochemical properties of manufactured nanoparticles on in vitro and in vivo evaluation of particle toxicity. J. Phys. Chem. Biophys. 2014, 4, 2. [Google Scholar] [CrossRef]
- Kim, J.S.; Kuk, E.; Yu, K.N.; Kim, J.H.; Park, S.J.; Lee, H.J.; Kim, S.H.; Park, Y.K.; Park, Y.H.; Hwang, C.Y.; et al. Antimicrobial effects of silver nanoparticles. Nanomedicine 2007, 1, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Keat, C.L.; Aziz, A.; Eid, A.M.; Elmarzugi, N.A. Biosynthesis of nanoparticles and silver nanoparticles. Bioresour. Bioprocess. 2015, 2, 47. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Li, Y.; Tjong, S.C. Bactericidal and Cytotoxic Properties of Silver Nanoparticles. Int. J. Mol. Sci. 2019, 20, 449. [Google Scholar] [CrossRef] [Green Version]
- Velmurugan, P.; Lee, S.-M.; Cho, M.; Park, J.-H.; Seo, S.-K.; Myung, H.; Bang, K.-S.; Oh, B.-T. Antibacterial activity of silver nanoparticle-coated fabric and leather against odor and skin infection causing bacteria. Appl. Microbiol. Biotechnol. 2014, 98, 8179–8189. [Google Scholar] [CrossRef]
- Tian, J.; Wong, K.K.; Ho, C.M.; Lok, C.N.; Yu, W.Y.; Che, C.M.; Cliu, J.F.; Tam, P.K. Topical delivery of silver nanoparticles promotes wound healing. ChemMedChem 2007, 2, 129–136. [Google Scholar] [CrossRef]
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. Nano Lett. 2012, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Durán, N.; Durán, M.; de Jesus, M.B.; Seabra, A.B.; Fávaro, W.J.; Nakazato, G. Silver nanoparticles: A new view on mechanistic aspects on antimicrobial activity. Nanomedicine 2016, 12, 789–799. [Google Scholar] [CrossRef]
- Marambio-Jones, C.; Hoek, E.M.V. A review of the antibacterial effects of silver nanomaterials and potential implications for human health and the environment. J. Nanopart. Res. 2010, 12, 1531–1551. [Google Scholar] [CrossRef]
- Russell, A.D.; Hugo, W.B. 7 Antimicrobial Activity and Action of Silver. Prog. Med. Chem. 1994, 31, 351–370. [Google Scholar]
- Zhang, X.-F.; Shen, W.; Gurunathan, S. Silver Nanoparticle-Mediated Cellular Responses in Various Cell Lines: An In Vitro Model. Int. J. Mol. Sci. 2016, 17, 1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Kim, D.W.; Lee, Y.H.; Oh, J.H.; Yoon, S.; Choi, M.S.; Lee, S.K.; Kim, J.W.; Lee, K.; Song, C.W. Silver nanoparticles induce apoptosis and G2/M arrest via PKCζ-dependent signaling in A549 lung cells. Arch. Toxicol. 2011, 85, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, M.J.; Lee, S.Y.; Oh, S.M.; Chung, K.H. Genotoxic effects of silver nanoparticles stimulated by oxidative stress in human normal bronchial epithelial (BEAS-2B) cells. Mutat. Res. Toxicol. Environ. Mutagen. 2011, 726, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Iram, S.; Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Ahmad, K.; Khan, M.S.; Kim, J. Fruit Derived Potentially Bioactive Bioengineered Silver Nanoparticles. Int. J. Nanomed. 2021, 16, 7711–7726. [Google Scholar] [CrossRef]
- Syed, A.; Saraswati, S.; Kundu, G.C.; Ahmad, A. Biological synthesis of silver nanoparticles using the fungus Humicola sp. and evaluation of their cytoxicity using normal and cancer cell lines. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 114, 144–147. [Google Scholar] [CrossRef]
- Horie, M.; Nishio, K.; Fujita, K.; Kato, H.; Nakamura, A.; Kinugasa, S.; Endoh, S.; Miyauchi, A.; Yamamoto, K.; Murayama, H.; et al. Ultrafine NiO Particles Induce Cytotoxicity in Vitro by Cellular Uptake and Subsequent Ni(II) Release. Chem. Res. Toxicol. 2009, 22, 1415–1426. [Google Scholar] [CrossRef]
- Cronholm, P.; Karlsson, H.L.; Hedberg, J.; Lowe, T.A.; Winnberg, L.; Elihn, K.; Wallinder, I.O.; Möller, L. Intracellular Uptake and Toxicity of Ag and CuO Nanoparticles: A Comparison Between Nanoparticles and their Corresponding Metal Ions. Small 2013, 9, 970–982. [Google Scholar] [CrossRef]
- Horie, M.; Kato, H.; Fujita, K.; Endoh, S.; Iwahashi, H. In VitroEvaluation of Cellular Response Induced by Manufactured Nanoparticles. Chem. Res. Toxicol. 2011, 25, 605–619. [Google Scholar] [CrossRef]
- Tabei, Y.; Sonoda, A.; Nakajima, Y.; Biju, V.; Makita, Y.; Yoshida, Y.; Horie, M. Intracellular accumulation of indium ions released from nanoparticles induces oxidative stress, proinflammatory response and DNA damage. J. Biochem. 2015, 159, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Fukui, H.; Horie, M.; Endoh, S.; Kato, H.; Fujita, K.; Nishio, K.; Komaba, L.K.; Maru, J.; Miyauhi, A.; Nakamura, A.; et al. Association of zinc ion release and oxidative stress induced by intratracheal instillation of ZnO nanoparticles to rat lung. Chem. Interact. 2012, 198, 29–37. [Google Scholar] [CrossRef]
- Karlsson, H.L.; Cronholm, P.; Gustafsson, J.; Möller, L. Copper Oxide Nanoparticles Are Highly Toxic: A Comparison between Metal Oxide Nanoparticles and Carbon Nanotubes. Chem. Res. Toxicol. 2008, 21, 1726–1732. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lim, D.-H.; Lim, H.-J.; Kwon, T.; Choi, J.-S.; Jeong, S.; Choi, I.-H.; Cheon, J. Size dependent macrophage responses and toxicological effects of Ag nanoparticles. Chem. Commun. 2011, 47, 4382–4384. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, V.; Cornaghi, L.; Gianazza, E.; Potenza, M.A.; Donetti, E.; Marinovich, M.; Corsini, E. In vitro assessment of silver nanoparticles immunotoxicity. Food Chem. Toxicol. 2018, 112, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Shimizu, K.; Tabei, Y. Validation of metallothionein, interleukin-8, and heme oxygenase-1 as markers for the evaluation of cytotoxicity caused by metal oxide nanoparticles. Toxicol. Mech. Methods 2018, 28, 630–638. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Khan, M.M.; Alrokayan, S.; Alhadlaq, H. Oxidative stress mediated cytotoxicity and apoptosis response of bismuth oxide (Bi2O3) nanoparticles in human breast cancer (MCF-7) cells. Chemosphere 2019, 216, 823–831. [Google Scholar] [CrossRef]
- Moratin, H.; Scherzad, A.; Gehrke, T.; Ickrath, P.; Radeloff, K.; Kleinsasser, N.; Hackenberg, S. Toxicological characterization of ZnO nanoparticles in malignant and non-malignant cells. Environ. Mol. Mutagen. 2018, 59, 247–259. [Google Scholar] [CrossRef]
- Siddiqui, M.; Alhadlaq, H.; Ahmad, J.; Al-Khedhairy, A.; Musarrat, J.; Ahamed, M. Copper Oxide Nanoparticles Induced Mitochondria Mediated Apoptosis in Human Hepatocarcinoma Cells. PLoS ONE 2013, 8, e69534. [Google Scholar] [CrossRef] [Green Version]
- Ahamed, M.; Ali, D.; Alhadlaq, H.; Akhtar, M. Nickel oxide nanoparticles exert cytotoxicity via oxidative stress and induce apoptotic response in human liver cells (HepG2). Chemosphere 2013, 93, 2514–2522. [Google Scholar] [CrossRef]
- Mittal, S.; Pandey, A.K. Cerium oxide nanoparticles induced toxicity in human lung cells: Role of ROS mediated DNA damage and apoptosis. Biomed. Res. Int. 2014, 2014, 891934. [Google Scholar] [CrossRef]
- Tang, J.; Lu, X.; Chen, B.; Cai, E.; Liu, W.; Jiang, J.; Chen, F.; Shan, X.; Zhang, H. Mechanisms of silver nanoparticles-induced cytotoxicity and apoptosis in rat tracheal epithelial cells. J. Toxicol. Sci. 2019, 44, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, C.; Ariyoshi, T.; Kawaguchi, H.; Nagasaka, S.; Kashiwada, S. Salinity increases the toxicity of silver nanocolloids to Japanese medaka embryos. Environ. Sci. Nano 2015, 2, 94–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code in This Study | Primary Particle Size (nm) | Included Chemicals in the Suspension | Application | Manufacturer |
|---|---|---|---|---|
| AgNP-A | 28.4 ± 8.5 1 | Nothing | Cosmetically suppliment | Utopia Silver Supplements |
| AgNP-B | 20 | Nitrogen containing compound | Industrial application | Tokuriki Honten Co., Ltd. |
| Acidic Condition 1 | DMEM with 10% FBS | ||
|---|---|---|---|
| Rereased Ag2+ | AgNP-A | 22.2 | 5.6 |
| (μmol/L) | AgNP-B | 7.4 | 4.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, K.; Kashiwada, S.; Horie, M. Cellular Effects of Silver Nanoparticle Suspensions on Lung Epithelial Cells and Macrophages. Appl. Sci. 2022, 12, 3554. https://doi.org/10.3390/app12073554
Shimizu K, Kashiwada S, Horie M. Cellular Effects of Silver Nanoparticle Suspensions on Lung Epithelial Cells and Macrophages. Applied Sciences. 2022; 12(7):3554. https://doi.org/10.3390/app12073554
Chicago/Turabian StyleShimizu, Kaori, Shosaku Kashiwada, and Masanori Horie. 2022. "Cellular Effects of Silver Nanoparticle Suspensions on Lung Epithelial Cells and Macrophages" Applied Sciences 12, no. 7: 3554. https://doi.org/10.3390/app12073554
APA StyleShimizu, K., Kashiwada, S., & Horie, M. (2022). Cellular Effects of Silver Nanoparticle Suspensions on Lung Epithelial Cells and Macrophages. Applied Sciences, 12(7), 3554. https://doi.org/10.3390/app12073554