
Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both?



Abstract
:1. Introduction
2. Material and Methods
2.1. Fish and Environment
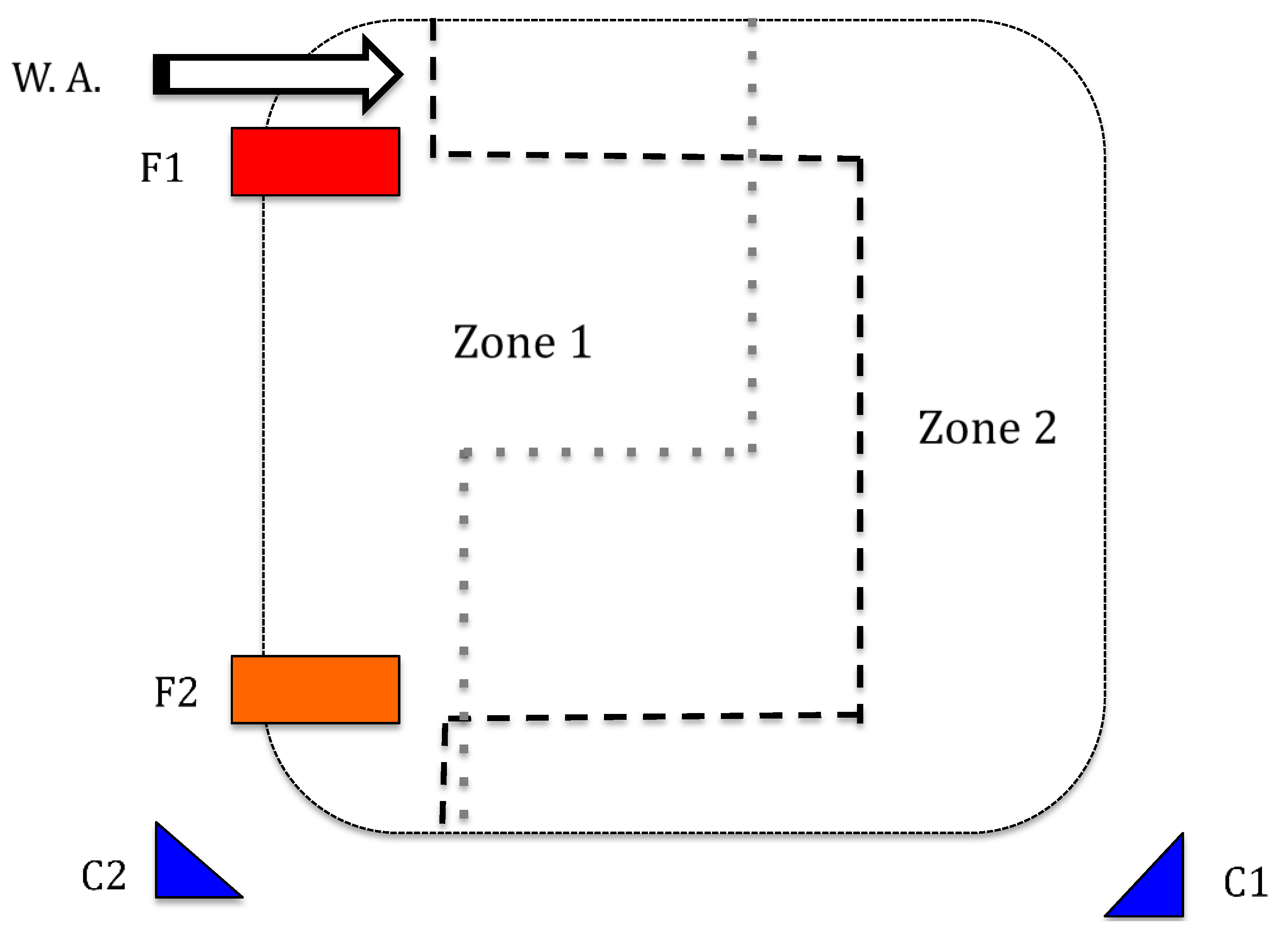
2.2. Experimental Design
2.3. Physical Enrichment
2.4. Behavioural Parameters
- Particular swimming. Two different behaviours were observed: (i) the fish swam on the back (SB) with the ventral body above, and (ii) the fish swam vertically (SV) with the head above the water surface.
- Jumps (J) were also observed; the fish was out of the water with its entire body above the surface. This behaviour was observed mainly along the wall of the tank.
2.5. Growth Characteristic Assessment
2.6. Statistical Analyses
3. Results
3.1. Survival
3.2. Space Occupation
3.3. Behaviours
3.4. Growth of the Fish
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action Rome; FAO: Rome, Italy, 2020; 401p. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current issues in fish welfare. J. Fish Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef] [Green Version]
- Arechavala-Lopez, P.; Cabrera-Alvarez, M.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2021, 14, 704–728. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Kientz, J.L.; Barnes, M.E. Structural complexity improves the rearing performance of rainbow trout in circular tanks. N. Am. J. Aquac. 2016, 78, 203–207. [Google Scholar] [CrossRef]
- Kientz, J.L.; Crank, K.M.; Barnes, M.E. Enrichment of circular tanks with vertically suspended strings of colored balls improves rainbow trout rearing performance. N. Am. J. Aquac. 2018, 80, 162–167. [Google Scholar] [CrossRef]
- Crank, K.M.; Kientz, J.L.; Barnes, M.E. An evaluation of vertically suspended environmental structures during rainbow trout rearing. N. Am. J. Aquac. 2019, 81, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, A.J.; Fletcher, B.L.; Barnes, M.E.; Treft, C.E.; Bursell, B.R. Vertically suspended environmental enrichment structures improve the growth of juvenile landlocked fall Chinook salmon. Int. J. Innov. Stu. Aquat. Biol. Fish. 2019, 5, 17–24. [Google Scholar] [CrossRef]
- White, S.C.; Barnes, M.E.; Krebs, E.; Huysman, N.; Voorhees, J.M. Addition of vertical enrichment structures does not improve growth of three salmon species during hatchery rearing. J. Marine Biol. Aquac. 2018, 4, 48–52. [Google Scholar]
- Ebbessson, L.O.E.; Braithwaite, V.A. Environmental effects on fish neural plasticity and cognition. J. Fish Biol. 2012, 81, 2151–2174. [Google Scholar] [CrossRef]
- Berejikian, B.A.; Tezak, E.R.; Flagg, T.A.; LaRae, A.L.; Kummerow, E.; Mahnken, C.V.W. Social dominance, growth and habitat use of Age-0 steelhead (Oncorhynchus mykiss) grown in enriched and conventional hatchery rearing environments. C. J. Fish Aquat. Sci. 2000, 57, 628–636. [Google Scholar] [CrossRef]
- Brown, C.; Davidson, T.; Laland, K. Environmental enrichment and prior experience of live prey improve foraging behaviour in hatchery-reared Atlantic salmon. J. Fish Biol. 2003, 63, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Braithwaite, V.A.; Salvanes, A.G.V. Environmental variability in the early rearing environment generates behaviourally flexible cod: Implications for reability wild populations. Proc. R. Soc. B 2005, 272, 1107–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlbeck-Bergendahl, I.; Salvanes, A.G.V.; Braithwaite, V.A. Determining the effects of duration and recency of exposure environmental enrichment. Appl. Anim. Behav. Sci. 2016, 176, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Zuberi, A.; Khan, K.U.; Ahmad, S.; Thornquist, P.; Winberg, S. Effects of enrichment of the development of behaviour of an endangered fish masheer (Tor putitora). Appl. Anim. Behav. Sci. 2017, 186, 93–100. [Google Scholar] [CrossRef]
- Salvanes, A.G.; Moberg, O.; Ebbesson, L.O.; Nilsen, T.O.; Jensen, K.H.; Braithwaite, V.A. Environmental enrichment promotes neural plasticity and cognitive abilities in fish. Proc. Biol. Sci. Royal Soc. 2013, 280, 20131331. [Google Scholar] [CrossRef]
- Carfagnini, A.G.; Rodd, F.H.; Jeffers, K.B.; Bruce, A.E.E. The effects of habitat complexity on aggression and fecundity in zebrafish (Danio rerio). Environ. Biol. Fishes 2009, 86, 403–409. [Google Scholar] [CrossRef]
- Mikheev, V.N.; Pasternak, A.F.; Tischler, G.; Wanzenbock, J. Contestable shelters provoke aggression among 0+ perch, Perca fluviatilis. Environ. Biol. Fishes 2005, 73, 227–231. [Google Scholar] [CrossRef]
- Gwak, W.S. Effects of shelter on growth and survival in age-0 black sea bass, Centropristis striata (L.). Aquac. Res. 2003, 34, 1387–1390. [Google Scholar] [CrossRef]
- Christou, P.; Savin, R.; Costa-Pierce, B.A.; Misztal, I.; Whitelaw, C.B.A. Sustainable Food Production; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Kozlowski, M.; Szczepkowski, M.; Wunderlich, K.; Szczepkowska, B.; Piotrowska, I. Polyculture of Juveniles pikeperch (Sander lucioperca (L.)) and sterlet (Acipencer ruthenus L.) in a recirculating system. Arch. Polish Fish 2014, 22, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.; Lecocq, T.; Abregal, C.; Nahon, S.; Aubin, J.; Jaeger, C.; Wilfart, A.; Schaeffer, L.; Ledoré, Y.; Puillet, L.; et al. The effects of polyculture on behaviour and production of pikeperch in recirculation systems. Aquac. Rep. 2020, 17, 100333. [Google Scholar] [CrossRef]
- Martins, C.I.M.; Galhardo, L.; Noble, C.; Damsgård, B.; Spedicato, M.T.; Zupa, W.; Beauchaud, M.; Kulczykowska, E.; Massabuau, J.C.; Carter, T.; et al. Behavioural indicators of welfare in farmed fish. Fish Physiol. Biochem. 2012, 38, 17–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Näslund, J.; Rosengren, M.; Del Villar, D.; Gansel, L.; Norrgard, J.R.; Persson, L.; Winkowski, J.J.; Kvingedal, E. Hatchery tank enrichment affects cortisol levels and shelter-seeking in Atlantic Salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2013, 70, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, X.; Li, Z.; Zhang, X. Effects of different level of environmental enrichment on the sheltering behaviors, brain development and cortisol levels of black rockfish Sebastes schlegelli. Appl. Anim. Behav. Sci. 2019, 218, 104825. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, Q.; Xu, X.; Guo, H.; Zhang, X. Effects of environmental enrichment on the welfare of juvenile black rockfish Sebastes schlegelli. Growth, behaviour and physiology. Aquaculture 2020, 518, 734782. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, X.; Wang, Y.; Zhang, X. Effects of environmental enrichment on growth performance, aggressive behaviour and stress-induced changes in cortisol release and neurogenesis of black rockfish Sebastes schlegelli. Aquaculture 2020, 528, 735483. [Google Scholar] [CrossRef]
- Zhang, Z.; Fu, Y.; Zhang, Z.; Zhang, X.; Chen, S. A comparative study on two territorial fishes: The influence of physical enrichment an aggressive behaviour. Anim. Open Access J. 2021, 11, 1868. [Google Scholar] [CrossRef]
- Mork, O.I.; Bjerkeng, B.; Rye, M. Aggressive interactions in pure and mixed groups of juveniles farms and hatchery-reared wild Atlantic salmon Salmo salar L. in relation to tank substrate. Aquac. Res. 1999, 30, 571–578. [Google Scholar] [CrossRef]
- Basquill, S.P.; Grant, J.W.A. An increase in habitat complexity reduces aggression and monopolization of food by zebra fish (Danio rerio). Can. J. Fish Aquat. Sci. 1998, 76, 770–772. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, L.; Owen, S.F.; Readman, G.D.; Sloman, K.A.; Wilson, R.W. Does structural enrichment for toxicology studies improve zebrafish welfare. Appl. Anim. Behav. Sci. 2012, 139, 143–150. [Google Scholar] [CrossRef]
- Bosakowski, T.; Wagner, E.J. Experimental use of cobble substrates in concrete raceways for improving fin condition of cutthroat (Oncorhynchus clarki) and rainbow trout (Oncorhynchus mykiss). Aquaculture 1995, 130, 159–165. [Google Scholar] [CrossRef]
- Wagner, E.I.; Intelmann, S.S.; Routledge, M.D. The effect of fry rearing density on hatchery performance, fin condition, and agonistic behaviour of rainbow trout Oncorhynchus mykiss. J. World. Aquac. Soc. 1996, 27, 264–274. [Google Scholar] [CrossRef]
- Arndt, R.E.; Routledge, D.; Wagner, E.J.; Mellenthin, R.F. Influence of raceway substrate and design on fin erosion and hatchery performance of rainbow trout. N. Am. J. Aquac. 2001, 63, 312–320. [Google Scholar] [CrossRef]
- Berejikian, B.A.; Tezak, E.P. Rearing in enriched hatchery tanks improves dorsal fin quality of juvenile steelhead. N. Am. J. Aquac. 2005, 65, 289–293. [Google Scholar] [CrossRef]
- Coulibaly, A.; Ouattara, I.N.; Koné, T.; Douba, V.N.; Snoeks, J.; Gooré Bi, G.; Kouamélian, E.P. First results of floating cage culture of the African catfish Heterobranchus longifilis Valenciennes 1840: Effect of stocking density on survival and growth rates. Aquaculture 2007, 263, 61–67. [Google Scholar] [CrossRef]
- Gesto, M.; Jokumsen, A. Effects of shelter on growth performance and welfare of rainbow trout juveniles. Aquaculture 2022, 551, 737930. [Google Scholar] [CrossRef]
- Barcellos, L.J.G.; Kreutz, L.C.; Quevedo, R.M.; Da Rosa, J.G.S.; Koakoski, G.; Centenaro, L.; Pottker, E. Influence of color backround and shelter availability on jundia (Rhamdia quelen) stress response. Aquaculture 2009, 288, 51–56. [Google Scholar] [CrossRef]
- Millidine, K.J.; Amstrong, J.D.; Metcalfe, N.B. Presence of shelter reduces maintenance metabolism of juvenile salmon. Funct. Ecol. 2006, 20, 839–845. [Google Scholar] [CrossRef]
- Hecht, T.; Appelbaum, S. Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Clarias gariepinus (Clariidae: Pisces) under controlled conditions. J. Zool. 1988, 214, 21–44. [Google Scholar] [CrossRef]
- Rahmah, S.; Kato, K.; Yamamoto, S.; Takii, K.; Murata, O.; Senoo, S. Improved survival and growth performances with stocking density manipulation and shelter availability in bagrid catfish Mystus nemurus (Cuvier & Valenciennes 1840) larvae. Aquac. Res. 2014, 45, 2000–2009. [Google Scholar] [CrossRef]
- Mihailov, S.A.; Mihoc, N.A.; Lalescu, D.; Grozea, A. Polyculture of the pikeperch (Sander lucioperca) fingerlings into recirculating aquaculture system, with sterlet (Acipenser ruthenus) or european catfish (Silurus glanis). A preliminaty study. Res. J. Agri. Sci. 2017, 49, 193–198. [Google Scholar]
- Prokes, M.; Barus, V.; Mares, J.; Penaz, M.; Baranek, V. Growth of sterlet Acipencer ruthenus under experimental and farm conditions of the Czeck republic with remarks on other sturgeons. Acta Univ. Agric. Sylvi. Mendel. Brun. 2011, 59, 81–290. [Google Scholar]
- Baekelandt, S.; Redivo, B.; Mandiki, S.N.M.; Bournonville, T.; Houndji, A.; Bernard, B.; El Kertaoul, N.; Schmitz, M.; Fontaine, P.; Gardeur, J.N.; et al. Multifactorial analysis revealed optimal aquaculture modalities improving husbandry fitness without clear effect on stress and immune status of pikeperch Sander lucioperca. Gen. Comp. Endocrinol. 2018, 258, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, B.; Feledi, T.; Lengyel, S.; Ronyal, A. Effect of feeding rate on growth performance, food utilization and meat yield of sterlet (Acipencer ruthenus Linné, 1758). J. Fish Sci. 2013, 7, 216–224. [Google Scholar] [CrossRef]
- Bayrami, A.; Noverian, H.A.; Sharif, E.A. Effects of backround colour on growth indexes and stress of young sterlet (Acipencer ruthenus) in a closed circulated system. Aquac. Res. Dev. 2017, 48, 2004–2011. [Google Scholar] [CrossRef]
- Luchiari, A.C.; De Morais Freire, F.A.; Koskela, J.; Pirhonen, J. Light intensity preference of juvenile pikeperch Sander lucioperca. Aquac. Res. 2006, 37, 1572–1577. [Google Scholar] [CrossRef]
- Moine, J.; Barnes, M.E.; Kientz, J.; Simpson, G. Flow patterns in circular rearing tanks containing vertical structures. J. Fish. Livest. Prod. 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Fieszl, J.; Bogacka-Kapusta, E.; Kapusta, A.; Szymanska, U.; Martyniak, A. Feeding ecology of the sterlet Acipenser ruthenus L. in the Hungarian section of the Danube river. Arch. Pol. Fish. 2011, 19, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Vehanen, T.; Lathi, M. Movements and habitat use by pikeperch (Stizostedion lucioperca (L.)) in a hydropeaking reservoir. Ecol. Freswater Fish 2003, 12, 203–215. [Google Scholar] [CrossRef]
- Giraudoux, P. Pgirmess: Data Analysis in Ecology. R Package Version 1.5.8. 2013. Available online: http://CRAN.R-project.org/package=pgirmess (accessed on 17 January 2022).
- Colchen, T.; Gisbert, E.; Ledoré, Y.; Teletchea, F.; Fontaine, P.; Pasquet, A. Is a cannibal different from its conspecifics? A behavioural, morphological, muscular and retinal structure study with pikeperch juveniles under farming conditions. Appl. Anim. Behav. 2020, 224, 104947. [Google Scholar] [CrossRef]
- Kotrschal, A.; Taborsky, B. Environmental change enhances cognitive abilities in fish. PLoS ONE 2010, 8, e1000351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, R.; Magurran, A.E.; Smith, C.L. Spatial cognition in zebrafish: The role of strain and rearing environment. Anim. Cogni. 2011, 14, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Hazegawa, K.; Maekawa, K. Potential of habitat complexity for mitigating interference competition between native and non-native salmonidae species. Can. J. Fish Aquat. Sci. 2008, 86, 386–393. [Google Scholar] [CrossRef]
- Woodward, M.A.; Winder, L.A.; What, P.J. Enrichment increases aggression in zebrafish. Fishes 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Krebs, E.; Huysman, N.; Voorhes, J.M.; Barnes, M. Suspended arrays improve rainbow trout growth during hatchery rearing in circular tanks. Int. J. Aquac. Fish. Sci. 2018, 4, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, J.M.; Huysman, N.; Krebs, E.; Barnes, M.E. Influence of water velocity and vertically suspended structures on rainbow trout rearing performance. Open J. Anim. Sci. 2020, 10, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Huysman, N.; Krebs, E.; Voorhees, J.M.; Barnes, M.E. Use of large vertically-suspended rod array in circular tanks during juvenile rainbow trout rearing. Int. J. Mar. Biol. Res. 2019, 4, 1–5. [Google Scholar]
- Slavík, O.; Maciak, M.; Horký, P. Shelter use of familiar and unfamiliar groups of juvenile European catfish Silurus glanis. Appl. Anim. Behav. Sci. 2012, 142, 116–123. [Google Scholar] [CrossRef]
- Broom, D.M.; Johnson, K.G. Stress and Animal Welfare, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Pounder, K.C.; Mitchell, J.L.; Thomson, J.S.; Pottinger, T.G.; Buckley, J.; Sneddon, L.U. Does environmental enrichment promote recovery from stress in rainbow trout? Appl. Anim. Behav. Sci. 2016, 176, 136–142. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enrichment (E) | ||||||||
| Species | Wi (g) | FCFi | Wf (g) | FCFf | BMi (g) | BMf (g) | BG (%) | SGR (%/day) |
| Sterlet | 133.1 ± 21.2 | 0.40 ± 0.00 | 324.7 ± 65.2 | 0.4 ± 0.0 | 3993.2 ± 17.6 | 7472.2 ± 674.3 | 87 ± 17.5 | 0.99 ± 0.08 |
| Pikeperch | 142.0 ± 38.1 | 0.7 ± 0.1 | 381.9 ± 88.4 | 0.8 ± 0.1 | 6373.6 ± 39.7 | 14763.5 ± 303.1 | 131.6 ± 3.4 | 1.10 ± 0.03 |
| No Enrichment (NE) | ||||||||
| Species | Wi (g) | FCFi | Wf (g) | FCFf | BMi (g) | BMf (g) | BG (%) | SGR (%/day) |
| Sterlet | 132.6 ± 21.1 | 0.38 ± 0.04 | 337.1 ± 51.4 | 0.46 ± 0.05 | 3979.0 ± 11.7 | 7434.5 ± 941.6 | 87 ± 23.2 | 1.04 ± 0.05 |
| Pikeperch | 143.4 ± 44.6 | 0.7 ± 0.1 | 369.7 ± 94.0 | 0.8 ± 0.1 | 6452.4 ± 92.5 | 14412.0 ± 617.0 | 123.3 ± 7.8 | 1.05 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, M.; Reynaud, J.-G.; Ledoré, Y.; Pasquet, A.; Lecocq, T. Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both? Appl. Sci. 2022, 12, 3674. https://doi.org/10.3390/app12073674
Thomas M, Reynaud J-G, Ledoré Y, Pasquet A, Lecocq T. Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both? Applied Sciences. 2022; 12(7):3674. https://doi.org/10.3390/app12073674
Chicago/Turabian StyleThomas, Marielle, Jean-Gabriel Reynaud, Yannick Ledoré, Alain Pasquet, and Thomas Lecocq. 2022. "Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both?" Applied Sciences 12, no. 7: 3674. https://doi.org/10.3390/app12073674
APA StyleThomas, M., Reynaud, J. -G., Ledoré, Y., Pasquet, A., & Lecocq, T. (2022). Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both? Applied Sciences, 12(7), 3674. https://doi.org/10.3390/app12073674