

Cosmeceutical Potentials of Equisetum debile Roxb. ex Vaucher Extracts

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Materials
2.3. Plant Extraction Procedures
2.3.1. Preparation of Crude Extracts
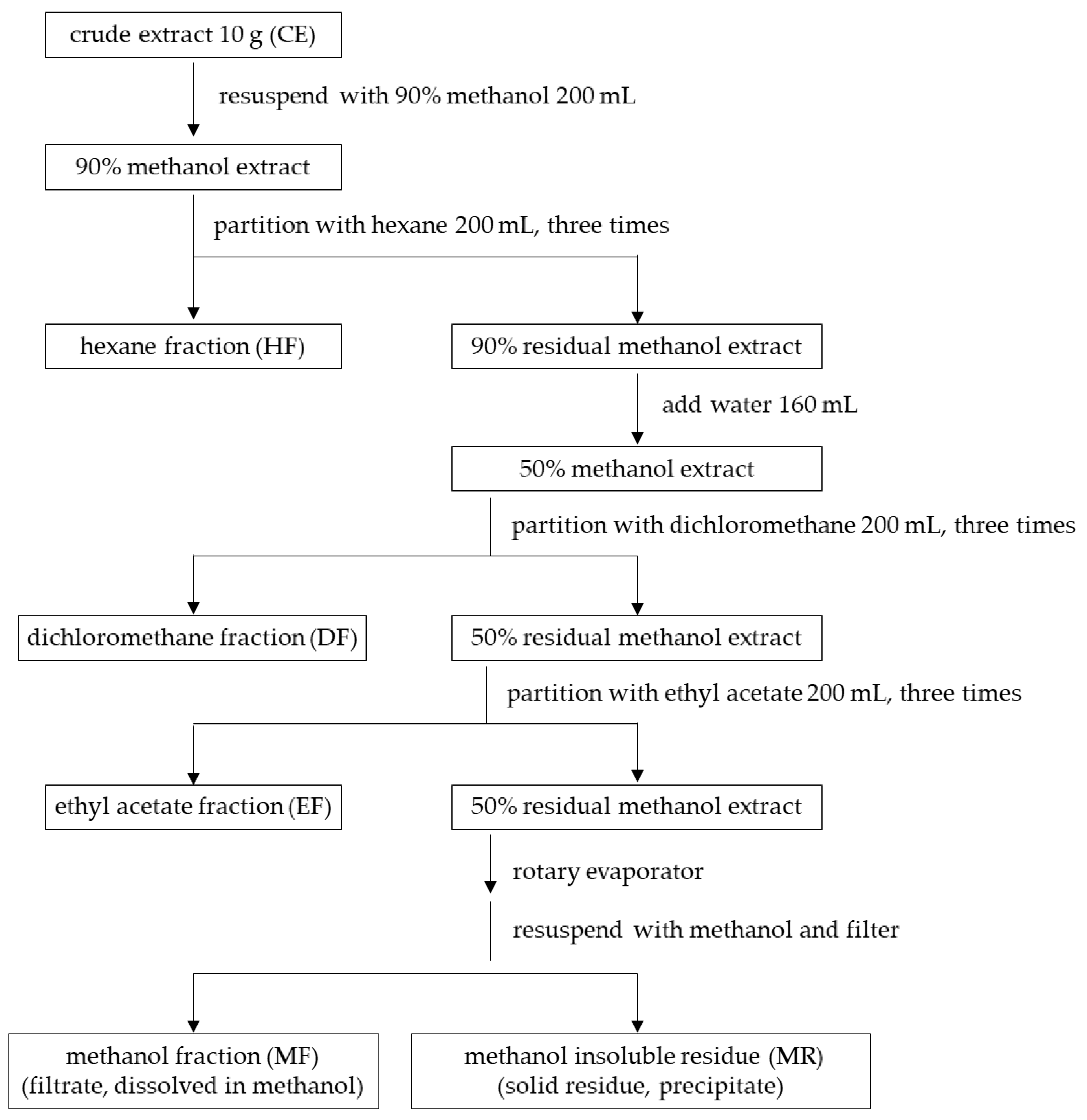
2.3.2. Isolation of Semi-Purified Fractions by Solvent–Solvent Partitioning Method
2.4. Determination of Silicon (Si)
2.5. Determination of Total Phenolic Content by Folin–Ciocalteu Assay
2.6. Determination of Tyrosinase Inhibition (Skin-Whitening Activity)
2.7. Determination of Collagenase Inhibition (Anti-Wrinkle Activity)
2.8. Determination of 5α-Reductase Inhibition (Anti-Hair Loss Activity)
2.8.1. Preparation of Rat Microsomal Suspension
2.8.2. Inhibitory Assay of 5α-Reductase Activities
2.9. In Vitro Cytotoxicity Test Using MTT Assay
2.10. Stability Study
2.11. Statistical Analysis
3. Results
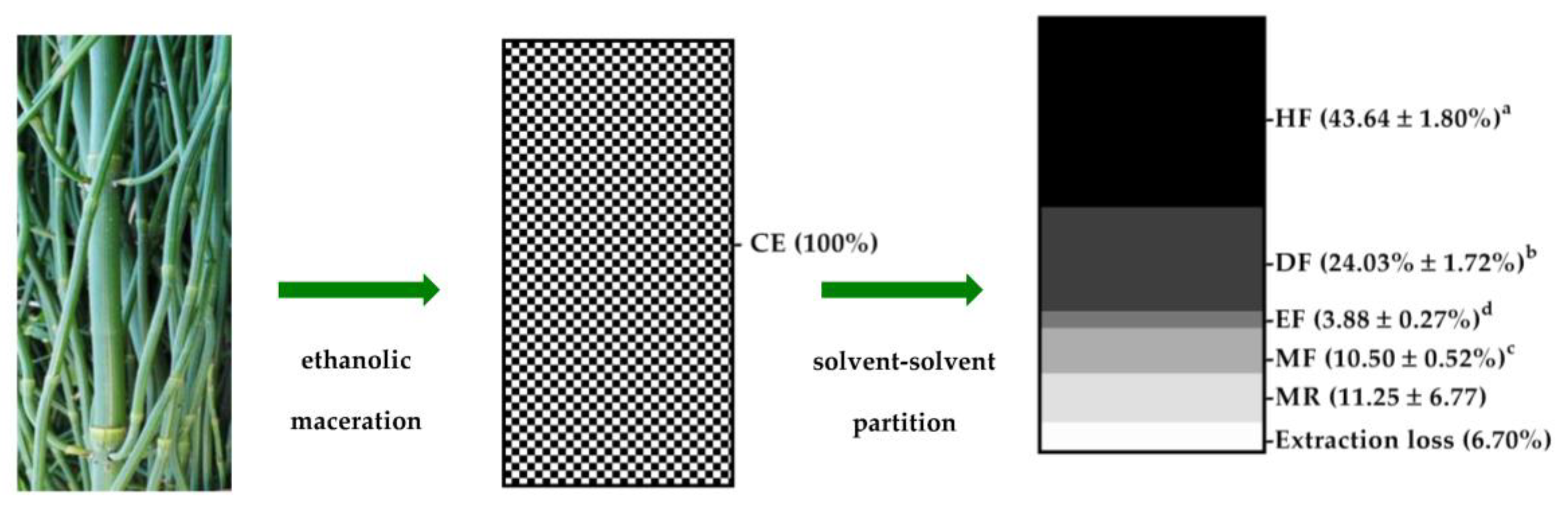
3.1. E. debile Crude Extract and Semi-Purified Fractions
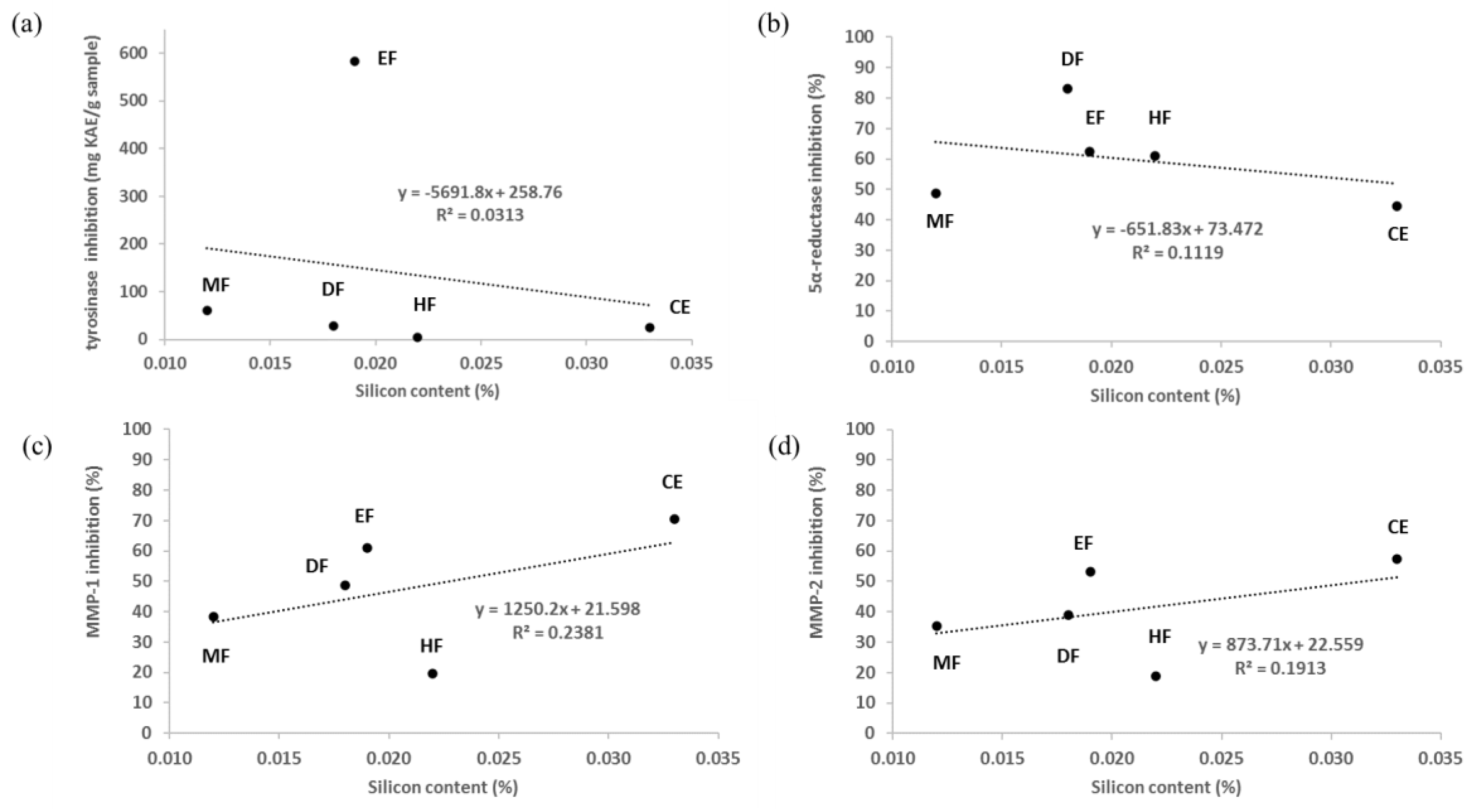
3.2. Determination of Silicon Contents
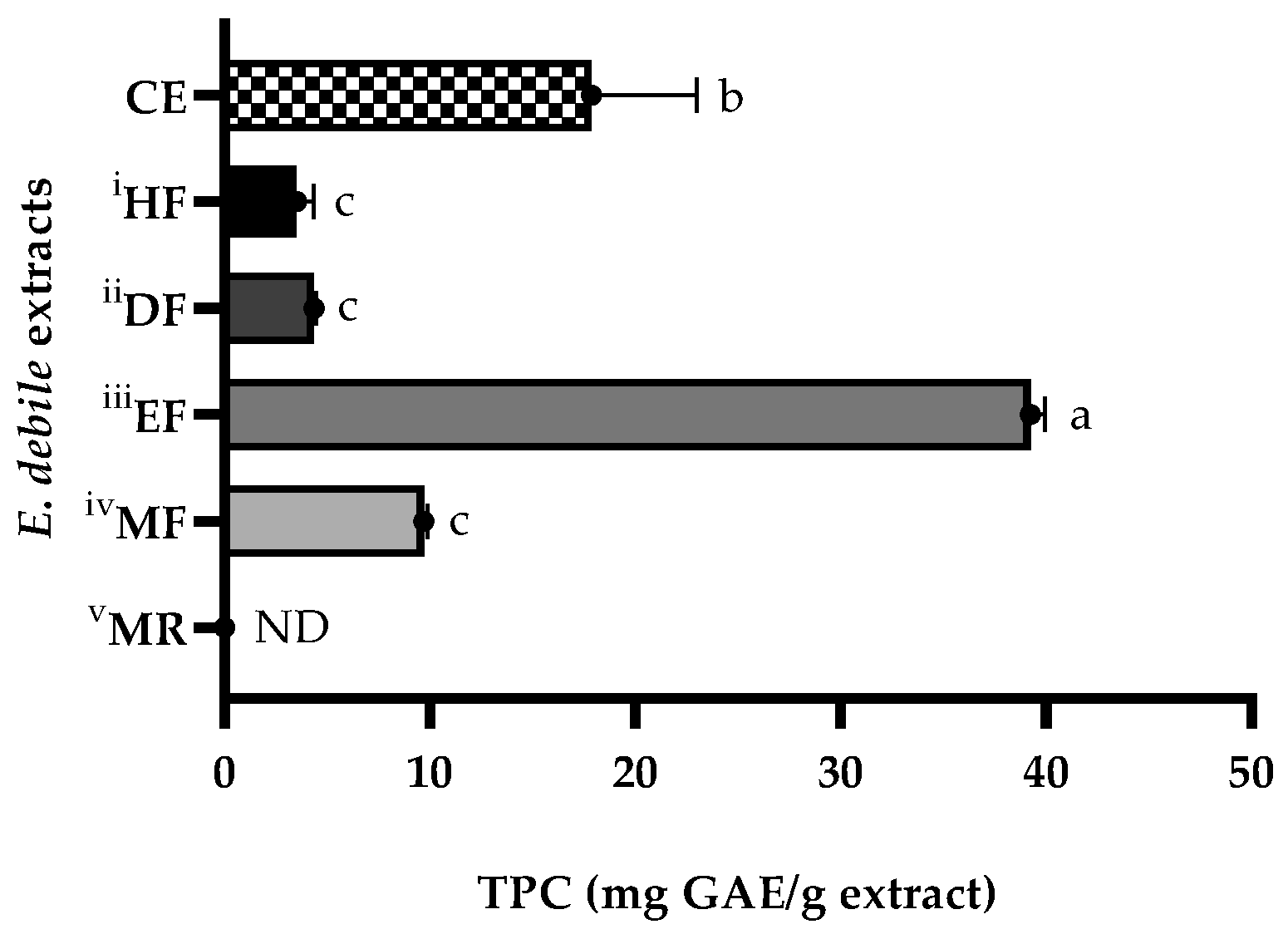
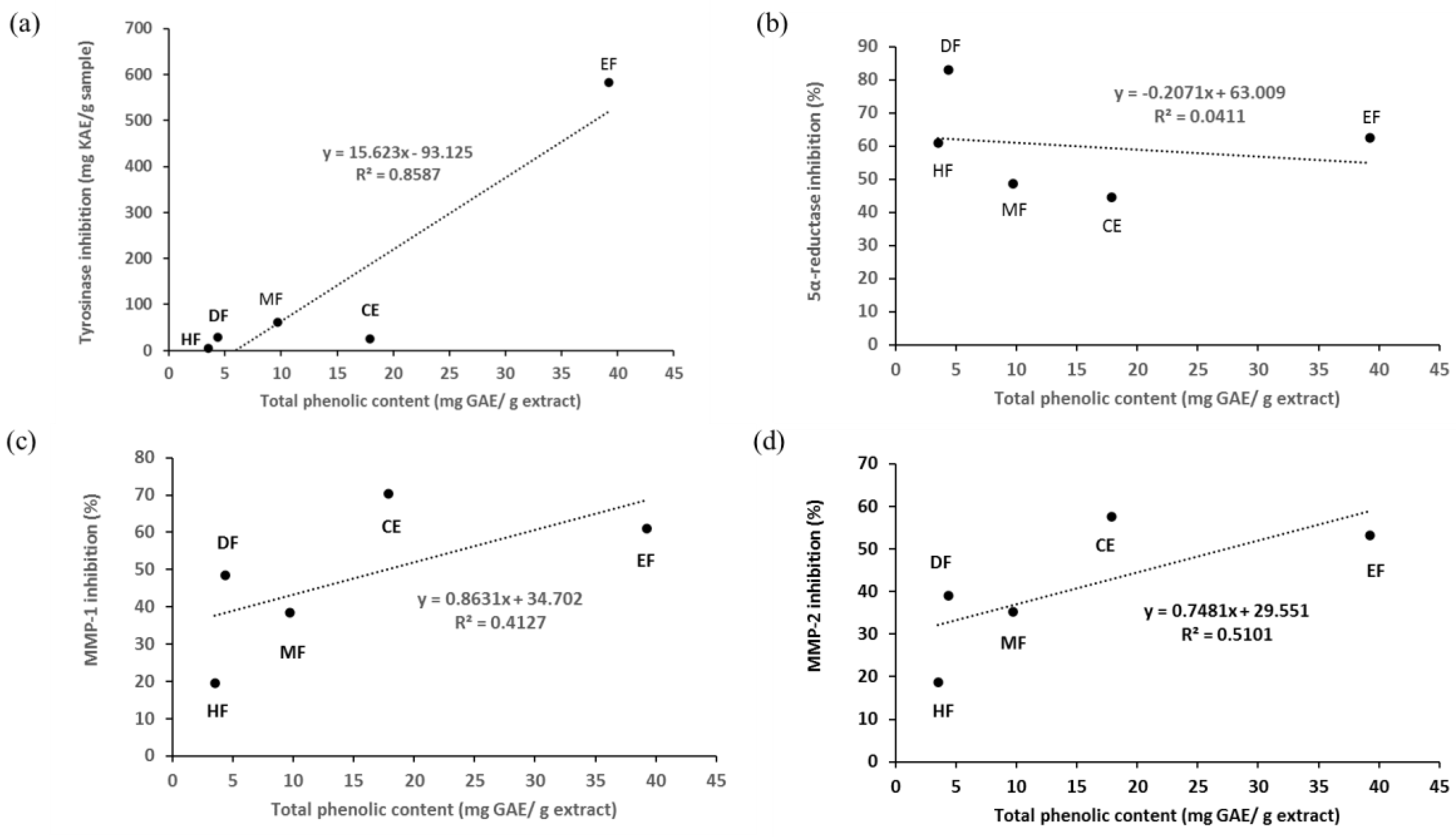
3.3. Determination of Total Phenolic Contents
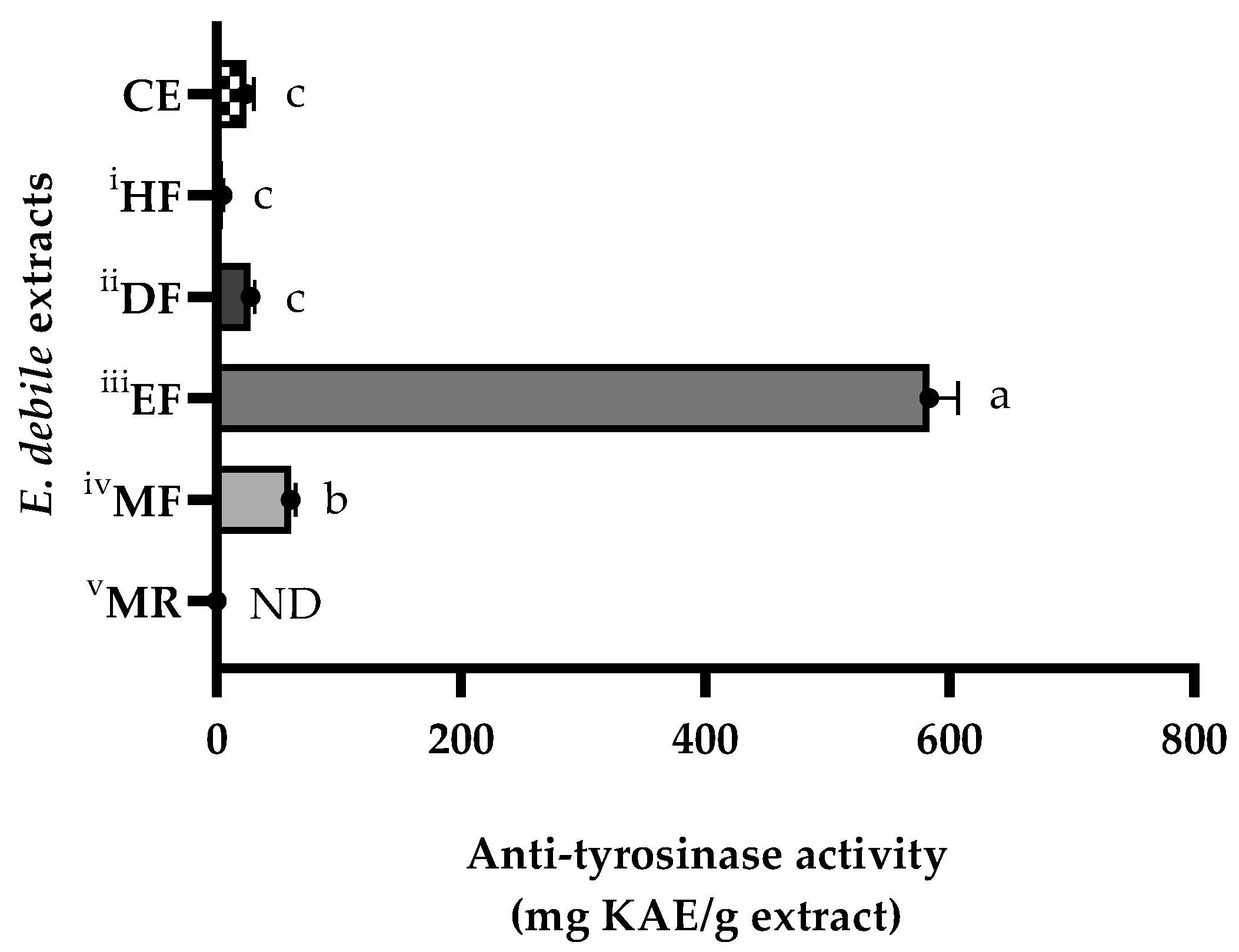
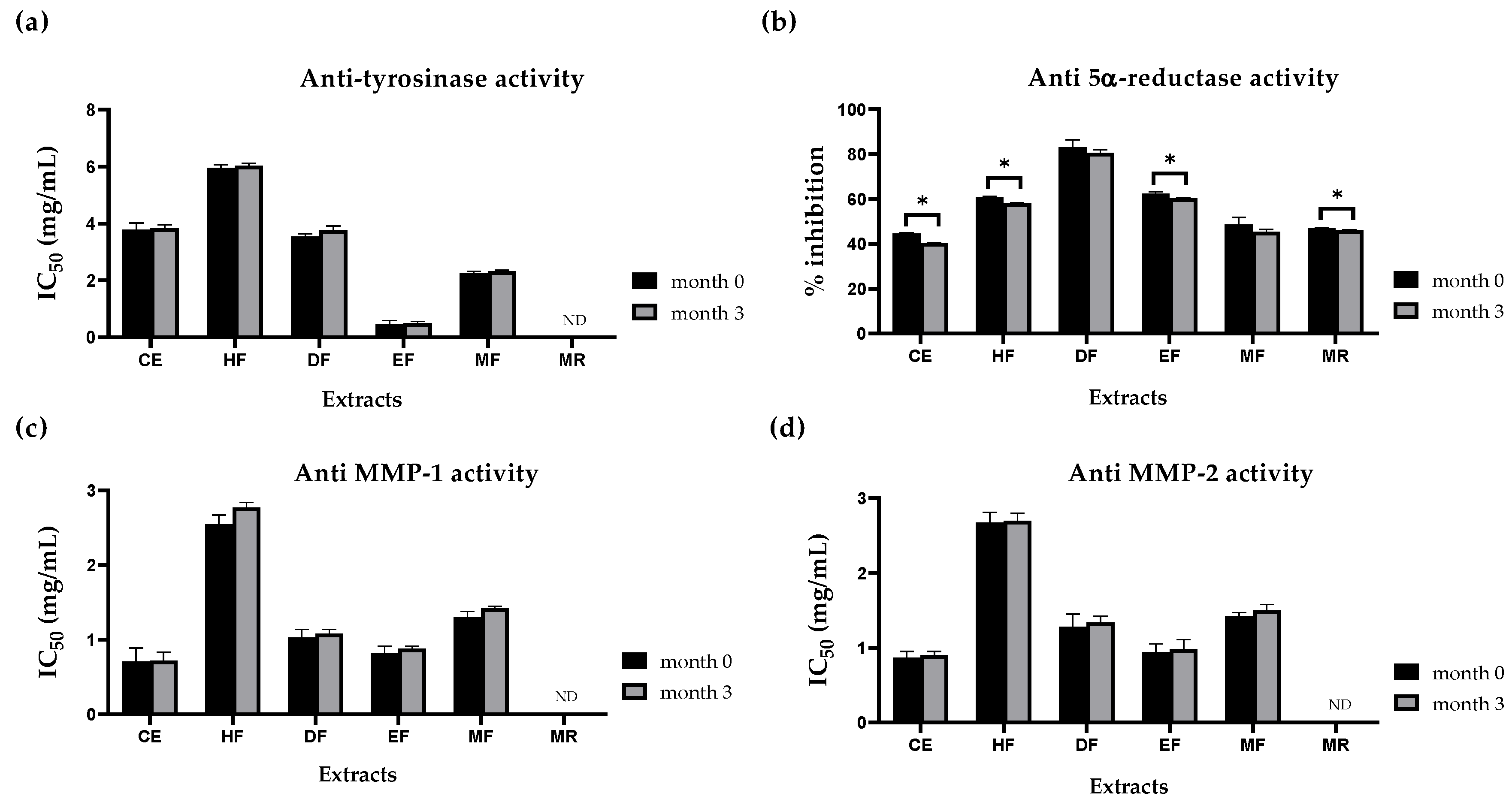
3.4. Skin-Whitening Activities of E. debile Extracts via the Inhibition of Tyrosinase Enzyme
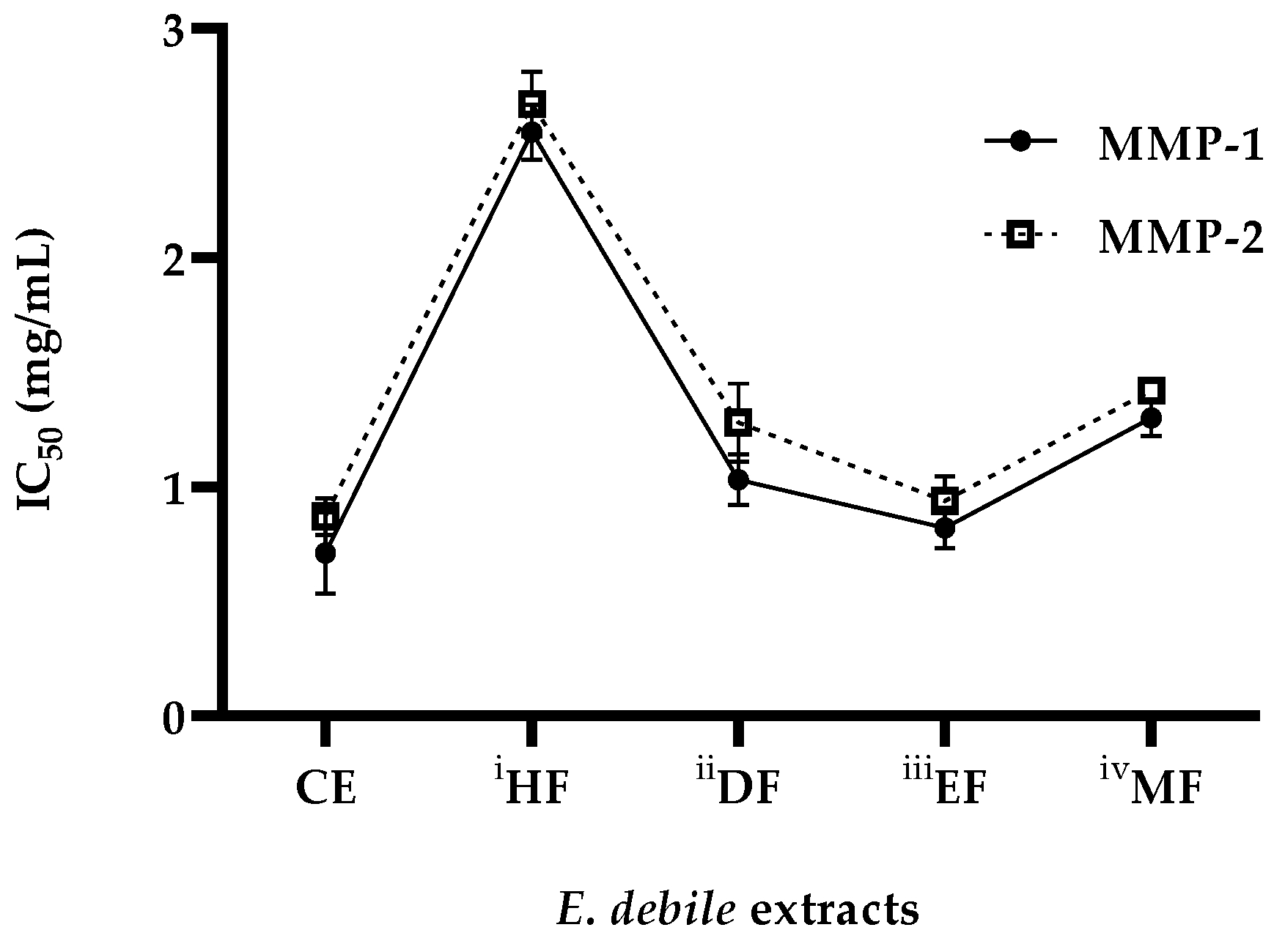
3.5. Anti-Wrinkle Activities of E. debile Extracts via the Inhibition of Collagenase Enzymes
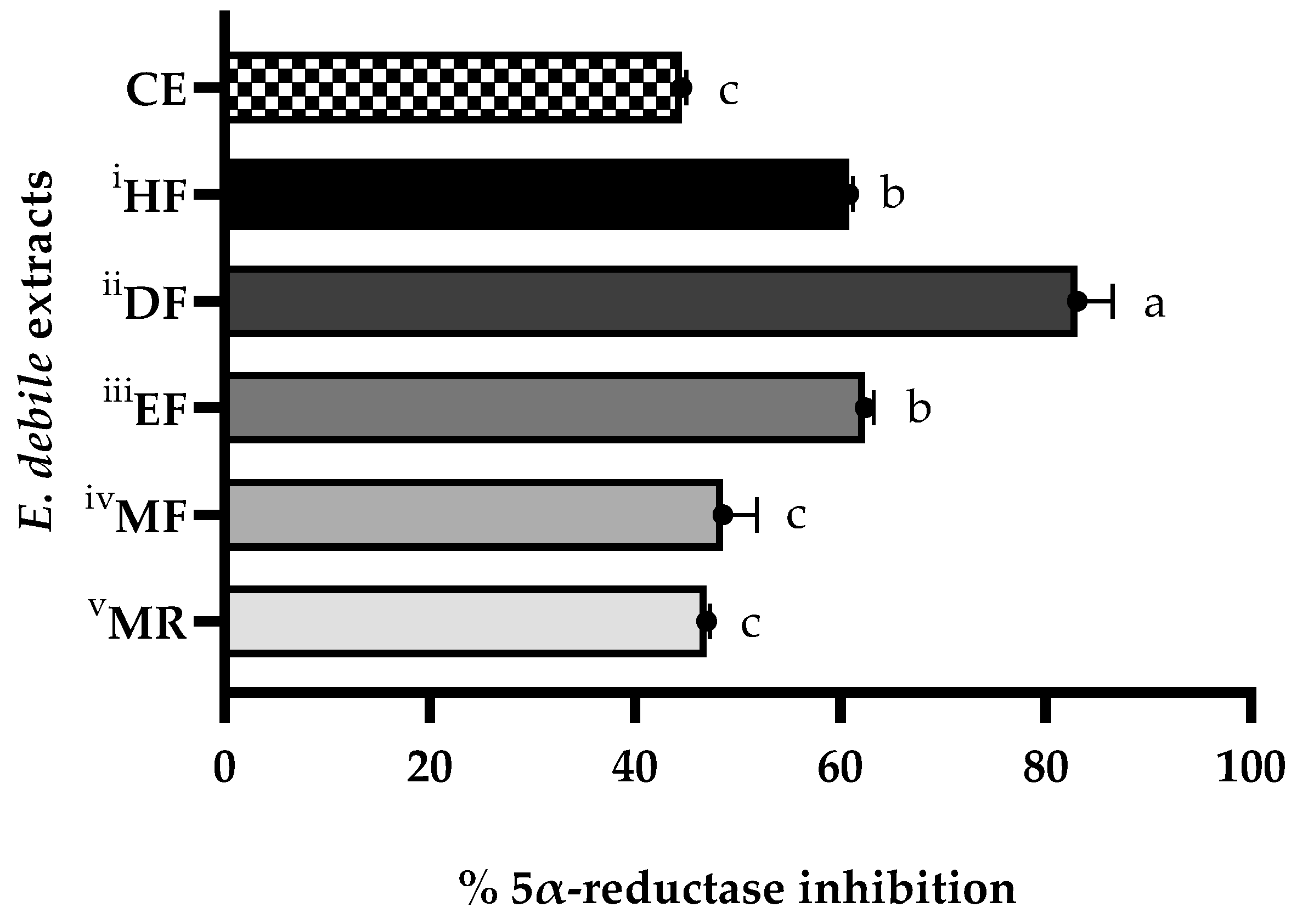
3.6. Anti-Hair Loss Activities of E. debile Extracts via the Inhibition of 5α-Reductase Enzyme
3.7. Evaluation of Anti-Tyrosinase, Anti-Collagenase, and Anti 5α-Reductase Activities of E. debile Extracts during Storage
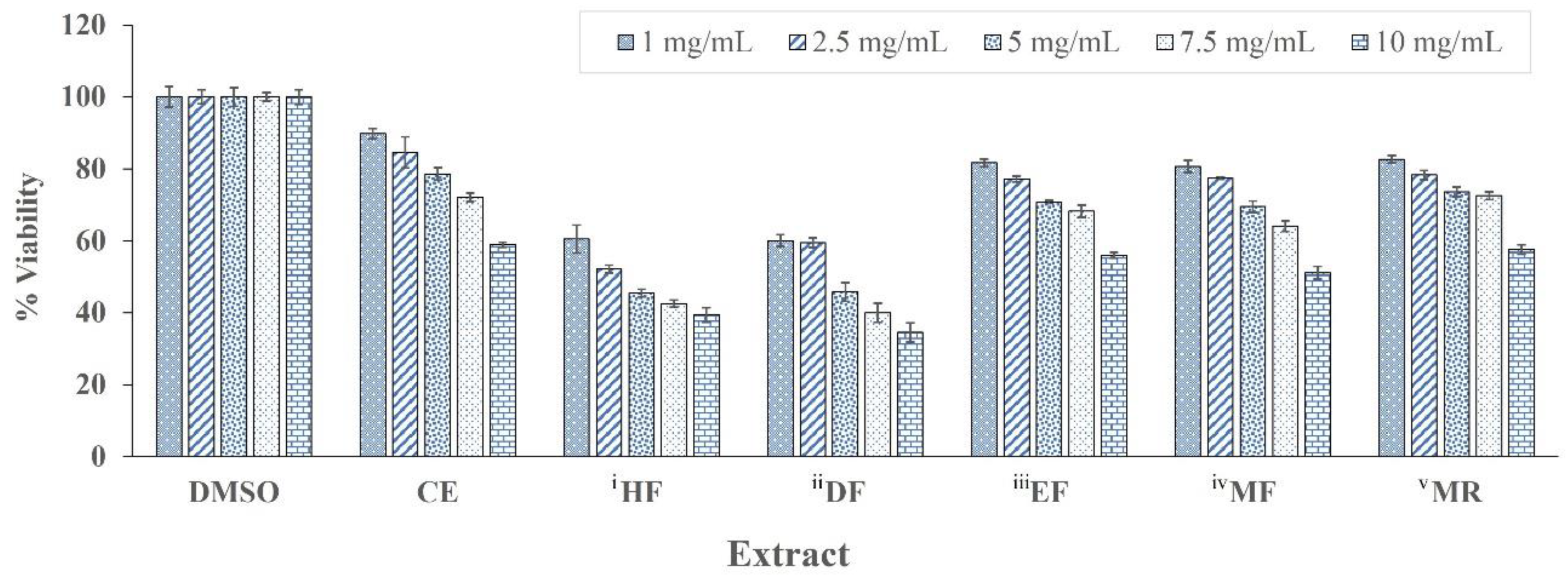
3.8. In Vitro Cytotoxicity Test Using MTT Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Akinseye, O.; Tovar-Garza, A.; Pandya, A. The effect of melasma on self-esteem: A pilot study. Int. J. Women’s Dermatol. 2018, 4, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Stough, D.; Stenn, K.; Haber, R.; Parsley, W.M.; Vogel, J.E.; Whiting, D.A.; Washenik, K. Psychological effect, pathophysiology, and management of androgenetic alopecia in men. Mayo Clin. Proc. 2005, 80, 1316–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assaf, H.; Adly, M.; Hussein, M. Aging and Intrinsic Aging, Pathogenesis and Manifestations. In Textbook of Aging Skin; Farage, M.A., Miller, K.W., Maibach, H.I., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 129–138. [Google Scholar]
- Demeule, M.; Brossard, M.; Pagé, M.; Gingras, D.; Béliveau, R. Matrix metalloproteinase inhibition by green tea catechins. Biochim. Biophys. Acta Prot. Struct. Mol. Enzym. 2000, 1478, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.U.; Kothari, S.; Chavan, B.; Spencer, J.D. Regulation of melanogenesis–controversies and new concepts. Exp. Dermatol. 2008, 17, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Sim, G.S.; Lee, B.C.; Cho, H.S.; Lee, J.W.; Kim, J.H.; Lee, D.H.; Kim, J.H.; Pyo, H.B.; Moon, D.C.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef]
- Winder, A.; Kobayashi, T.; Tsukamoto, K.; Urabe, K.; Aroca, P.; Kameyama, K.; Hearing, V.J. The tyrosinase gene family--interactions of melanogenic proteins to regulate melanogenesis. Cell Mol. Biol. Res. 1994, 40, 613–626. [Google Scholar]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Nicolaidou, E.; Katsambas, A.D. Pigmentation disorders: Hyperpigmentation and hypopigmentation. Clin. Dermatol. 2014, 32, 66–72. [Google Scholar] [CrossRef]
- Rigopoulos, D.; Gregoriou, S.; Katsambas, A. Hyperpigmentation and melasma. J. Cosmet. Dermatol. 2007, 6, 195–202. [Google Scholar] [CrossRef]
- Brett, D.A. review of collagen and collagen-based wound dressings. Wounds 2008, 20, 347–356. [Google Scholar] [PubMed]
- Vu, T.H.; Werb, Z. Matrix metalloproteinases: Effectors of development and normal physiology. Genes Dev. 2000, 14, 2123–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takema, Y.; Hattori, M.; Aizawa, K. The relationship between quantitative changes in collagen and formation of wrinkles on hairless mouse skin after chronic UV irradiation. J. Dermatol. Sci. 1996, 12, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J. Male pattern hair loss in man: Types and incidence. Ann. N. Y. Acad. Sci. 1951, 53, 708–728. [Google Scholar] [CrossRef] [PubMed]
- Severi, G.; Sinclair, R.; Hopper, J.L.; English, D.R.; McCredie, M.R.E.; Boyle, P.; Giles, G.G. Androgenetic alopecia in men aged 40–69 years: Prevalence and risk factors. Br. J. Dermatol. 2003, 149, 1207–1213. [Google Scholar] [CrossRef]
- Kaufman, K.D. Androgens and alopecia. Mol. Cell Endocrinol. 2002, 198, 89–95. [Google Scholar] [CrossRef]
- Demark-Wahnefried, W.; Halabi, S.; Paulson, D.F. Serum androgens: Associations with prostate cancer risk and hair patterning. J. Androl. 1997, 18, 495–500. [Google Scholar] [CrossRef]
- Signorelloa, L.B.; Wuu, J.; Hsieh, C.C.; Tzonou, A.; Trichopoulos, D.; Mantzoros, C.S. Hormones and hair patterning in men: A role for insulin-like growth factor 1? J. Am. Acad. Dermatol. 1999, 40, 200–203. [Google Scholar] [CrossRef]
- Olumide, Y.M.; Akinkugbe, A.O.; Altraide, D.; Mohammed, T.; Ahamefule, N.; Ayanlowo, S.; Onyekonwu, C.; Essen, N. Complications of chronic use of skin lightening cosmetics. Int. J. Dermatol. 2008, 47, 344–353. [Google Scholar] [CrossRef]
- Fischer, T.; Perosino, E.; Poli, F.; Viera, M.S.; Dreno, B. Chemical peels in aesthetic dermatology: An update 2009. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 281–292. [Google Scholar] [CrossRef]
- Rendon, M.I.; Berson, D.S.; Cohen, J.L.; Roberts, W.E.; Starker, I.; Wang, B. Evidence and considerations in the application of chemical peels in skin disorders and aesthetic resurfacing. J. Clin. Aesthet. Dermatol. 2010, 3, 32–43. [Google Scholar] [PubMed]
- Carneiro, D.M.; Jardim, T.V.; Araújo, Y.C.L.; Arantes, A.C.; Sousa, A.C.; Barroso, W.K.S.; Sousa, A.L.L.; Cunha, L.C.; Cirilo, H.N.C.; Bara, M.T.F.; et al. Equisetum arvense: New evidences supports medical use in daily clinic. Pharmacogn. Rev. 2019, 13, 50–58. [Google Scholar] [CrossRef]
- Schaffner, J.H. Geographic distribution of the species of Equisetum in relation to their phylogeny. Am. Fern. J. 1930, 20, 89–106. [Google Scholar] [CrossRef]
- Badole, S.; Kotwal, S. Equisetum arvense: Ethanopharmacological and Phytochemical review with reference to osteoporosis. Int. J. Pharm. Sci. Health Care 2014, 1, 131–141. [Google Scholar]
- Law, C.; Exley, C. New insight into silica deposition in horsetail (Equisetum arvense). BMC Plant Biol. 2011, 11, 112. [Google Scholar] [CrossRef] [Green Version]
- Pekmezci, E.; Dündar, C.; Türkoğlu, M. A proprietary herbal extract against hair loss in androgenetic alopecia and telogen effluvium: A placebo-controlled, single-blind, clinical-instrumental study. Acta Derm. Alp Pannon Adriat 2018, 27, 51–57. [Google Scholar] [CrossRef]
- Alexandru, V.; Gaspar, A.; Savin, S.; Toma, A.; Tatia, R.; Gille, E. Phenolic content, antioxidant activity and effect on collagen synthesis of a traditional wound healing polyherbal formula. Stud. Univ. Vasile Goldis Arad Ser. Stiint. Vietii 2015, 25, 41–46. [Google Scholar]
- Al-Snafi, A.E. The pharmacology of Equisetum arvense-A review. IOSR J. Pharm. 2017, 7, 31–42. [Google Scholar] [CrossRef]
- Carneiro, D.M.; Freire, R.C.; Deus Honório, T.C.; Zoghaib, I.; Cardoso, F.F.S.E.S.; Tresvenzol, L.M.F.; Paula, J.R.; Sousa, A.L.L.; Jardim, P.C.B.V.; Cunha, L.C. Randomized, double-blind clinical trial to assess the acute diuretic effect of Equisetum arvense (field horsetail) in healthy volunteers. Evid. Based Complement Altern. Med. 2014, 2014, 760683. [Google Scholar] [CrossRef] [Green Version]
- Council of Europe. European Pharmacopoeia, 8th ed.; European Directorate for the Quality of Medicines & Healthcare, Council of Europe: Strasbourg, France, 2015; pp. 2013–2014. [Google Scholar]
- Chaiyana, W.; Punyoyai, C.; Somwongin, S.; Leelapornpisid, P.; Ingkaninan, K.; Waranuch, N.; Srivilai, J.; Thitipramote, N.; Wisuitiprot, W.; Schuster, R.; et al. Inhibition of 5α-reductase, IL-6 secretion, and oxidation process of Equisetum debile Roxb. ex vaucher extract as functional food and nutraceuticals ingredients. Nutrients 2017, 9, 1105. [Google Scholar] [CrossRef]
- Kanchanapoom, T.; Otsuka, H.; Ruchirawat, S. Megastigmane glucosides from Equisetum debile and E. diffusum. Chem. Pharm. Bull. 2007, 55, 1277–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, B.; Raihan, S.M.A.; Sultana, N.; Rahman, R.; Islam, M.E.; Ahmed, S.; Akter, S. Cytotoxic, antibacterial and free radical scavenging activity studies of the solvent extracts of aerial stems of Equisetum debile Roxb. Int. J. Chem. Sci. 2012, 10, 19–26. [Google Scholar]
- Bye, R.; Thingstad, S.F.; Paulsen, B.S. Horsetail (Equisetum spp.) as a Source of Silicon Supplement in Human Nutrition—A Myth? J. Herbs Spices Med. Plants 2010, 16, 119–125. [Google Scholar] [CrossRef]
- Wickett, R.R.; Kossmann, E.; Barel, A.; Demeester, N.; Clarys, P.; Vanden Berghe, D.; Calomme, M. Effect of oral intake of choline-stabilized orthosilicic acid on hair tensile strength and morphology in women with fine hair. Arch. Dermatol. Res. 2007, 299, 499–505. [Google Scholar] [CrossRef]
- Barel, A.; Calomme, M.; Timchenko, A.; De Paepe, K.; Demeester, N.; Rogiers, V.; Clarys, P.; Vanden Berghe, D. Effect of oral intake of choline-stabilized orthosilicic acid on skin, nails and hair in women with photodamaged skin. Arch. Dermatol. Res. 2005, 297, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Organización Mundial de la Salud and World Health Organization. WHO Guidelines on Good Agricultural and Collection Practices (GACP) for Medicinal Plants; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- European Medicines Agency. Guideline on Good Agricultural and Collection Practice (GACP) for Starting Materials of Herbal Origin; European Medicines Agency Evaluation of Medicines for Human Use: London, UK, 2006; pp. 1–11.
- Chatha, S.A.S.; Anwar, F.; Manzoor, M. Evaluation of the antioxidant activity of rice bran extracts using different antioxidant assays. Grasas Y Aceites 2006, 57, 328–335. [Google Scholar]
- Siddhuraju, P.; Becker, K. Antioxidant properties of various solvent extracts of total phenolic constituents from three different agroclimatic origins of drumstick tree (Moringa oleifera Lam.) leaves. J. Agric Food Chem. 2003, 51, 2144–2155. [Google Scholar] [CrossRef] [PubMed]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Emran, T.B.; Rahman, A.; Uddin, M.N.; Rahman, M.; Uddin, Z.; Dash, R.; Layzu, C. Effects of organic extracts and their different fractions of five Bangladeshi plants on in vitro thrombolysis. BMC Complement Altern Med. 2015, 15, 128. [Google Scholar] [CrossRef] [Green Version]
- Thammarat, P.; Sirilun, S.; Phongpradist, R.; Raiwa, A.; Pandith, H.; Jiaranaikulwanitch, J. Validated HPTLC and antioxidant activities for quality control of catechin in a fermented tea (Camellia sinensis var. assamica). Food Sci. Nutr. 2021, 9, 3228–3239. [Google Scholar] [CrossRef]
- Fox, C. Advances in cosmetic science and technology. IV: Skin care and treatment. Cosmet. Toilet. 1995, 110, 63–93. [Google Scholar]
- Chattuwatthana, T.; Okello, E. Anti-collagenase, anti-elastase and antioxidant activities of Pueraria candollei var. mirifica root extract and Coccinia grandis fruit juice extract: An in vitro study. Eur. J. Med. Plants 2015, 5, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Bigg, H.F.; Clark, I.M.; Cawston, T.E. Fragments of human fibroblast collagenase: Interaction with metalloproteinase inhibitors and substrates. Biochim. Biophys. Acta Prot. Struct. Mol. Enzym. 1994, 1208, 157–165. [Google Scholar] [CrossRef]
- Eun Lee, K.; Bharadwaj, S.; Yadava, U.; Gu Kang, S. Evaluation of caffeine as inhibitor against collagenase, elastase and tyrosinase using in silico and in vitro approach. J. Enzym. Inhib. Med. Chem. 2019, 34, 927–936. [Google Scholar] [CrossRef] [Green Version]
- EnzChek™ Gelatinase/Collagenase Assay Kit. Available online: https://assets.fishersci.com/TFS-Assets/LSG/manuals/mp12052.pdf (accessed on 5 January 2023).
- Kumar, T.; Chaiyasut, C.; Rungseevijitprapa, W.; Suttajit, M. Screening of steroid 5α-reductase inhibitory activity and total phenolic content of Thai plants. J. Med. Plant Res. 2011, 5, 1265–1271. [Google Scholar]
- Khat-udomkiri, N.; Toejing, P.; Sirilun, S.; Chaiyasut, C.; Lailerd, N. Antihyperglycemic effect of rice husk derived xylooligosaccharides in high-fat diet and low-dose streptozotocin-induced type 2 diabetic rat model. Food Sci. Nutr. 2020, 8, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein determination by a modified Folin phenol method. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.R.; Castro, V.C.; Graças Figueiredo Vilela, P.; Camargo, S.E.A.; Carvalho, C.A.T.; Jorge, A.O.C.; Oliveira, L.D. Cytotoxicity of Brazilian plant extracts against oral microorganisms of interest to dentistry. BMC Complement Altern Med. 2013, 13, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazekawa, M.; Nishinakagawa, T.; Kawakubo-Yasukochi, T.; Nakashima, M. Evaluation of IC50 levels immediately after treatment with anticancer reagents using a real time cell monitoring device. Exp. Ther. Med. 2019, 18, 3197–3205. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, B.; Raihan, S.; Sultana, N.; Islam, M.E. Isolation of two steroids and two flavonoids having antioxidant, antibacterial and cytotoxic properties from aerial stems of Equisetum debile Roxb. Nat. Univ. J. Sci. 2014, 1, 31–42. [Google Scholar]
- Inanaga, S.; Okasaka, A.; Tanaka, S. Does silicon exist in association with organic compounds in rice plant? Soil. Sci. Plant Nutr. 1995, 41, 111–117. [Google Scholar] [CrossRef]
- Perry, C.C.; Lu, Y. Preparation of silicas from silicon complexes: Role of cellulose in polymerisation and aggregation control. J. Chem. Soc. Faraday Trans. 1992, 88, 2915–2921. [Google Scholar] [CrossRef]
- Li, P.H.; Chiu, Y.P.; Shih, C.C.; Wen, Z.H.; Ibeto, L.K.; Huang, S.H.; Chiu, C.C.; Ma, D.L.; Leung, C.H.; Chang, Y.N.; et al. Biofunctional activities of Equisetum ramosissimum extract: Protective effects against oxidation, melanoma, and melanogenesis. Oxid. Med. Cell Longev. 2016, 2016, 2853543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.H.; Ryu, Y.B.; Curtis-Long, M.J.; Ryu, H.W.; Baek, Y.S.; Kang, J.E.; Lee, W.S.; Park, K.H. Tyrosinase inhibitory polyphenols from roots of Morus lhou. J. Agric. Food Chem. 2009, 57, 1195–1203. [Google Scholar] [CrossRef]
- Malešev, D.; Kuntić, V. Investigation of metal-flavonoid chelates and the determination of flavonoids via metal-flavonoid complexing reactions. J. Serb Chem. Soc. 2007, 72, 921–939. [Google Scholar] [CrossRef]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- El-Nashar, H.A.; El-Din, M.I.G.; Hritcu, L.; Eldahshan, O.A. Insights on the inhibitory power of flavonoids on tyrosinase activity: A survey from 2016 to 2021. Molecules 2011, 26, 7546. [Google Scholar] [CrossRef]
- Rho, H.S.; Ghimeray, A.K.; Yoo, D.S.; Ahn, S.M.; Kwon, S.S.; Lee, K.H.; Cho, D.H.; Cho, J.U. Kaempferol and kaempferol rhamnosides with depigmenting and anti-inflammatory properties. Molecules 2011, 16, 3338–3344. [Google Scholar] [CrossRef]
- Madhan, B.; Krishnamoorthy, G.; Rao, J.R.; Nair, B.U. Role of green tea polyphenols in the inhibition of collagenolytic activity by collagenase. Int. J. Biol. Macromol. 2007, 41, 16–22. [Google Scholar] [CrossRef]
- McDonald, M.; Mila, I.; Scalbert, A. Precipitation of metal ions by plant polyphenols: Optimal conditions and origin of precipitation. J. Agric Food Chem. 1996, 44, 599–606. [Google Scholar] [CrossRef]
- Lim, H.; Kim, H.P. Inhibition of mammalian collagenase, matrix metalloproteinase-1, by naturally-occurring flavonoids. Planta Med. 2007, 73, 1267–1274. [Google Scholar] [CrossRef]
- Balli, U.; Cetinkaya, B.O.; Keles, G.C.; Keles, Z.P.; Guler, S.; Sogut, M.U.; Erisgin, Z. Assessment of MMP-1, MMP-8 and TIMP-2 in experimental periodontitis treated with kaempferol. J. Periodontal. Implant. Sci. 2016, 46, 84–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.P.; Oh, K.N.; Yun, H.J.; Jeong, H.G. The flavonoids apigenin and luteolin suppress ultraviolet A-induced matrix metalloproteinase-1 expression via MAPKs and AP-1-dependent signaling in HaCaT cells. J. Dermatol. Sci. 2011, 61, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Ha, S.J.; Jung, C.H.; Kim, Y.T.; Lee, H.K.; Kim, M.O.; Lee, M.H.; Mottamal, M.; Bode, A.M.; Lee, K.W.; et al. Naringenin targets ERK 2 and suppresses UVB-induced photoaging. J. Cell Mol. Med. 2016, 20, 909–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Shimizu, K.; Kondo, R. Anti-androgenic activity of fatty acids. Chem. Biodivers 2009, 6, 503–512. [Google Scholar] [CrossRef]
- Lee, S.Y.; Choi, E.J.; Bae, D.H.; Lee, D.W.; Kim, S. Effects of 1-tetradecanol and β-sitosterol isolated from Dendropanax morbifera Lev. on skin whitening, moisturizing and preventing hair loss. J. Soc. Cosmet. Sci. Korea 2015, 41, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Cabeza, M.; Eugene, B.; Ivonne, H.; Elena, R.L.; Mauricio, S.; Eugenio, F. Effect of β-sitosterol as Inhibitor of 5α-reductase in Hamster Prostate. Proc. West Pharm. Soc. 2003, 46, 153–155. [Google Scholar]
- Hiipakka, R.A.; Zhang, H.Z.; Dai, W.; Dai, Q.; Liao, S. Structure–activity relationships for inhibition of human 5α-reductases by polyphenols. Biochem. Pharm. 2002, 63, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Parker, T.D.; Adams, D.A.; Zhou, K.; Harris, M.; Yu, L. Fatty acid composition and oxidative stability of cold-pressed edible seed oils. J. Food Sci. 2003, 68, 1240–1243. [Google Scholar] [CrossRef]
- Makhija, P.; Kathuria, H.; Sethi, G.; Grobben, B. Polymeric hydrogels for controlled release of black tea and coffee extracts for topical applications. Gels 2021, 7, 174. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instrument Parameters | Operational Conditions |
|---|---|
| ICP RF power (w) | 1300 |
| Plasma gas (L/min) | 15 |
| Auxiliary gas (L/min) | 0.2 |
| Nebulizer gas (L/min) | 0.8 |
| Viewing type | Axial |
| Read delay (seconds) | 15 |
| Replicates | 3 |
| Sample flow rate (mL/min) | 1.5 |
| Flush time (seconds) | 25 |
| Wavelength (nm) | Si 212.412 |
| Extracts | Extract Mass (mg) | Extractable Si (mg ± SD) | % Extractable Si ± SD |
|---|---|---|---|
| CE | 32.6 | 0.0109 ± 0.00011 | 0.033 ± 0.0003 a,b |
| i HF | 35.1 | 0.0078 ± 0.00009 | 0.022 ± 0.0003 b |
| ii DF | 33.6 | 0.0060 ± 0.00004 | 0.018 ± 0.0001 b |
| iii EF | 31.0 | 0.0059 ± 0.00004 | 0.019 ± 0.0002 b |
| iv MF | 32.9 | 0.0041 ± 0.00007 | 0.012 ± 0.0001 b |
| v MR | 32.6 | 0.0012 ± 0.00003 | 0.004 ± 0.0001 c,b |
| Extracts | Anti-Tyrosinase IC50 (mg/mL) | Anti-5α-Reductase FEA (mg Finasteride/g Extract) |
|---|---|---|
| CE | 3.78 ± 0.24 b | 0.83 ± 0.01 c |
| I HF | 5.96 ± 0.11 a | 1.10 ± 0.01 b |
| ii DF | 3.54 ± 0.10 b | 1.48 ± 0.06 a |
| iii EF | 0.47 ± 0.12 d | 1.13 ± 0.01 b |
| iv MF | 2.25 ± 0.08 c | 0.90 ± 0.06 c |
| v MR | ND | 0.87 ± 0.01 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thammarat, P.; Jiaranaikulwanitch, J.; Phongpradist, R.; Raiwa, A.; Pandith, H.; Sawangrat, K.; Sirilun, S. Cosmeceutical Potentials of Equisetum debile Roxb. ex Vaucher Extracts. Appl. Sci. 2023, 13, 1336. https://doi.org/10.3390/app13031336
Thammarat P, Jiaranaikulwanitch J, Phongpradist R, Raiwa A, Pandith H, Sawangrat K, Sirilun S. Cosmeceutical Potentials of Equisetum debile Roxb. ex Vaucher Extracts. Applied Sciences. 2023; 13(3):1336. https://doi.org/10.3390/app13031336
Chicago/Turabian StyleThammarat, Phanit, Jutamas Jiaranaikulwanitch, Rungsinee Phongpradist, Araya Raiwa, Hataichanok Pandith, Kasirawat Sawangrat, and Sasithorn Sirilun. 2023. "Cosmeceutical Potentials of Equisetum debile Roxb. ex Vaucher Extracts" Applied Sciences 13, no. 3: 1336. https://doi.org/10.3390/app13031336
APA StyleThammarat, P., Jiaranaikulwanitch, J., Phongpradist, R., Raiwa, A., Pandith, H., Sawangrat, K., & Sirilun, S. (2023). Cosmeceutical Potentials of Equisetum debile Roxb. ex Vaucher Extracts. Applied Sciences, 13(3), 1336. https://doi.org/10.3390/app13031336