
Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor




Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. System Preparation
2.2. Molecular Docking and Molecular Modeling Details
2.3. MD Simulations Details
2.4. MM/G(P)BSA Calculations
2.5. Analysis
3. Results
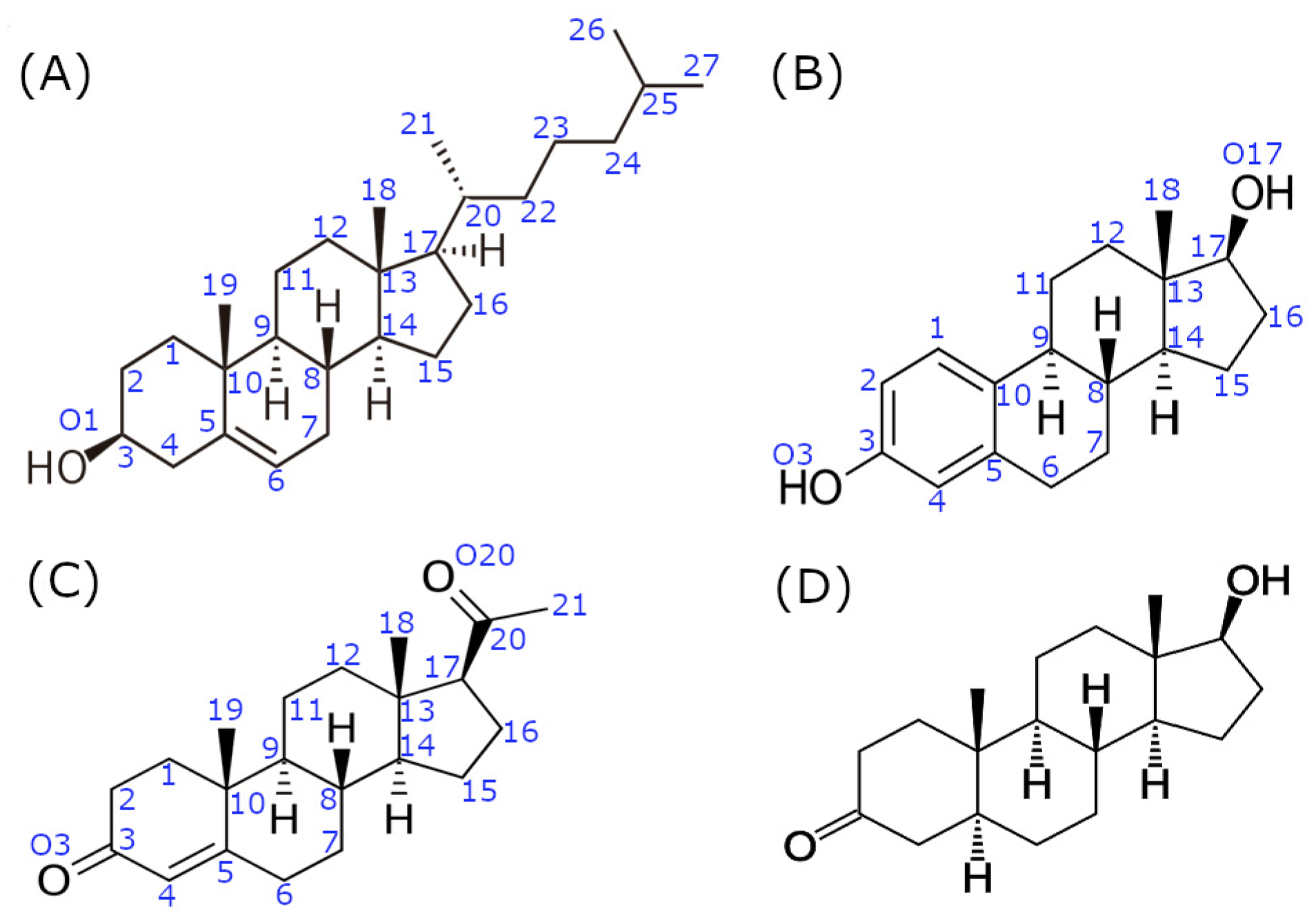
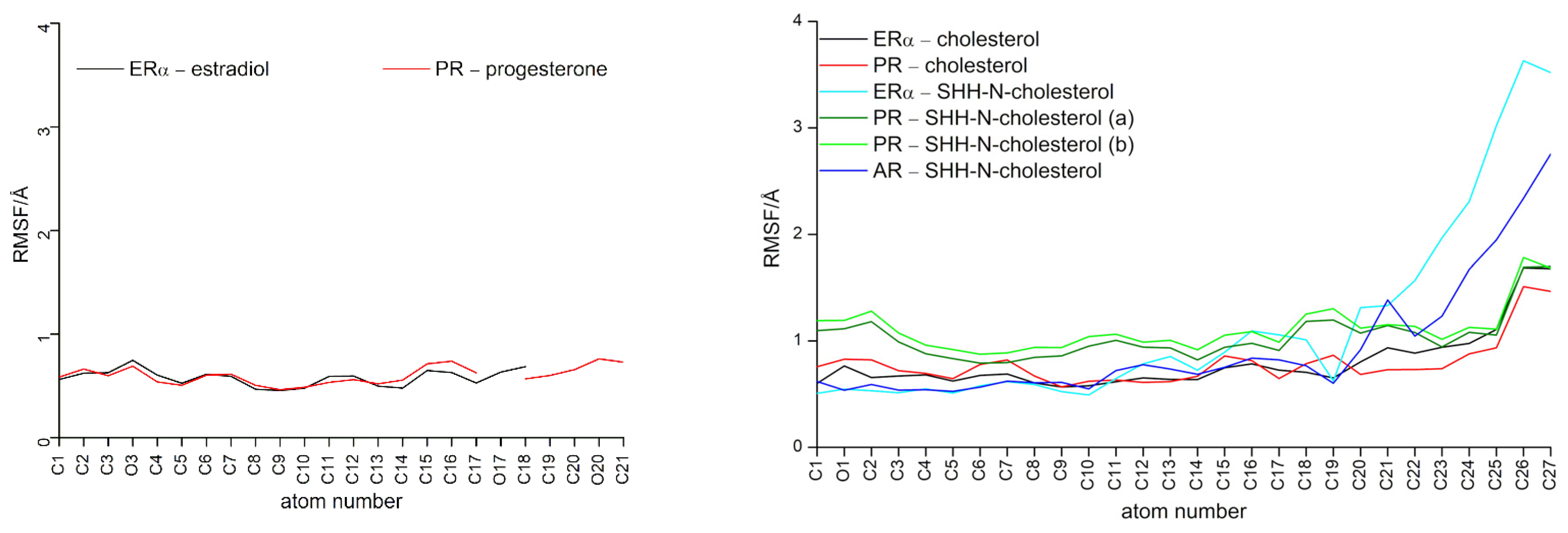
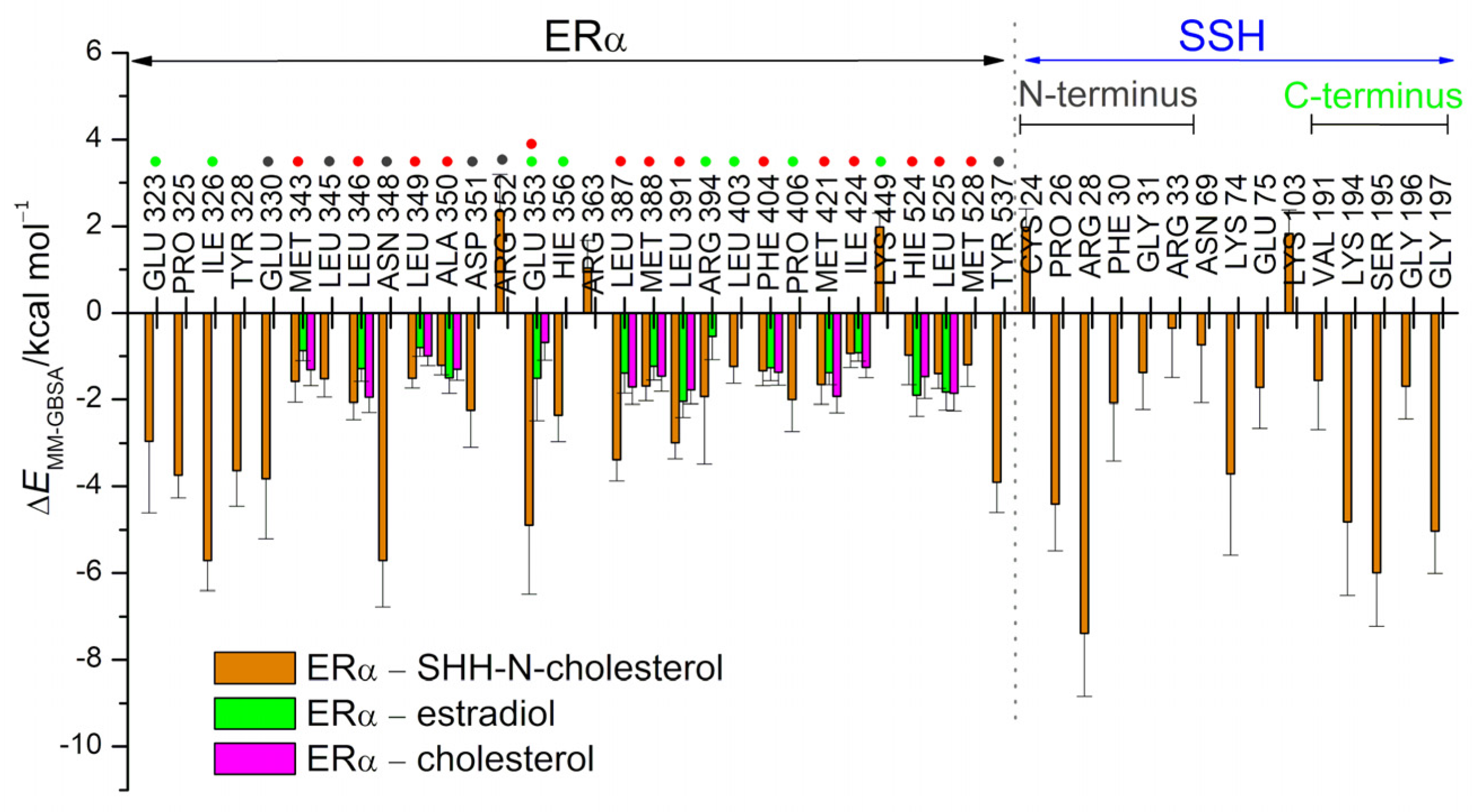
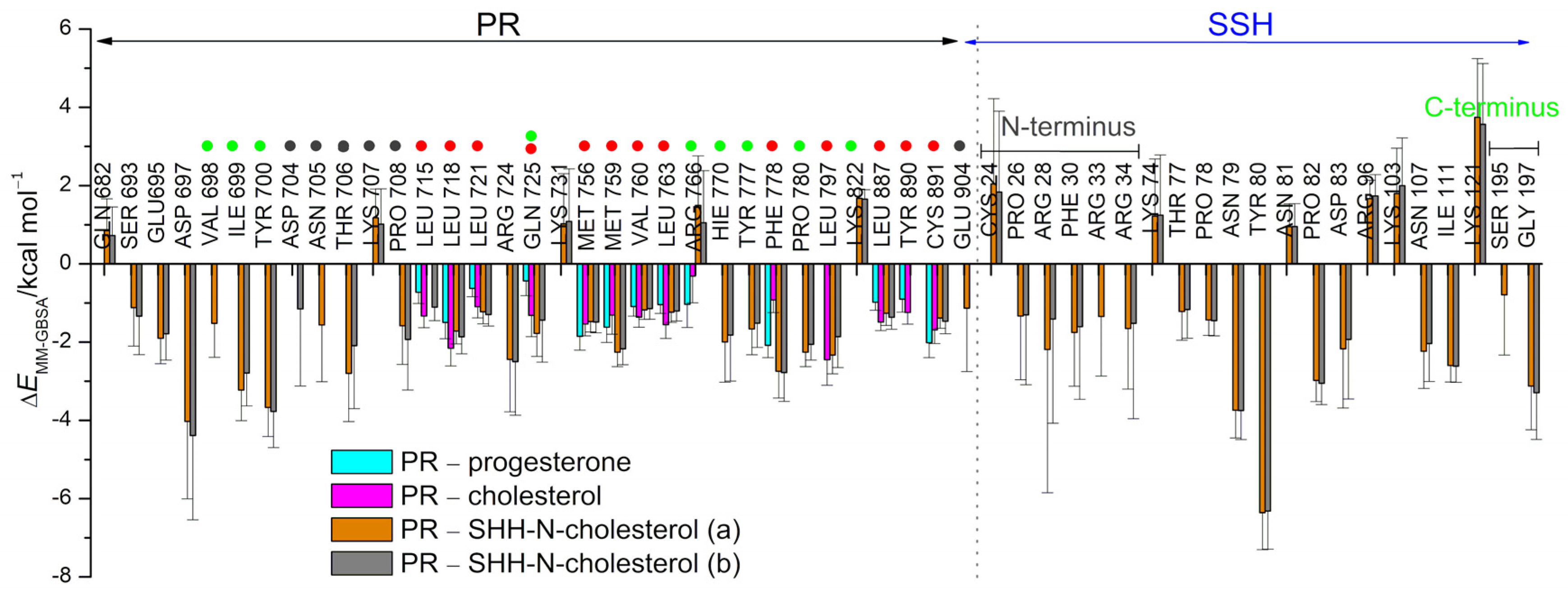
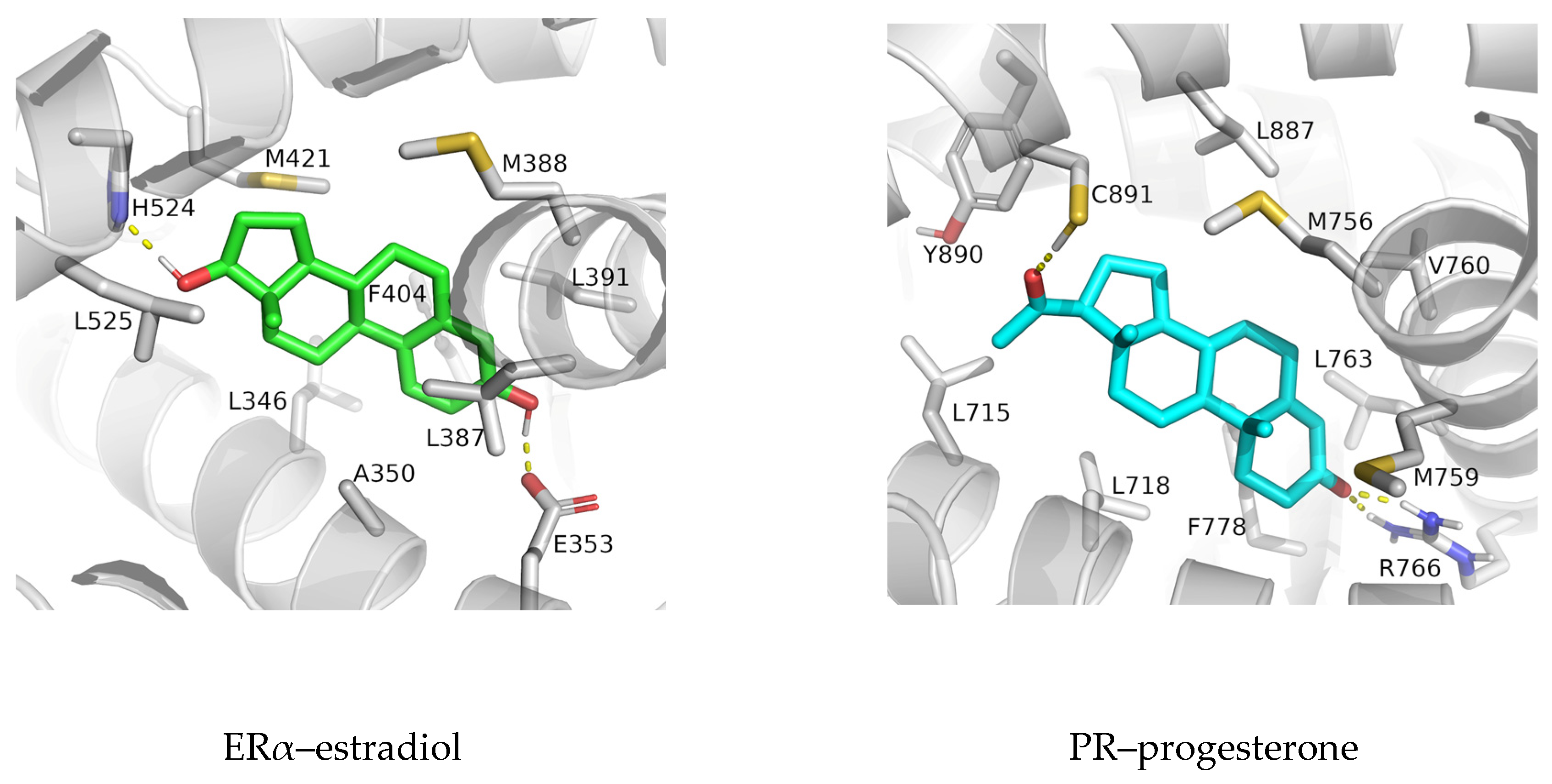
3.1. Complexes with Estradiol and Progesterone
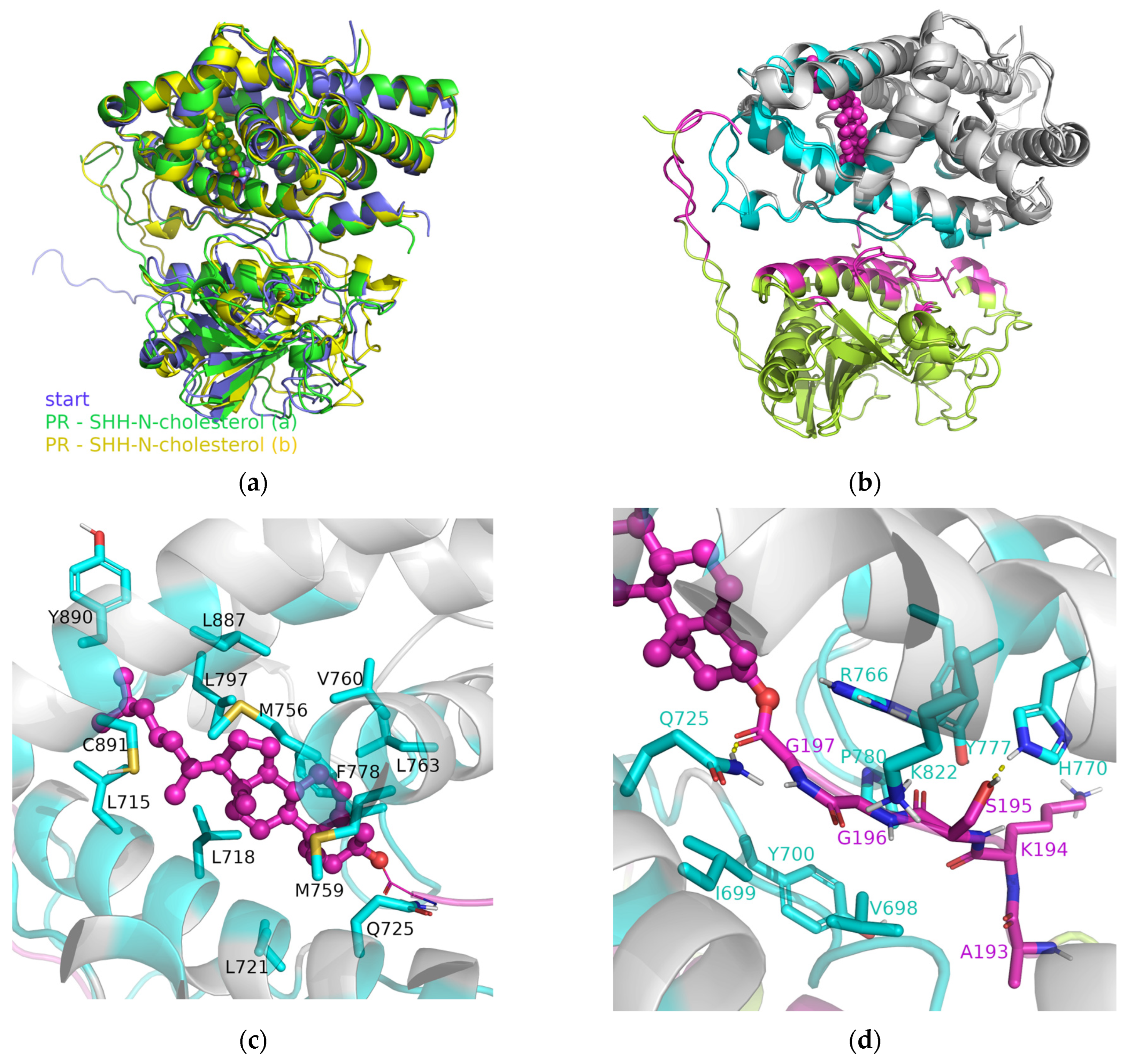
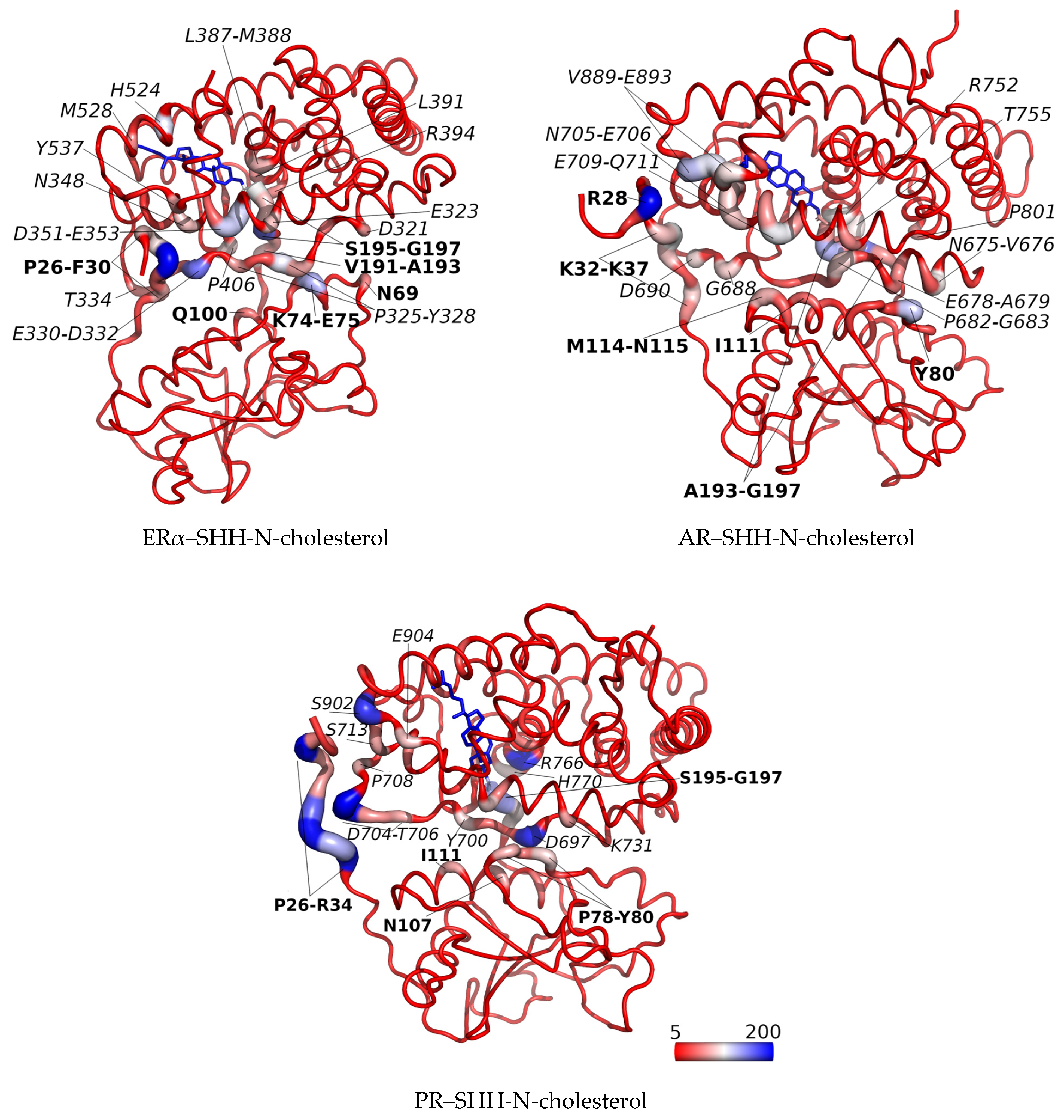
3.2. SHRs in Complex with SHH-N-Cholesterol (PP Complexes)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sigafoos, A.N.; Paradise, B.D.; Fernandez-Zapico, M.E. Hedgehog/GLI Signaling Pathway: Transduction, Regulation, and Implications for Disease. Cancers 2021, 13, 3410. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Li, S.-P.; Taipale, J. Divergence of Hedgehog Signal Transduction Mechanism between Drosophila and Mammals. Dev. Cell 2006, 10, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kurtović, M.; Piteša, N.; Bartoniček, N.; Ozretić, P.; Musani, V.; Čonkaš, J.; Petrić, T.; King, C.; Sabol, M. RNA-Seq and ChIP-Seq Identification of Unique and Overlapping Targets of GLI Transcription Factors in Melanoma Cell Lines. Cancers 2022, 14, 4540. [Google Scholar] [CrossRef] [PubMed]
- Dawber, R.J.; Hebbes, S.; Herpers, B.; Docquier, F.; van den Heuvel, M. Differential Range and Activity of Various Forms of the Hedgehog Protein. BMC Dev. Biol. 2005, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Taipale, J. Hedgehog: Functions and Mechanisms. Genes Dev. 2008, 22, 2454–2472. [Google Scholar] [CrossRef] [PubMed]
- Ciepla, P.; Magee, A.I.; Tate, E.W. Cholesterylation: A Tail of Hedgehog. Biochem. Soc. Trans. 2015, 43, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Palmitoylation of Hedgehog Proteins by Hedgehog Acyltransferase: Roles in Signalling and Disease. Open Biol. 2021, 11, 200414. [Google Scholar] [CrossRef]
- Musani, V.; Ozretić, P.; Trnski, D.; Sabol, M.; Poduje, S.; Tošić, M.; Šitum, M.; Levanat, S. Potential Hot Spot for de Novo Mutations in PTCH1 Gene in Gorlin Syndrome Patients: A Case Report of Twins from Croatia. Croat. Med. J. 2018, 59, 20–24. [Google Scholar] [CrossRef]
- Car, D.; Sabol, M.; Musani, V.; Ozretić, P.; Levanat, S. Epigenetic Regulation of the Hedgehog-Gli Signaling Pathway in Cancer. Period. Biol. 2010, 112, 419–423. [Google Scholar]
- Budimir, I.; Tomasović-Lončarić, Č.; Kralik, K.; Čonkaš, J.; Eljuga, D.; Žic, R.; Gorjanc, B.; Tucaković, H.; Caktaš, D.; Jaman, J.; et al. Higher Expressions of SHH and AR Are Associated with a Positive Receptor Status and Have Impact on Survival in a Cohort of Croatian Breast Cancer Patients. Life 2022, 12, 1559. [Google Scholar] [CrossRef]
- Levanat, S.; Sabol, M.; Musani, V.; Ozretić, P.; Trnski, D. Hedgehog Signaling Pathway as Genetic and Epigenetic Target in Ovarian Tumors. Curr. Pharm. Des. 2016, 22, 73–94. [Google Scholar] [CrossRef]
- Liao, X.; Siu, M.K.; Au, C.W.; Chan, Q.K.; Chan, H.Y.; Wong, E.S.; Ip, P.P.; Ngan, H.Y.; Cheung, A.N. Aberrant Activation of Hedgehog Signaling Pathway Contributes to Endometrial Carcinogenesis through β-Catenin. Mod. Pathol. 2009, 22, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Bushman, W. Hedgehog Signaling in Prostate Development, Regeneration and Cancer. J. Dev. Biol. 2016, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.E.; Feigelson, H.S. Hormonal Carcinogenesis. Carcinogenesis 2000, 21, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Rochefort, H.; Chalbos, D. The Role of Sex Steroid Receptors on Lipogenesis in Breast and Prostate Carcinogenesis: A Viewpoint. Horm. Cancer 2010, 1, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Olefsky, J.M. Nuclear Receptor Minireview Series. J. Biol. Chem. 2001, 276, 36863–36864. [Google Scholar] [CrossRef] [PubMed]
- Sever, R.; Glass, C.K. Signaling by Nuclear Receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef] [PubMed]
- Sabol, M.; Trnski, D.; Uzarevic, Z.; Ozretic, P.; Musani, V.; Rafaj, M.; Cindric, M.; Levanat, S. Combination of Cyclopamine and Tamoxifen Promotes Survival and Migration of MCF-7 Breast Cancer Cells—Interaction of Hedgehog-Gli and Estrogen Receptor Signaling Pathways. PLoS ONE 2014, 9, e114510. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Trnski, D.; Sabol, M.; Tomić, S.; Štefanac, I.; Mrčela, M.; Musani, V.; Rinčić, N.; Kurtović, M.; Petrić, T.; Levanat, S.; et al. SHH-N Non-Canonically Sustains Androgen Receptor Activity in Androgen-Independent Prostate Cancer Cells. Sci. Rep. 2021, 11, 14880. [Google Scholar] [CrossRef]
- Sartorius, C.A.; Melville, M.Y.; Hovland, A.R.; Tung, L.; Takimoto, G.S.; Horwitz, K.B. A Third Transactivation Function (AF3) of Human Progesterone Receptors Located in the Unique N-Terminal Segment of the B-Isoform. Mol. Endocrinol. 1994, 8, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Hopp, T.A.; Weiss, H.L.; Hilsenbeck, S.G.; Cui, Y.; Allred, D.C.; Horwitz, K.B.; Fuqua, S.A.W. Breast Cancer Patients with Progesterone Receptor PR-A-Rich Tumors Have Poorer Disease-Free Survival Rates. Clin. Cancer Res. 2004, 10, 2751–2760. [Google Scholar] [CrossRef] [PubMed]
- Jaffrain-Rea, M.L.; Petrangeli, E.; Ortolani, F.; Fraioli, B.; Lise, A.; Esposito, V.; Spagnoli, L.G.; Tamburrano, G.; Frati, L.; Gulino, A. Cellular Receptors for Sex Steroids in Human Pituitary Adenomas. J. Endocrinol. 1996, 151, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Liu, D. Gene Signatures of Estrogen and Progesterone Receptor Pathways Predict the Prognosis of Colorectal Cancer. FEBS J. 2016, 283, 3115–3133. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, G.; De Leo, S.; Perrino, M.; Rossi, S.; Tosi, D.; Cirello, V.; Colombo, C.; Bulfamante, G.; Vicentini, L.; Fugazzola, L. Impact of Estrogen and Progesterone Receptor Expression on the Clinical and Molecular Features of Papillary Thyroid Cancer. Eur. J. Endocrinol. 2015, 173, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Elmaogod, E.A.; Khairy, D.; Mahmoud, A. Clinicopathological Role of Progesterone Hormone in IDH-Mutant Astrocytoma. J. Microsc. Ultrastruct. 2023. [Google Scholar] [CrossRef]
- Deli, T.; Orosz, M.; Jakab, A. Hormone Replacement Therapy in Cancer Survivors—Review of the Literature. Pathol. Oncol. Res. 2020, 26, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lan, K.-C.; Ou, Y.-C.; Wu, C.-H.; Kang, H.-Y.; Chuang, I.-C.; Fu, H.-C. Highly Expressed Progesterone Receptor B Isoform Increases Platinum Sensitivity and Survival of Ovarian High-Grade Serous Carcinoma. Cancers 2021, 13, 5578. [Google Scholar] [CrossRef]
- Yang, S.; Thiel, K.W.; Leslie, K.K. Progesterone: The Ultimate Endometrial Tumor Suppressor. Trends Endocrinol. Metab. 2011, 22, 145–152. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Das, C.; Kumari, M.; Sen, A.; Mukhopadhyay, B.; Mukhopadhyay, B. Spectrum of Meningioma with Special Reference to Prognostic Utility of ER, PR and Ki67 Expression. J. Lab. Physicians 2017, 9, 308–313. [Google Scholar] [CrossRef]
- Brzozowski, A.M.; Pike, A.C.W.; Dauter, Z.; Hubbard, R.E.; Bonn, T.; Engström, O.; Öhman, L.; Greene, G.L.; Gustafsson, J.-Å.; Carlquist, M. Molecular Basis of Agonism and Antagonism in the Oestrogen Receptor. Nature 1997, 389, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.P.; Sigler, P.B. Atomic Structure of Progesterone Complexed with Its Receptor. Nature 1998, 393, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, A.F.; Kinnebrew, M.; Kowatsch, C.; Ansell, T.B.; El Omari, K.; Bishop, B.; Pardon, E.; Schwab, R.A.; Malinauskas, T.; Qian, M.; et al. The Morphogen Sonic Hedgehog Inhibits Its Receptor Patched by a Pincer Grasp Mechanism. Nat. Chem. Biol. 2019, 15, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Pereira de Jésus-Tran, K.; Côté, P.; Cantin, L.; Blanchet, J.; Labrie, F.; Breton, R. Comparison of Crystal Structures of Human Androgen Receptor Ligand-binding Domain Complexed with Various Agonists Reveals Molecular Determinants Responsible for Binding Affinity. Protein Sci. 2006, 15, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef] [PubMed]
- Izadi, S.; Anandakrishnan, R.; Onufriev, A.V. Building Water Models: A Different Approach. J. Phys. Chem. Lett. 2014, 5, 3863–3871. [Google Scholar] [CrossRef]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. Ff19SB: Amino-Acid-Specific Protein Backbone Parameters Trained against Quantum Mechanics Energy Surfaces in Solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Tomić, A.; Abramić, M.; Spoljarić, J.; Agić, D.; Smith, D.M.; Tomić, S. Human Dipeptidyl Peptidase III: Insights into Ligand Binding from a Combined Experimental and Computational Approach. J. Mol. Recognit. 2011, 24, 804–814. [Google Scholar] [CrossRef]
- Loncharich, R.J.; Brooks, B.R.; Pastor, R.W. Langevin Dynamics of Peptides: The Frictional Dependence of Isomerization Rates OfN-Acetylalanyl-N?-Methylamide. Biopolymers 1992, 32, 523–535. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Götz, A.W.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 2. Explicit Solvent Particle Mesh Ewald. J. Chem. Theory Comput. 2013, 9, 3878–3888. [Google Scholar] [CrossRef] [PubMed]
- Götz, A.W.; Williamson, M.J.; Xu, D.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 1. Generalized Born. J. Chem. Theory Comput. 2012, 8, 1542–1555. [Google Scholar] [CrossRef] [PubMed]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An Overview of the Amber Biomolecular Simulation Package. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Swanson, J.M.J.; Henchman, R.H.; McCammon, J.A. Revisiting Free Energy Calculations: A Theoretical Connection to MM/PBSA and Direct Calculation of the Association Free Energy. Biophys. J. 2004, 86, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.L. Analytical Molecular Surface Calculation. J. Appl. Crystallogr. 1983, 16, 548–558. [Google Scholar] [CrossRef]
- Siebold, C.; Rohatgi, R. The Inseparable Relationship Between Cholesterol and Hedgehog Signaling. Annu. Rev. Biochem. 2023, 92, 273–298. [Google Scholar] [CrossRef]
- Tukachinsky, H.; Kuzmickas, R.P.; Jao, C.Y.; Liu, J.; Salic, A. Dispatched and Scube Mediate the Efficient Secretion of the Cholesterol-Modified Hedgehog Ligand. Cell Rep. 2012, 2, 308–320. [Google Scholar] [CrossRef]
- Rozeboom, B.; Dey, N.; De, P. ER+ Metastatic Breast Cancer: Past, Present, and a Prescription for an Apoptosis-Targeted Future. Am. J. Cancer Res. 2019, 9, 2821–2831. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Time Interval/ns | (ΔHbind ± SD)/(kcal/mol) |
|---|---|---|
| ERα–estradiol | 0–500 | −12.87 ± 2.68 |
| ERα–cholesterol | −17.50 ± 3.42 | |
| PR–progesterone | −19.02 ± 2.97 | |
| PR–cholesterol | −19.48 ± 3.63 | |
| PR–SHH-N–cholesterol | 0–500 | −23.53 ± 8.33 |
| 300–500 | −20.99 ± 7.14 | |
| 500–1000 (a) * | −27.95 ± 7.80 | |
| 500–1000 (b) * | −23.25 ± 7.66 | |
| AR–SHH-N–cholesterol | 0–500 | −33.03 ± 10.27 |
| 300–500 | −29.73 ± 11.09 | |
| ERα–SHH-N–cholesterol | 0–300 | −20.70 ± 7.28 |
| 300–500 | −40.12 ± 9.25 | |
| 600–800 | −45.56 ± 9.5 | |
| 800–1000 | −36.29 ± 8.66 | |
| 1100–1350 | −42.40 ± 8.45 | |
| 1400–2000 | −51.51 ± 7.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomić, A.; Čonkaš, J.; Ozretić, P. Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Appl. Sci. 2024, 14, 562. https://doi.org/10.3390/app14020562
Tomić A, Čonkaš J, Ozretić P. Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Applied Sciences. 2024; 14(2):562. https://doi.org/10.3390/app14020562
Chicago/Turabian StyleTomić, Antonija, Josipa Čonkaš, and Petar Ozretić. 2024. "Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor" Applied Sciences 14, no. 2: 562. https://doi.org/10.3390/app14020562
APA StyleTomić, A., Čonkaš, J., & Ozretić, P. (2024). Let’s Talk about Sex Hormone Receptors and Their Physical Interaction with Sonic Hedgehog Protein: A Computational Study with Emphasis on Progesterone Receptor. Applied Sciences, 14(2), 562. https://doi.org/10.3390/app14020562