Image Analysis of Dynamic Brain Activity Based on Gray Distance Compensation


Abstract
:1. Introduction
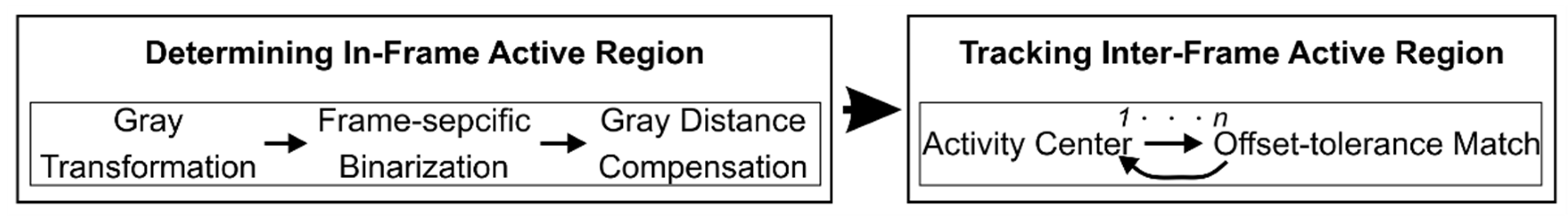
2. Automated Detection of Cellular Activity
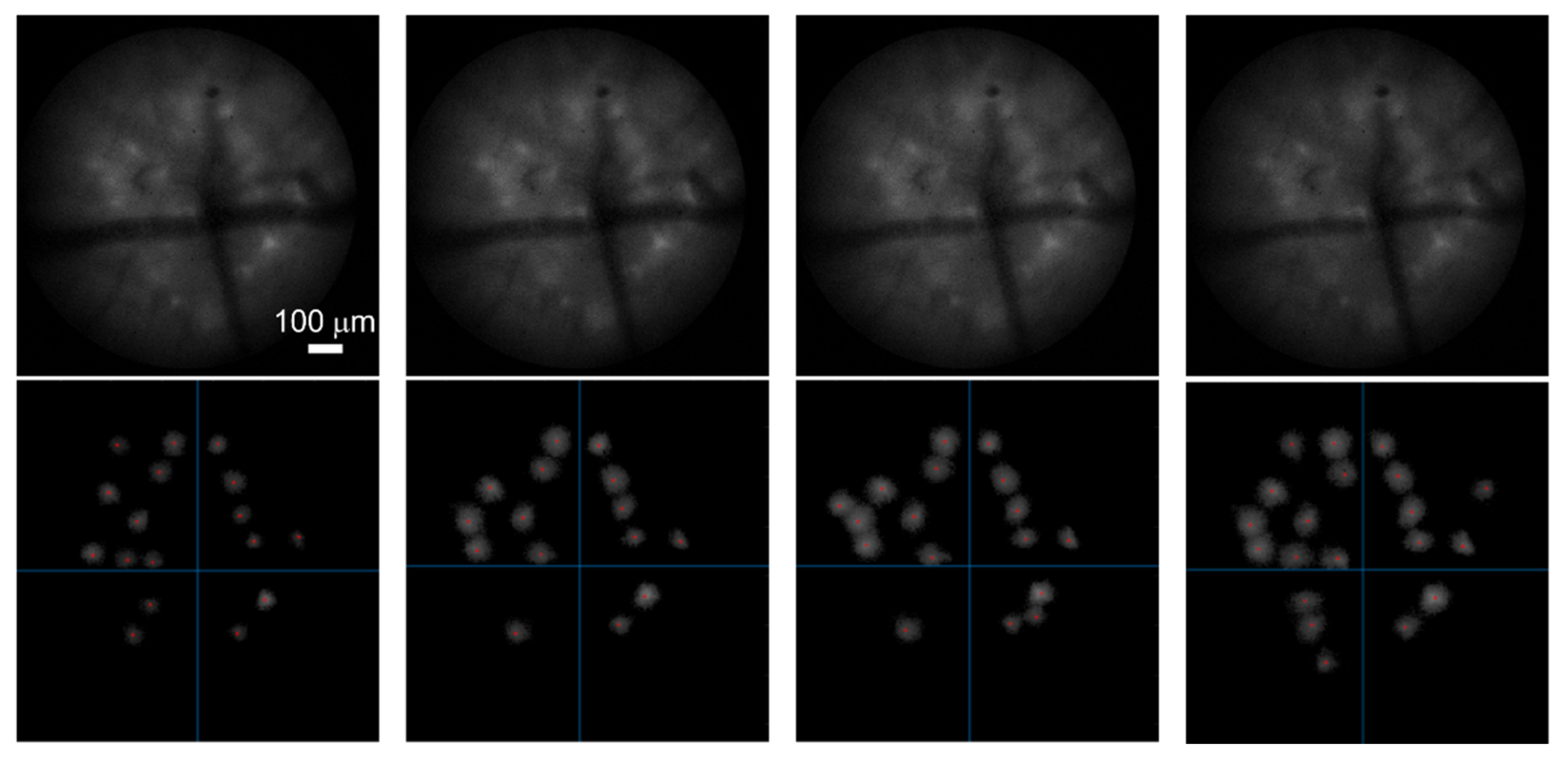
2.1. Detection of Active Regions
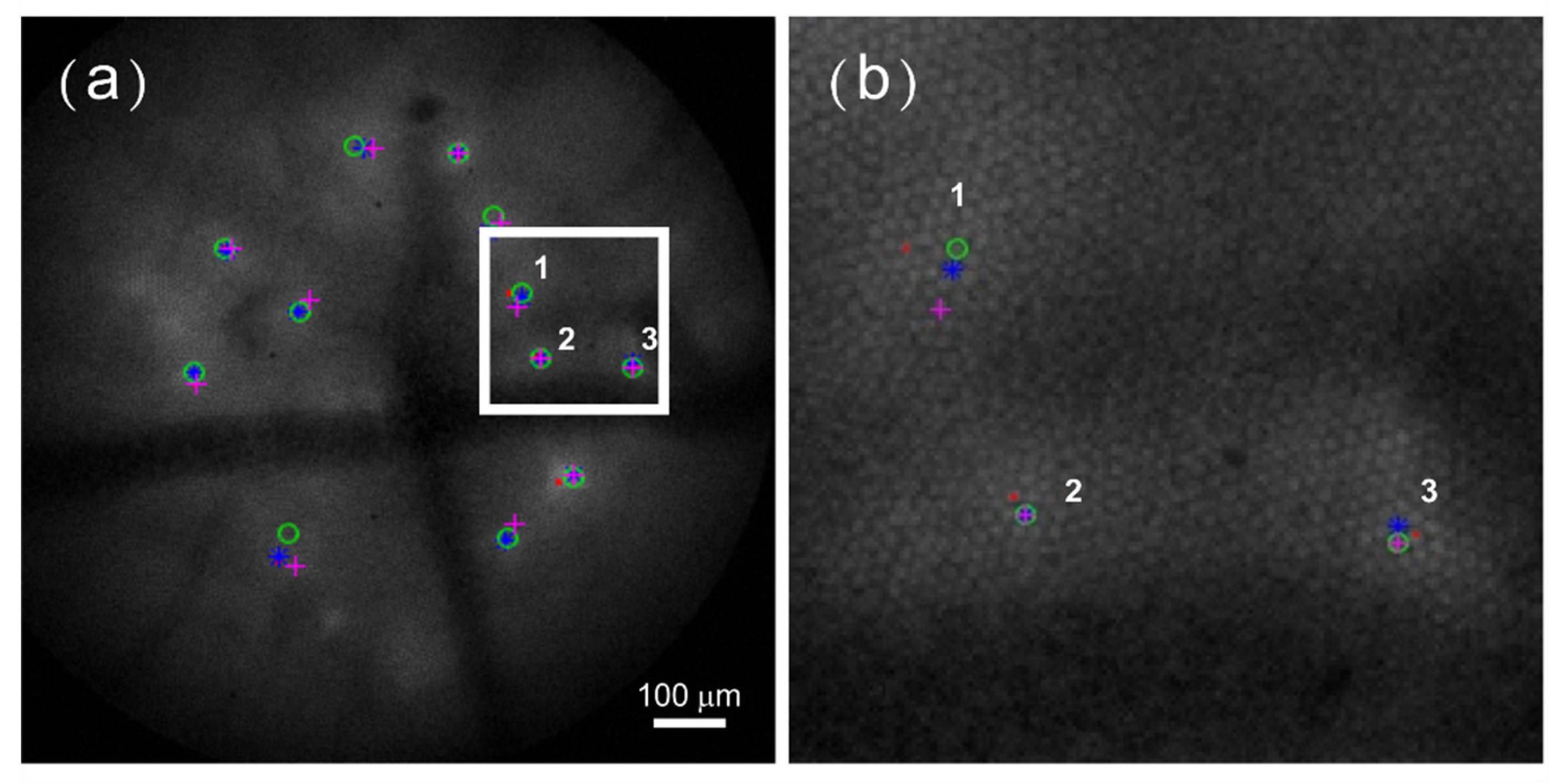
2.2. Activity Center Tracing in Serial Images
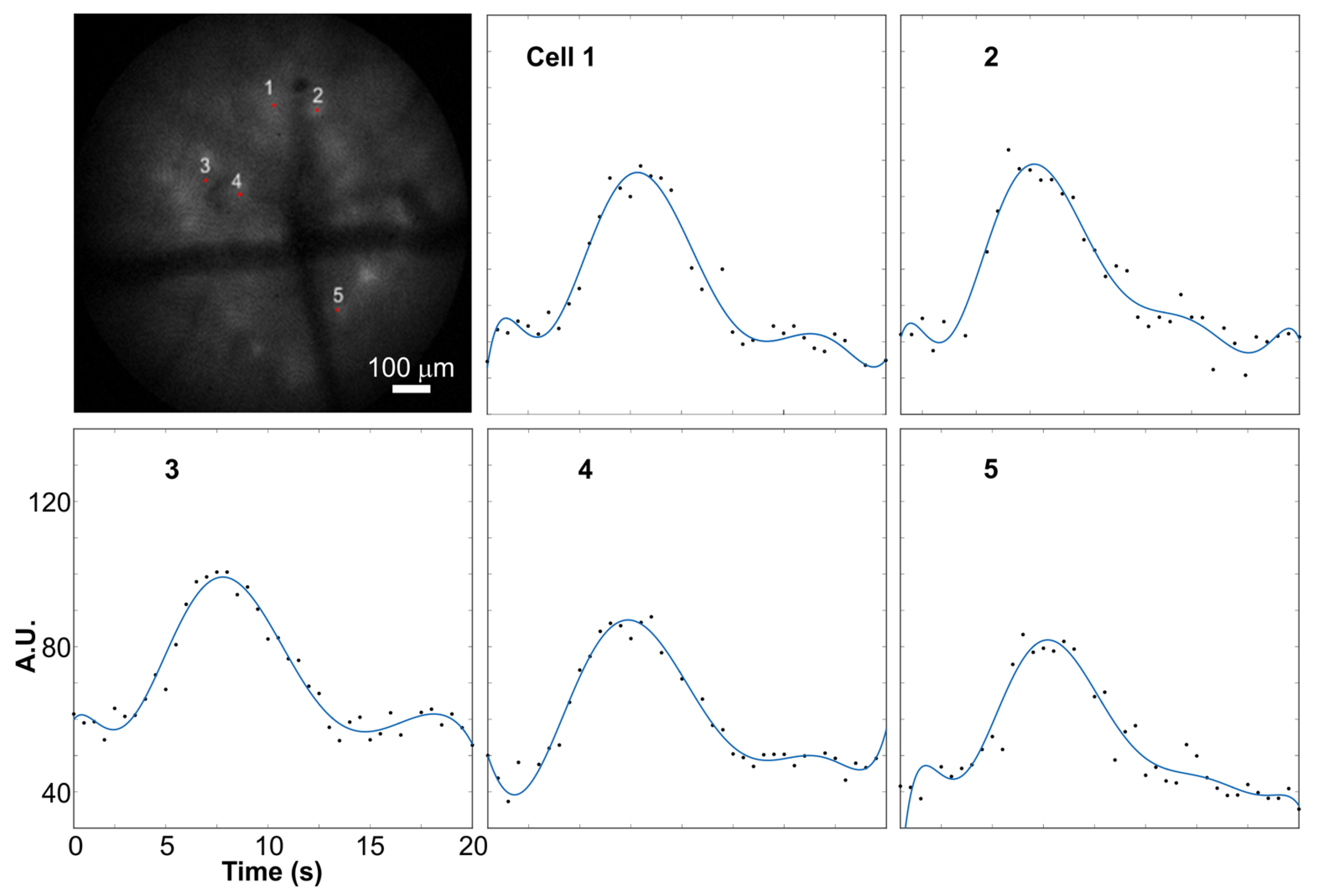
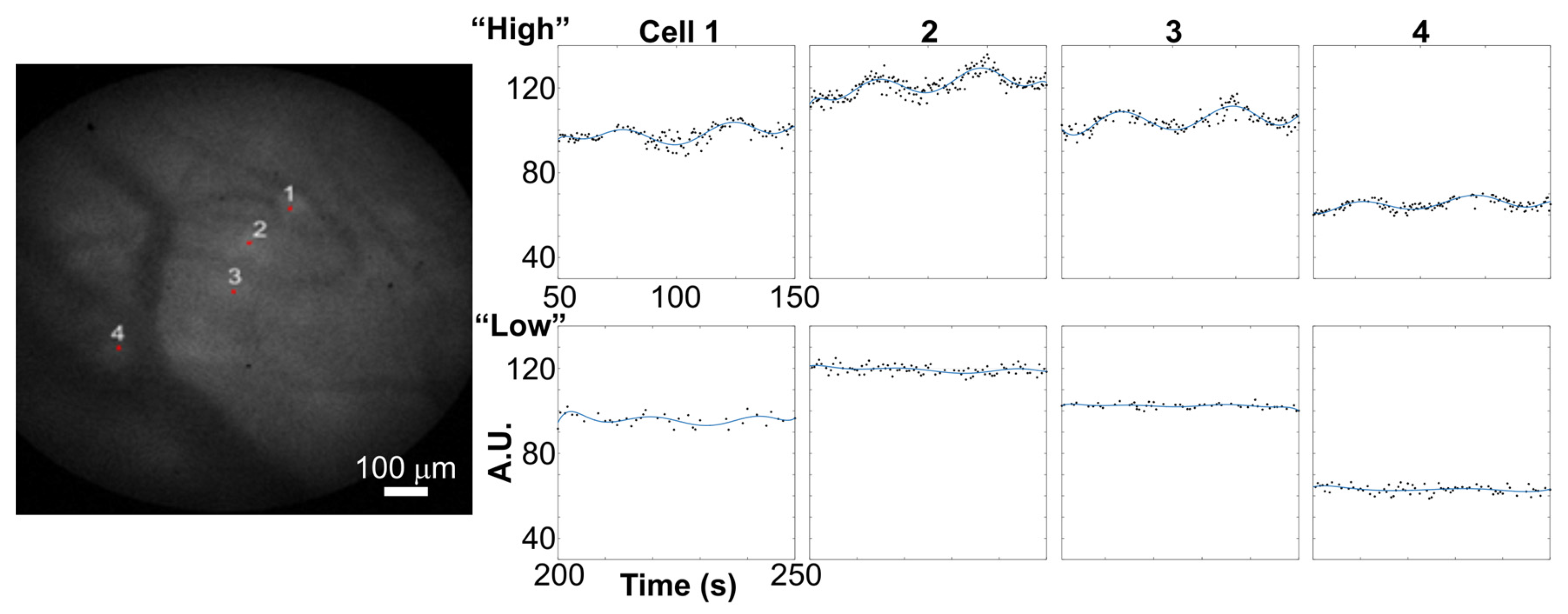
3. Performance of Analysis Routine on Distinct Imaging Datasets
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raichle, M.E. Functional brain imaging and human brain function. J. Neurosci. 2003, 23, 3959–3962. [Google Scholar] [PubMed]
- Ovsepian, S.V.; Dolly, J.O.; Zaborszky, L. Intrinsic voltage dynamics govern the diversity of spontaneous firing profiles in basal forebrain noncholinergic neurons. J. Neurophysiol. 2012, 108, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Paukert, M.; Agarwal, A.; Cha, J.; Doze, V.A.; Kang, J.U.; Bergles, D.E. Norepinephrine controls astroglial responsiveness to local circuit activity. Neuron 2014, 82, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Paukert, M.; Bergles, D.E.; Kang, J.U. Fiber optic fluorescence microscopy for functional brain imaging in awake mobile mice. Proc. SPIE 2014, 8928. [Google Scholar] [CrossRef]
- Cha, J.; Cheon, G.W.; Kang, J.U. Automated long-term tracking of freely moving animal and functional brain imaging based on fiber-optic microscopy. Proc. SPIE 2015, 931708. [Google Scholar] [CrossRef]
- Ding, F.; O’Donnell, J.; Thrane, A.S.; Zeppenfeld, D.; Kang, H.; Xie, L.; Wang, F.; Nedergaard, M. α1-Adrenergic receptors mediate coordinated Ca2+ signaling of cortical astrocytes in awake, behaving mice. Cell Calcium 2013, 54, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca2+ signaling in astrocytes from Ip3r2−/− mice in brain slices and during startle responses in vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Lv, X.L.; Wu, C.; Tang, H.C. Research on infrared image enhancement based on human visual system. Infrared Technol. 2010, 32, 708–712. [Google Scholar]
- Di, H.; Yu, Q.F.; Zhang, X.H. An algorithm for infrared image enhancement based on gray scale transform. J. Appl. Opt. 2006, 27, 12–14. [Google Scholar]
- Zhang, J.; Wang, Y. The infrared image segmentation based on distance-gray compensation and improved maximum entropy. Semicond. Optoelectron. 2016, 37, 126–130. [Google Scholar]
- Cheon, G.W.; Cha, J.; Kang, J.U. Random transverse motion-induced spatial compounding for fiber bundle imaging. Opt. Lett. 2014, 39, 4368–4371. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial set | Set 1 | Set 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Active regions # | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 |
| Matched frames # | 39 | 40 | 39 | 40 | 39 | 452 | 659 | 500 | 641 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Gau, Y.-T.A.; Le, H.N.D.; Bergles, D.E.; Kang, J.U. Image Analysis of Dynamic Brain Activity Based on Gray Distance Compensation. Appl. Sci. 2017, 7, 858. https://doi.org/10.3390/app7080858
Wang Y, Gau Y-TA, Le HND, Bergles DE, Kang JU. Image Analysis of Dynamic Brain Activity Based on Gray Distance Compensation. Applied Sciences. 2017; 7(8):858. https://doi.org/10.3390/app7080858
Chicago/Turabian StyleWang, Ying, Yung-Tian A. Gau, Hanh N. D. Le, Dwight E. Bergles, and Jin U. Kang. 2017. "Image Analysis of Dynamic Brain Activity Based on Gray Distance Compensation" Applied Sciences 7, no. 8: 858. https://doi.org/10.3390/app7080858
APA StyleWang, Y., Gau, Y. -T. A., Le, H. N. D., Bergles, D. E., & Kang, J. U. (2017). Image Analysis of Dynamic Brain Activity Based on Gray Distance Compensation. Applied Sciences, 7(8), 858. https://doi.org/10.3390/app7080858