A Synthetic Nervous System Controls a Simulated Cockroach

Abstract
:1. Introduction
2. Methods
2.1. Modeling Overview
2.2. System Dynamics
2.2.1. Neuron Dynamics
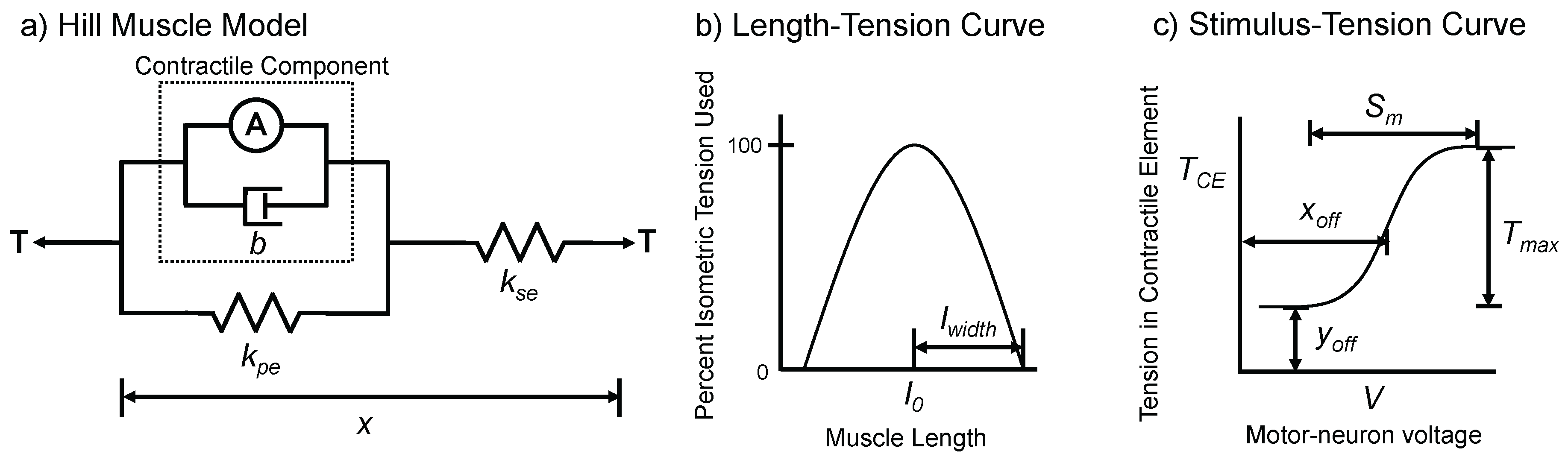
2.2.2. Muscle and Joint Dynamics
2.3. Physical Model
2.4. Neural Network as Control System
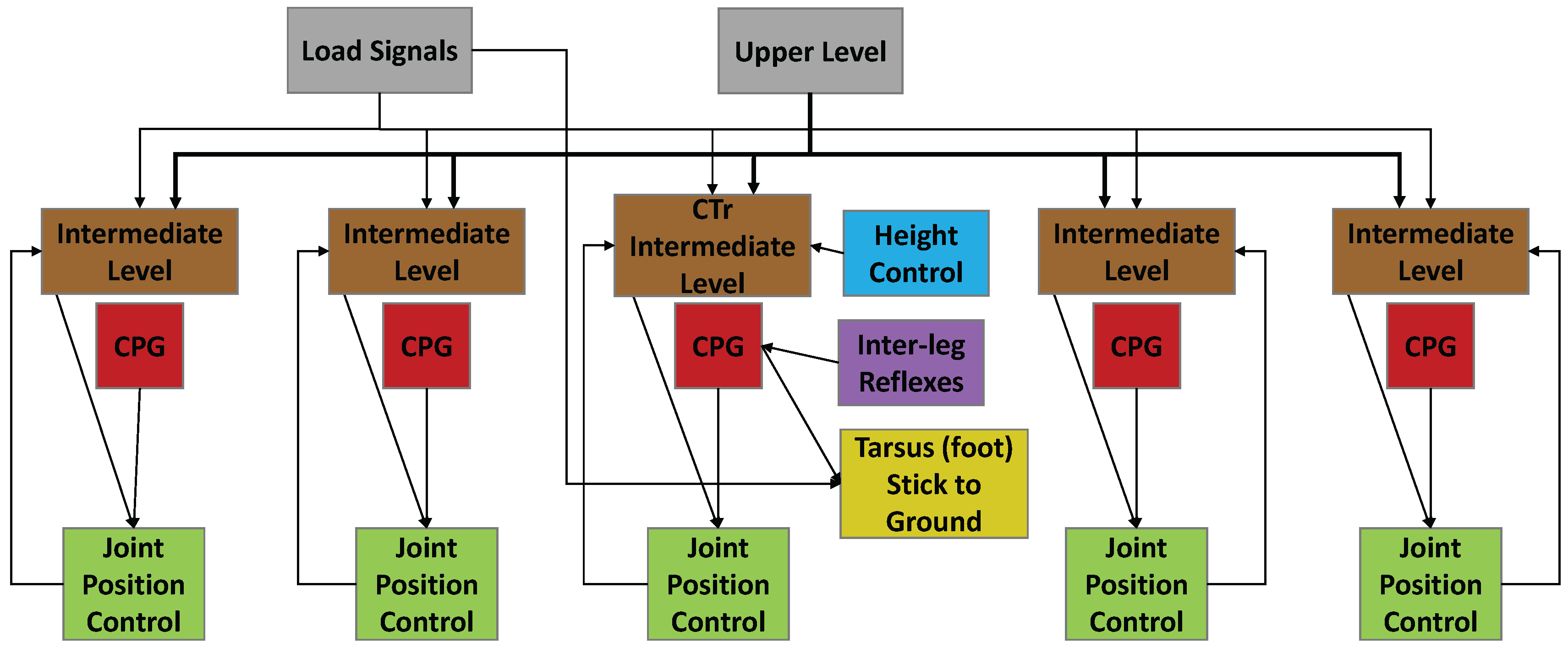
2.4.1. Overview of Synthetic Nervous System
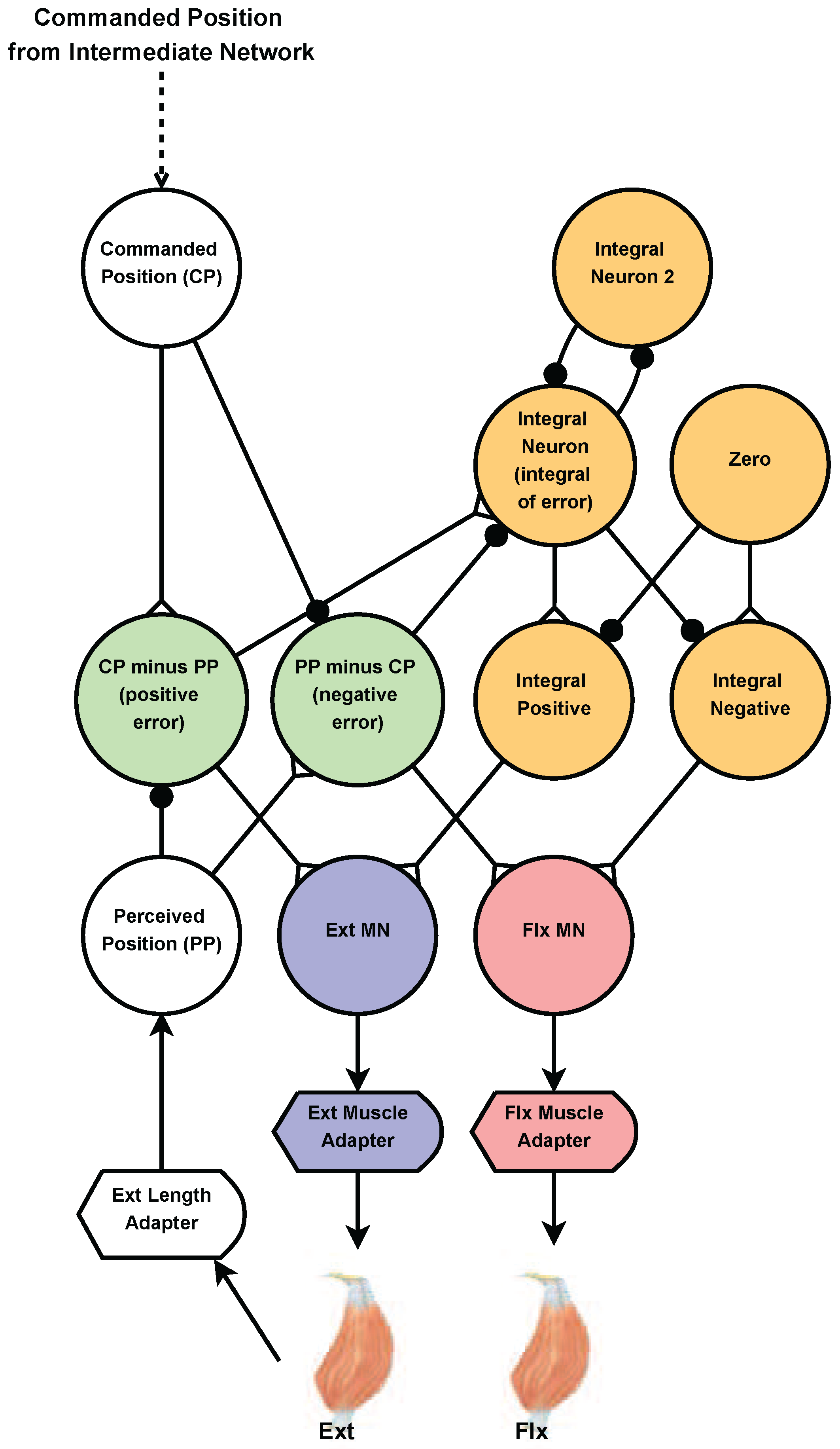
2.4.2. Joint Position Control
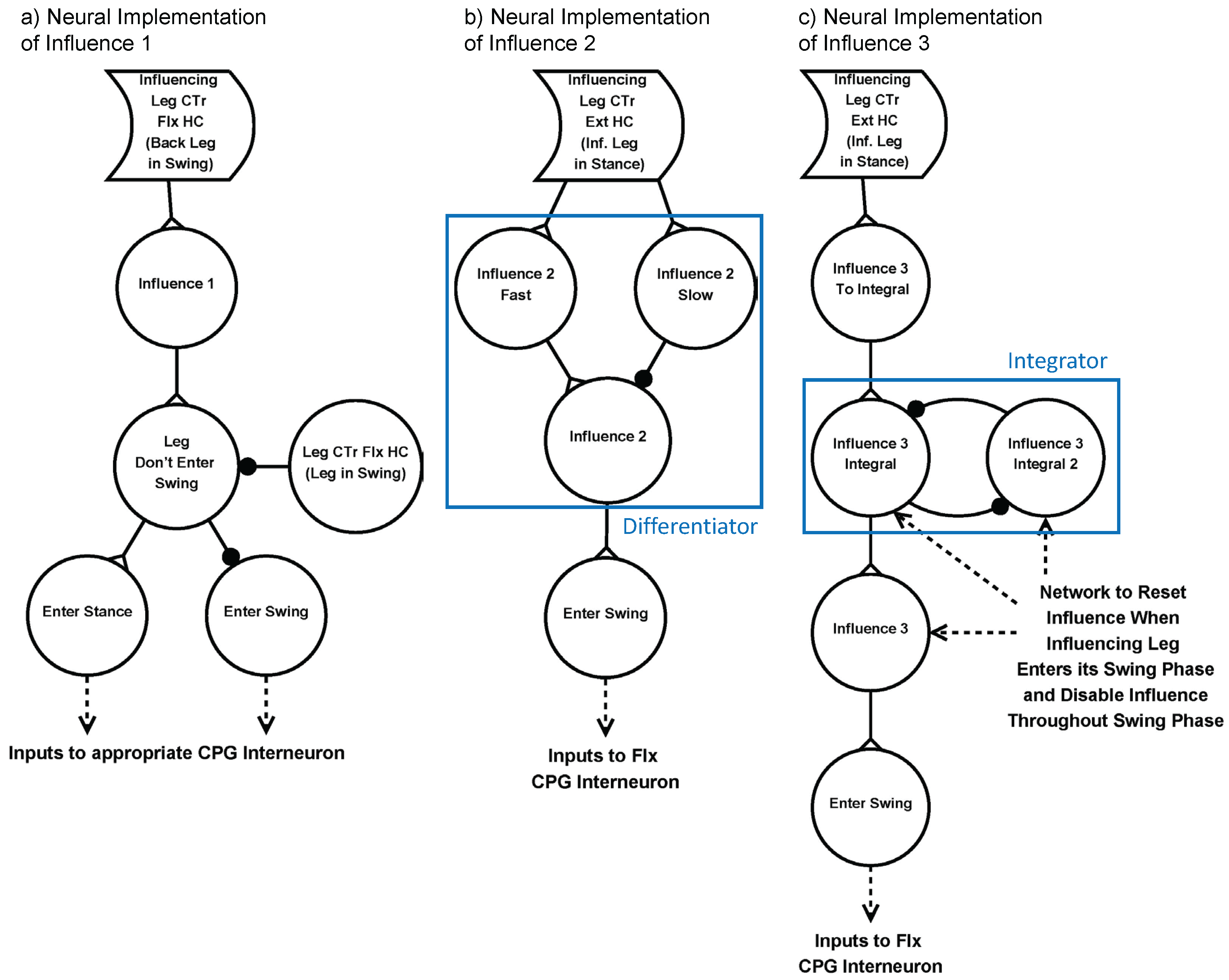
2.4.3. Inter-Leg Coordination
2.4.4. Height Control
2.5. Design Process for a Single Joint
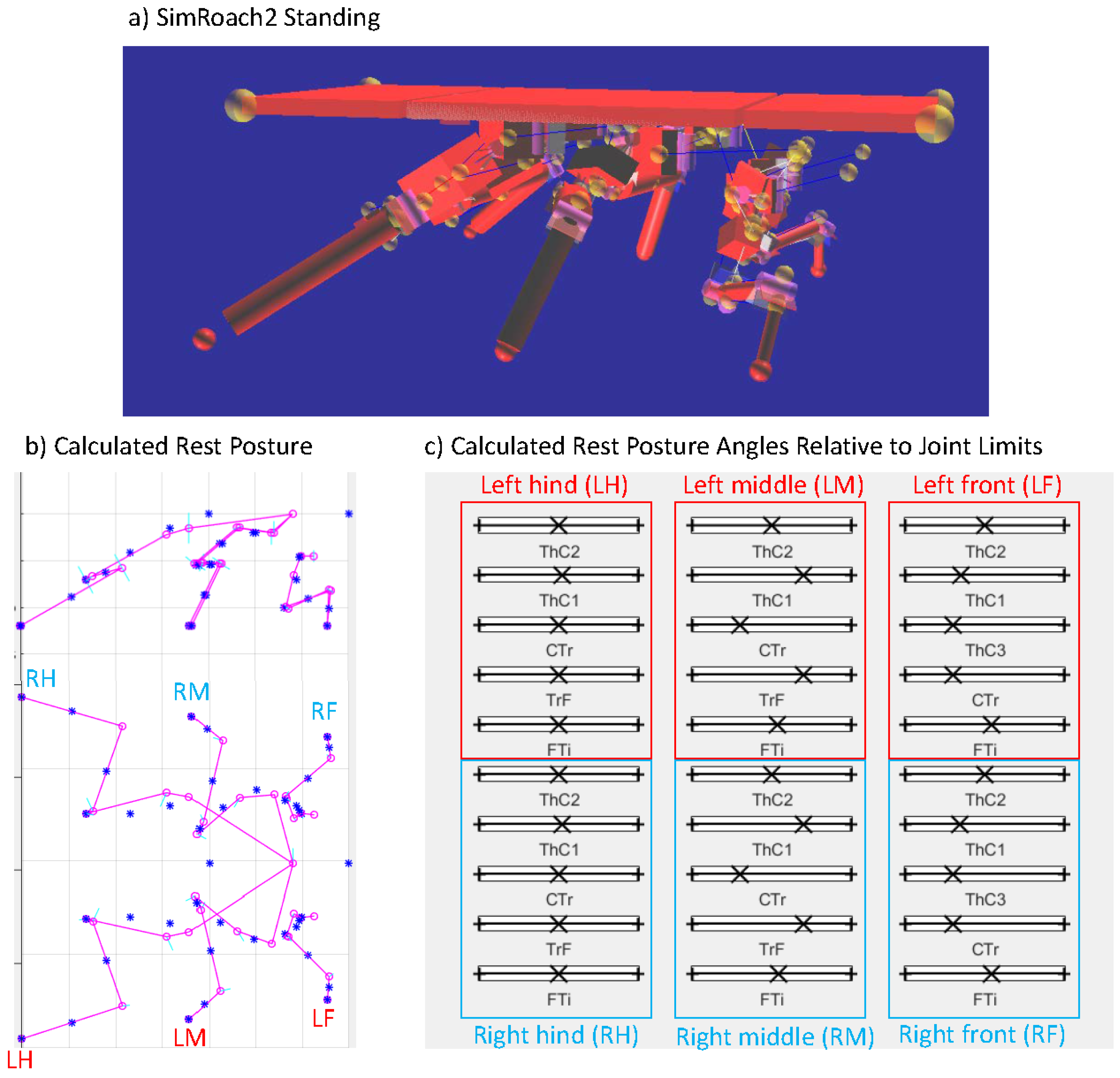
2.5.1. Resting Posture
2.5.2. Calculation of Passive Spring Stiffness and Damping Coefficients
2.5.3. Passive Muscle Force Parameters
2.5.4. Active Muscle Parameters
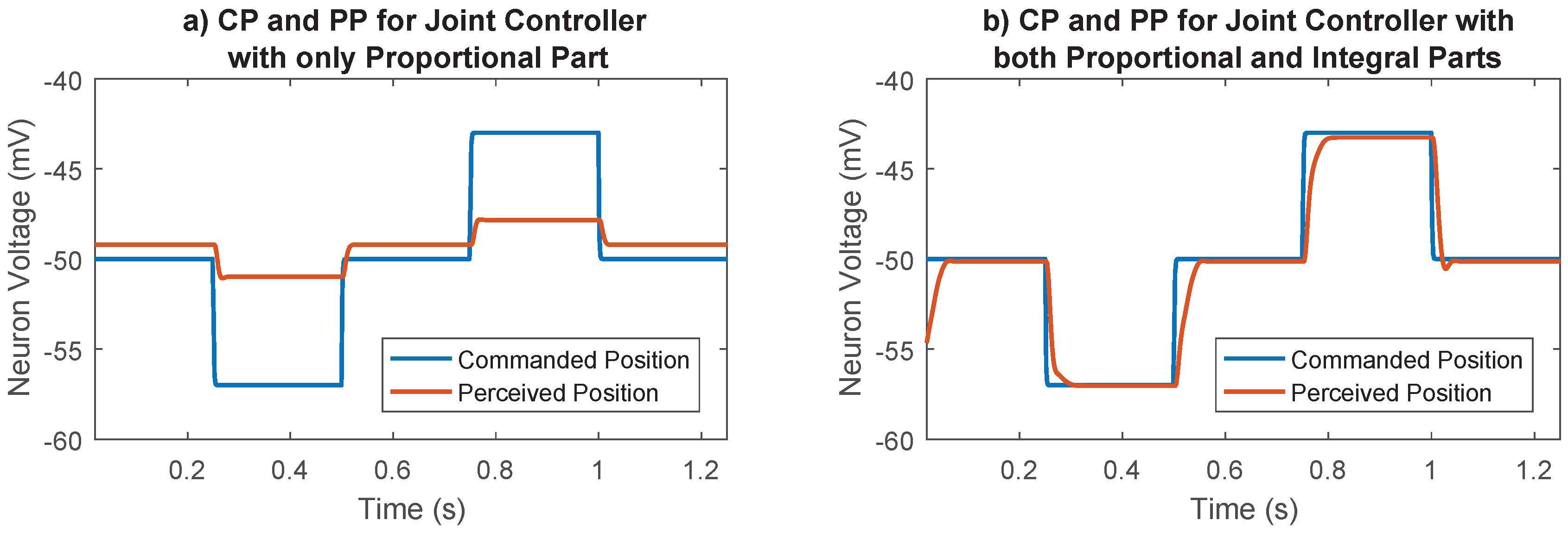
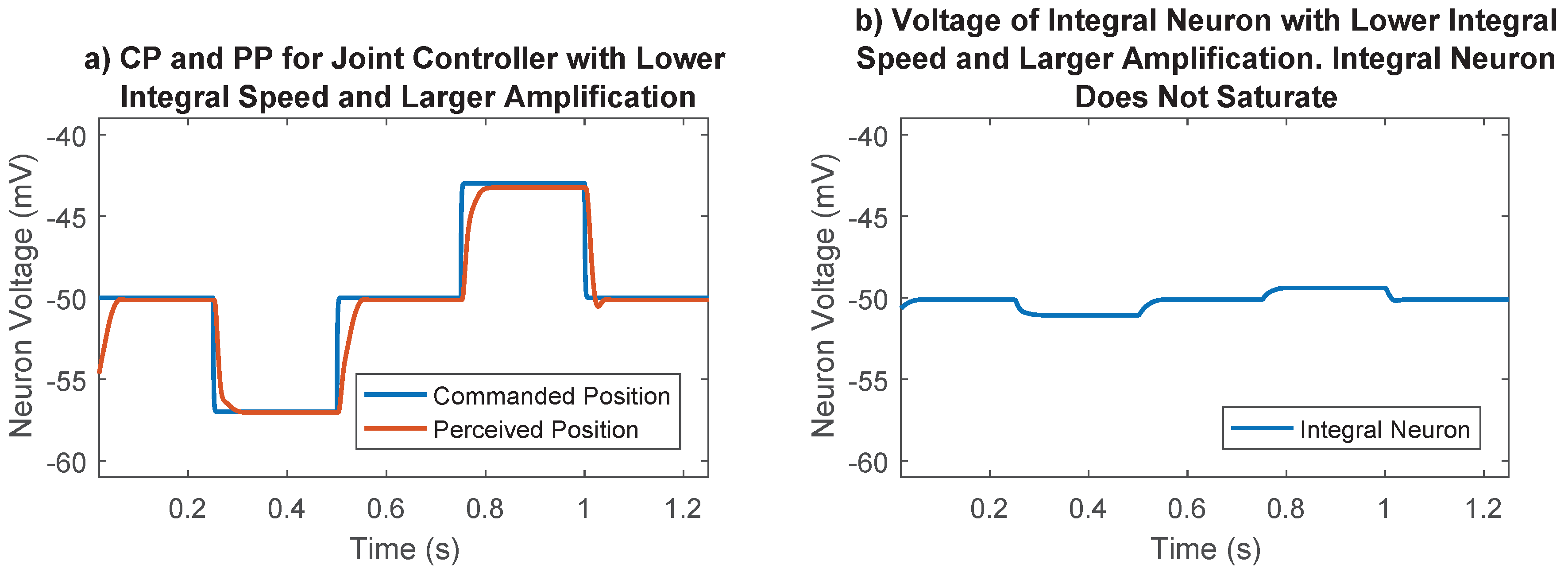
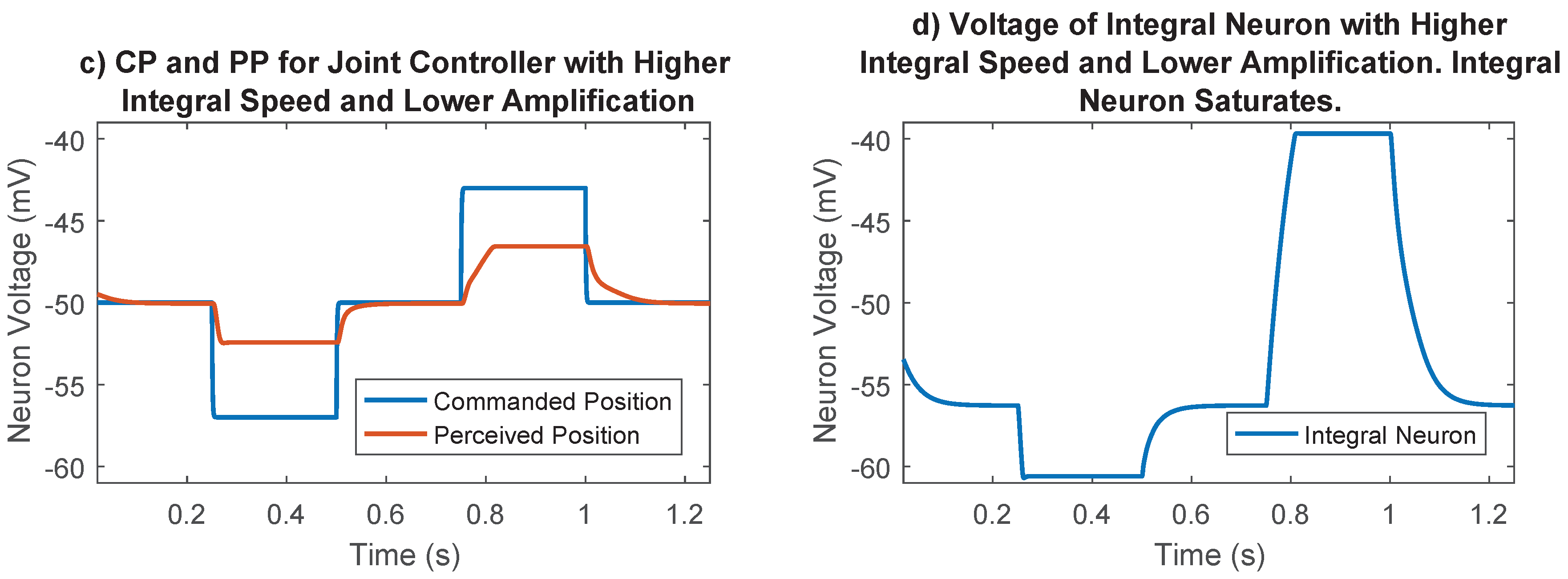
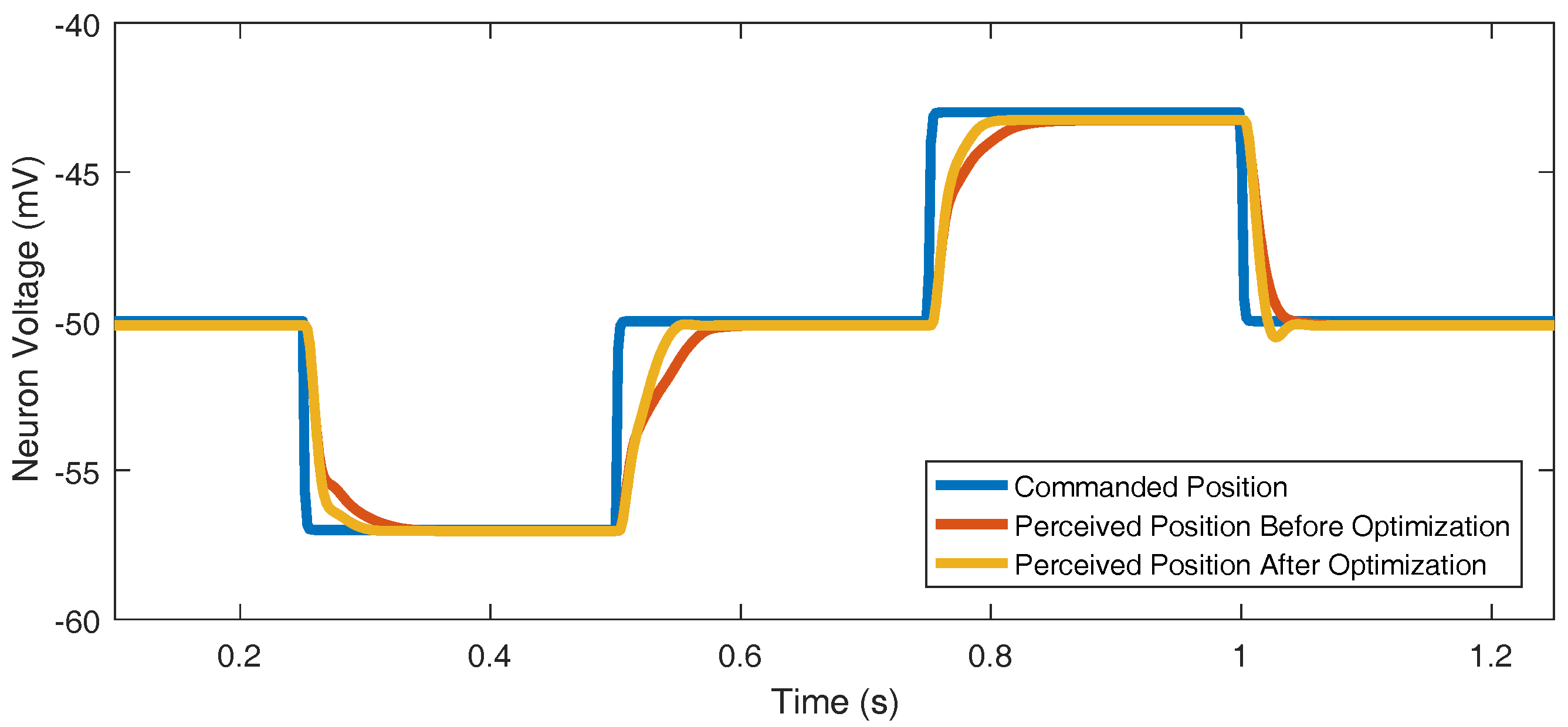
2.5.5. Synaptic Strength to Position Control Integral Neuron
2.5.6. Optimization Process
3. Results
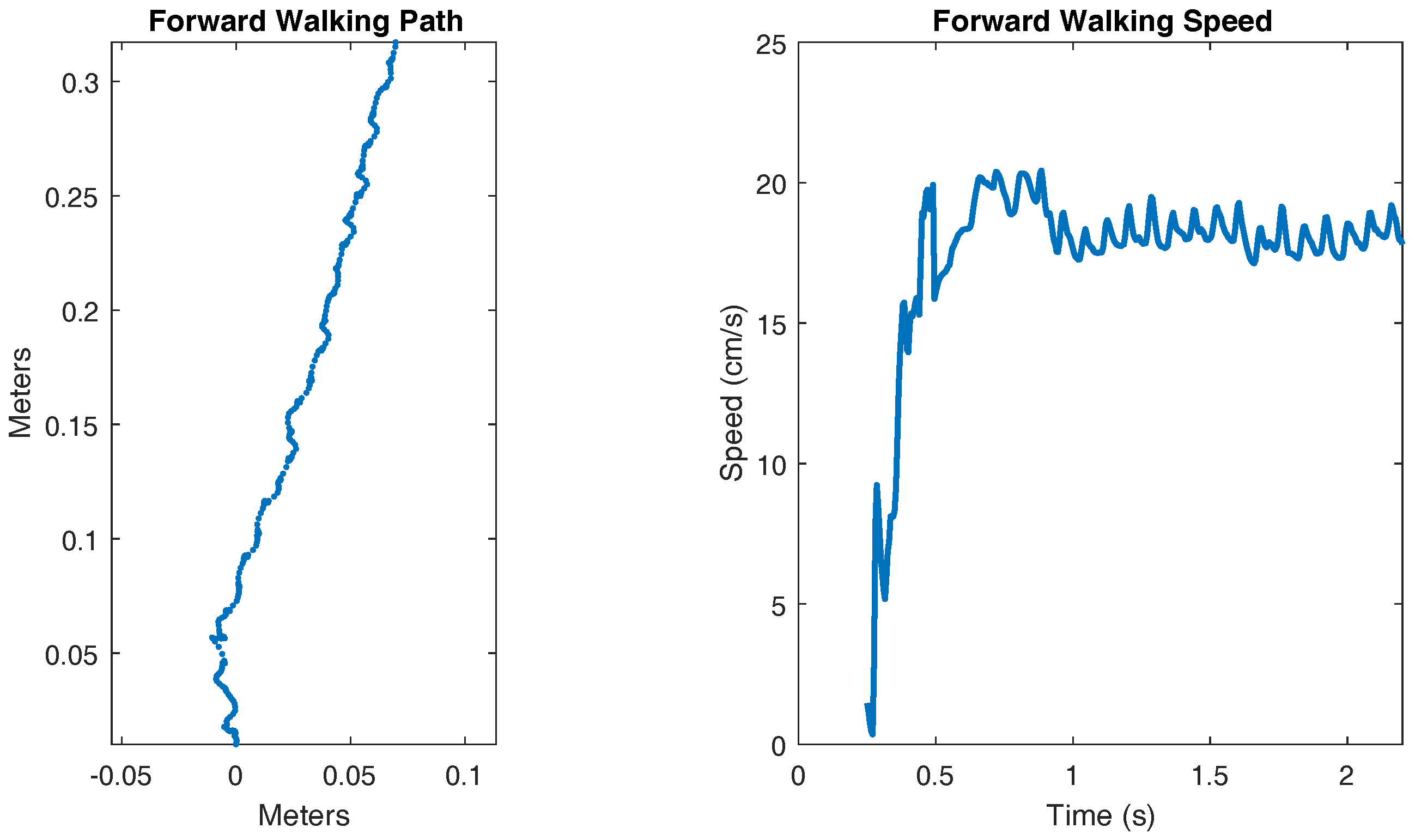
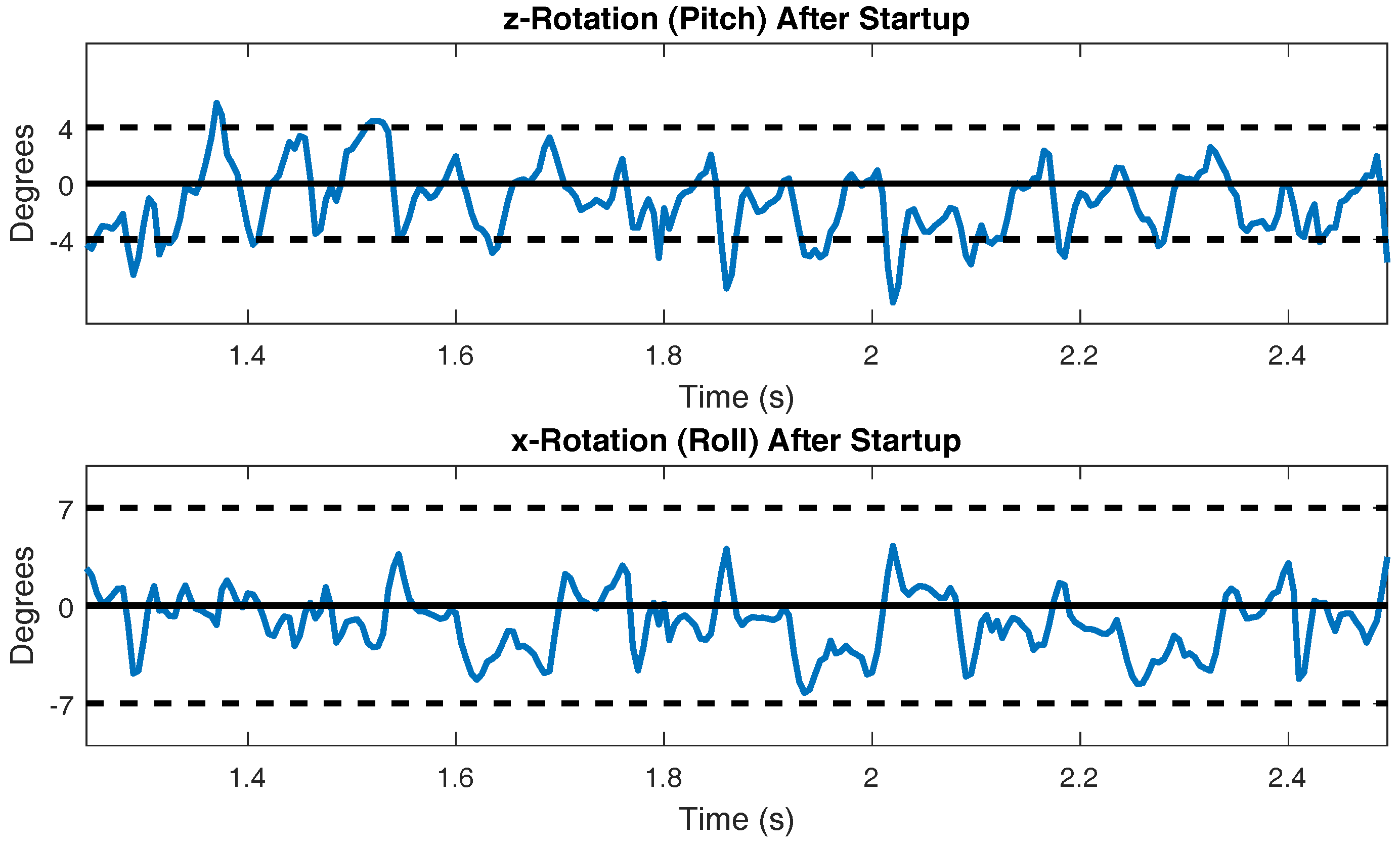
3.1. Forward Walking
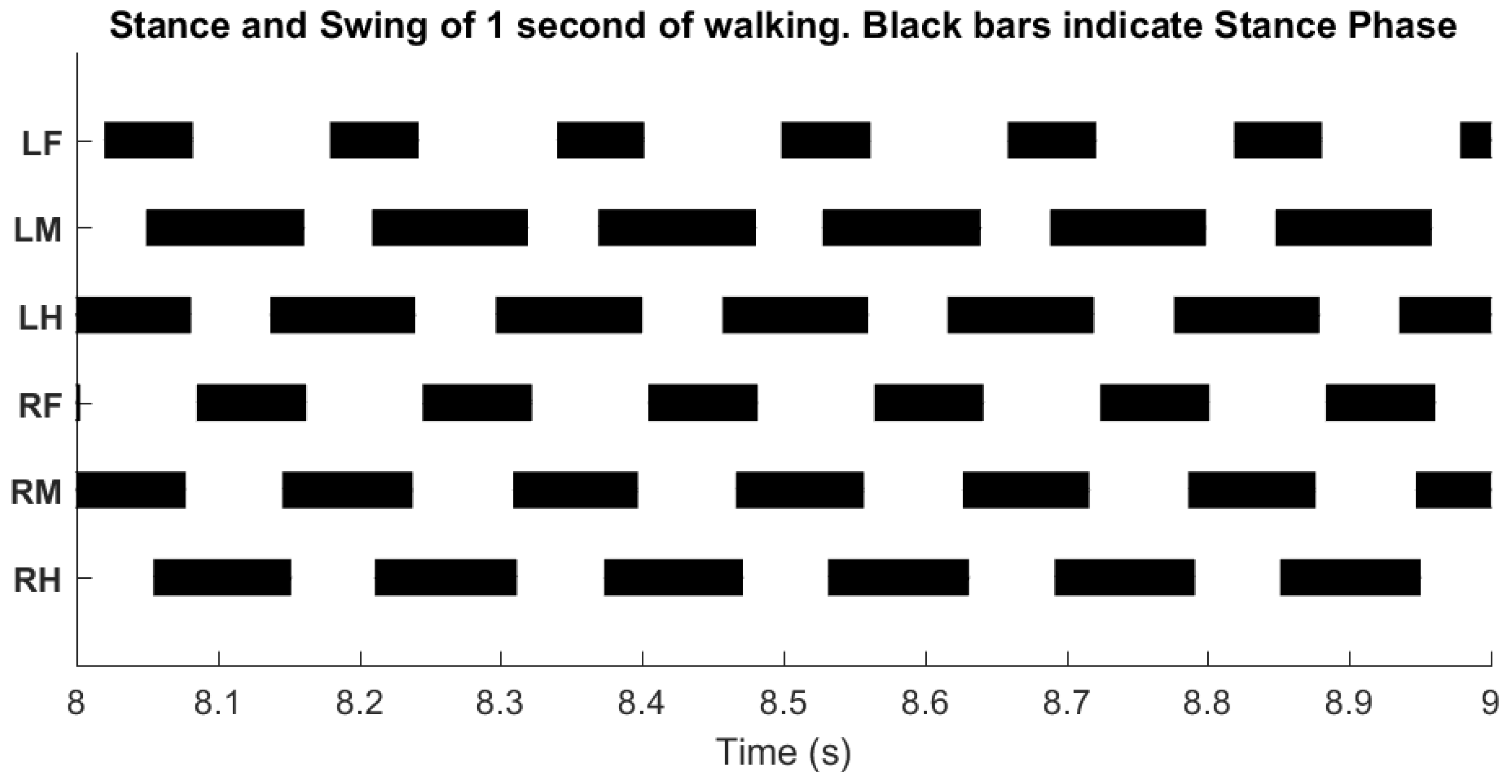
3.2. Inter-Leg Coordination
4. Discussion and Future Work
4.1. Importance of Simulated Animats for Biologically-Inspired Robots
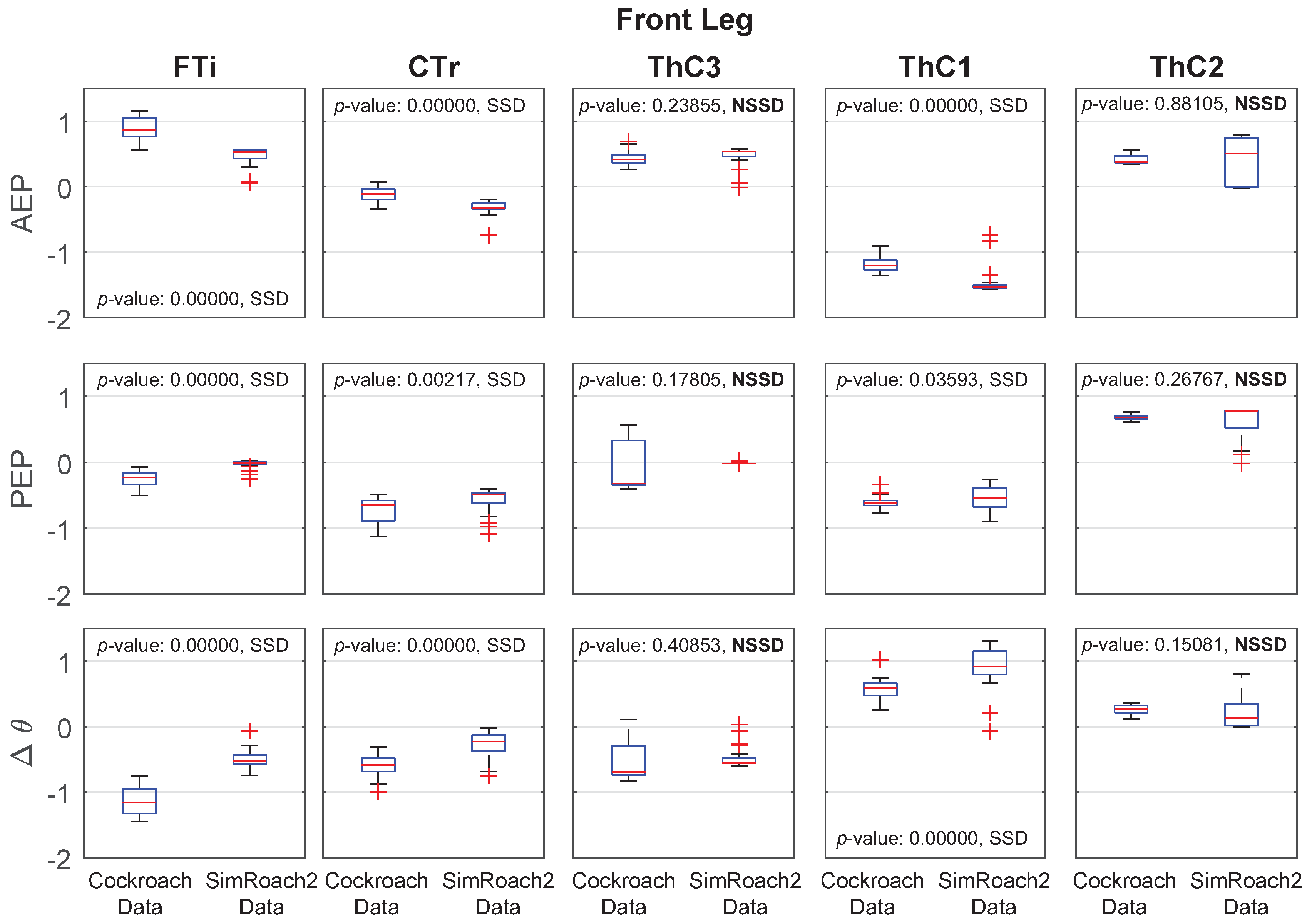
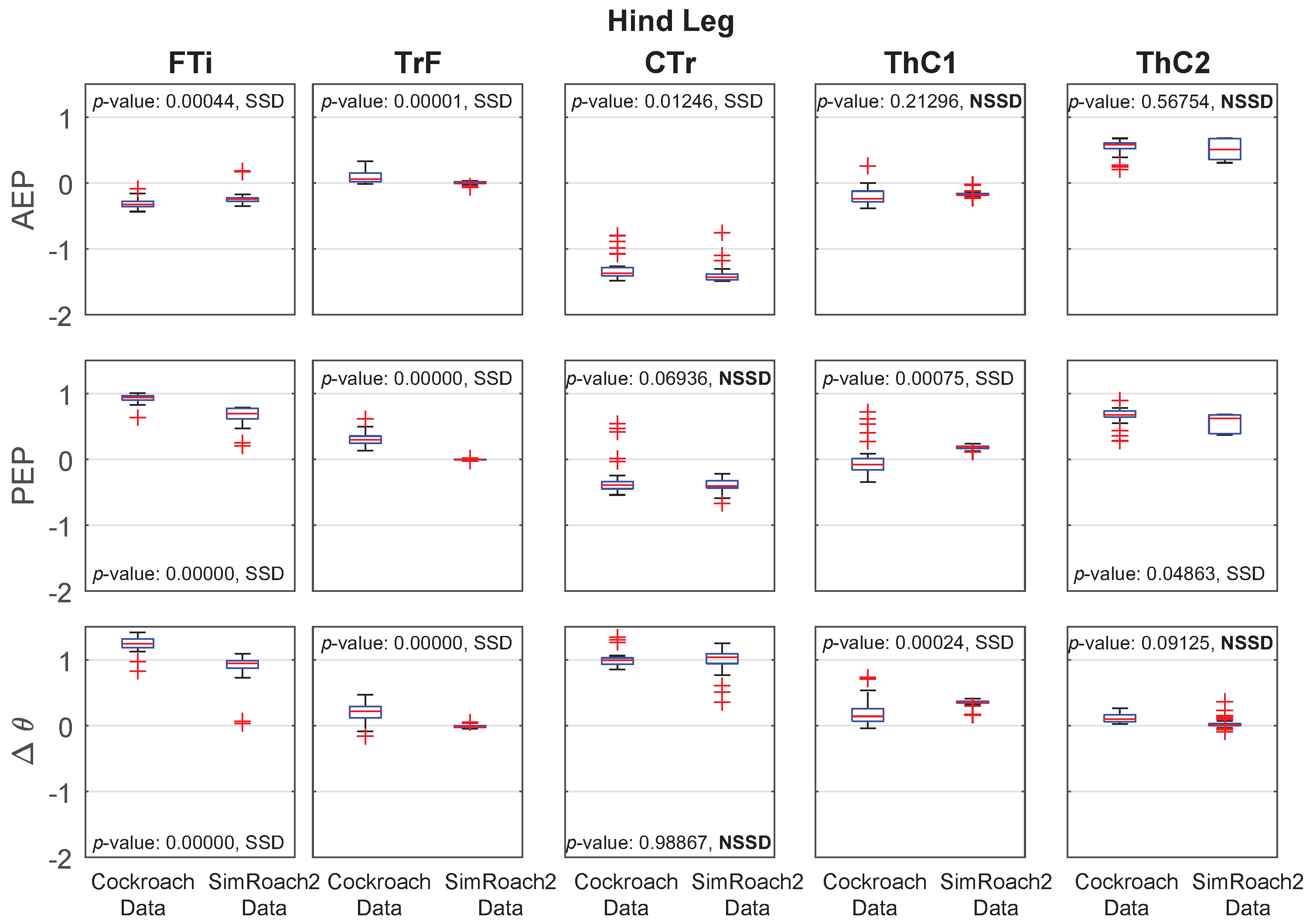
4.2. Walking Comparison between SimRoach2 and a Cockroach
Joint Angle Comparison
4.3. Redundant Parameters
4.4. Velocity Control for Muscles
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
| AEP | Anterior extreme position |
| ANN | Artificial neural network |
| CP | Commanded position |
| CPG | Central pattern generator |
| CTr | Coxa-trochanter |
| CX | Central complex |
| DOF | Degree of freedom |
| FSM | Finite state machine |
| FTi | Femur-tibia |
| HH | Hodgkin–Huxley |
| MN | Motor-neuron |
| PEP | Posterior extreme position |
| PI | Proportional-integral |
| PP | Perceived position |
| SNS | Synthetic nervous system |
| ThC | Thorax-coxa |
| TrF | Trochanter-femur |
References
- Ryckebusch, S.; Laurent, G. Rhythmic patterns evoked in locust leg motor neurons by the muscarinic agonist pilocarpine. J. Neurophysiol. 1993, 69, 1583–1595. [Google Scholar] [PubMed]
- Büschges, A.; Schmitz, J.; Bässler, U. Rhythmic patterns in the thoracic nerve cord of the stick insect induced by pilocarpine. J. Exp. Biol. 1995, 198, 435–456. [Google Scholar]
- Ryckebusch, S.; Laurent, G. Interactions between segmental leg central pattern generators during fictive rhythms in the locust. J. Neurophysiol. 1994, 72, 2771–2785. [Google Scholar] [PubMed]
- Noah, J.A.; Quimby, L.; Frazier, S.F.; Zill, S.N. Sensing the effect of body load in legs: Responses of tibial campaniform sensilla to forces applied to the thorax in freely standing cockroaches. J. Comp. Physiol. A 2004, 190, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Zill, S.; Schmitz, J.; Büschges, A. Load sensing and control of posture and locomotion. Arthropod Struct. Dev. 2004, 33, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Akay, T.; Haehn, S.; Schmitz, J.; Büschges, A. Signals From Load Sensors Underlie Interjoint Coordination During Stepping Movements of the Stick Insect Leg. J. Neurophysiol. 2004, 92, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Bucher, D.; Akay, T.; DiCaprio, R.a.; Buschges, A. Interjoint coordination in the stick insect leg-control system: The role of positional signaling. J. Neurophysiol. 2003, 89, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Hess, D.; Büschges, A. Role of Proprioceptive Signals From an Insect Femur-Tibia Joint in Patterning Motoneuronal Activity of an Adjacent Leg Joint. J. Neurophysiol. 1999, 81, 1856–1865. [Google Scholar] [PubMed]
- Mu, L.; Ritzmann, R.E. Kinematics and motor activity during tethered walking and turning in the cockroach, Blaberus discoidalis. J. Comp. Physiol. A 2005, 191, 1037–1054. [Google Scholar] [CrossRef] [PubMed]
- Hellekes, K.; Blincow, E.; Hoffmann, J.; Buschges, A. Control of reflex reversal in stick insect walking: Effects of intersegmental signals, changes in direction, and optomotor-induced turning. J. Neurophysiol. 2012, 107, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.P.; Guo, P.; Mu, L.; Harley, C.M.; Ritzmann, R.E. Central-Complex Control of Movement in the Freely Walking Cockroach. Curr. Biol. 2015, 25, 2795–2803. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, T.; Ewald, A.; von Twickel, A.; Büschges, A. Controlling legs for locomotion-insights from robotics and neurobiology. Bioinspir. Biomim. 2015, 10, 041001. [Google Scholar] [CrossRef] [PubMed]
- Daun-Gruhn, S. A mathematical modeling study of inter-segmental coordination during stick insect walking. J. Comput. Neurosci. 2011, 30, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Brown, A.E.; Bender, J.A.; Quinn, R.D.; Ritzmann, R.E. A neuromechanical simulation of insect walking and transition to turning of the cockroach Blaberus discoidalis. Biol. Cybern. 2014, 108, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H.; Kindermann, T.; Schumm, M.; Dean, J.; Schmitz, J. Walknet - A biologically inspired network to control six-legged walking. Neural Netw. 1998, 11, 1435–1447. [Google Scholar] [CrossRef]
- Schilling, M.; Paskarbeit, J.; Hoinville, T.; Hüffmeier, A.; Schneider, A.; Schmitz, J.; Cruse, H. A hexapod walker using a heterarchical architecture for action selection. Front. Comput. Neurosci. 2013, 7, 126. [Google Scholar] [CrossRef] [PubMed]
- Ekeberg, Ö.; Blümel, M.; Büschges, A. Dynamic simulation of insect walking. Arthropod Struct. Dev. 2004, 33, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Rutter, B.L.; Taylor, B.K.; Bender, J.A.; Blümel, M.; Lewinger, W.A.; Ritzmann, R.E.; Quinn, R.D. Descending commands to an insect leg controller network cause smooth behavioral transitions. In Proceedings of the IEEE International Conference on Intelligent Robots and Systems, San Francisco, CA, USA, 25–30 September 2011; pp. 215–220. [Google Scholar]
- Daun-Gruhn, S.; Tóth, T.I. An inter-segmental network model and its use in elucidating gait-switches in the stick insect. J. Comput. Neurosci. 2011, 31, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Toth, T.I.; Schmidt, J.; Büschges, A.; Daun-Gruhn, S. A Neuro-Mechanical Model of a Single Leg Joint Highlighting the Basic Physiological Role of Fast and Slow Muscle Fibres of an Insect Muscle System. PLoS ONE 2013, 8, e78247. [Google Scholar] [CrossRef] [PubMed]
- Tóth, T.I.; Grabowska, M.; Rosjat, N.; Hellekes, K.; Borgmann, A.; Daun-Gruhn, S. Investigating inter-segmental connections between thoracic ganglia in the stick insect by means of experimental and simulated phase response curves. Biol. Cybern. 2015, 109, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Schilling, M.; Hoinville, T.; Schmitz, J.; Cruse, H. Walknet, a bio-inspired controller for hexapod walking. Biol. Cybern. 2013, 107, 397–419. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.; Schneider, A.; Schilling, M.; Cruse, H. No need for a body model: Positive velocity feedback for the control of an 18-DOF robot walker. Appl. Bionics Biomech. 2008, 5, 135–147. [Google Scholar] [CrossRef]
- Dupeyroux, J.; Passault, G.; Ruffier, F.; Viollet, S.; Serres, J. Hexabot: A small 3D-printed six-legged walking robot designed for desert ant-like navigation tasks. In Proceedings of the IFAC Word Congress, Toulouse, France, 9–14 July 2017. [Google Scholar]
- Szczecinski, N.S.; Getsy, A.P.; Martin, J.P.; Ritzmann, R.E.; Quinn, R.D. Mantisbot is a robotic model of visually guided motion in the praying mantis. Arthropod Struct. Dev. 2017, 46, 736–751. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Quinn, R.D. Template for the neural control of directed stepping generalized to all legs of MantisBot. Bioinspir. Biomim. 2017, 12, 045001. [Google Scholar] [CrossRef] [PubMed]
- Gruhn, M.; Hoffmann, O.; Dübbert, M.; Scharstein, H.; Büschges, A. Tethered stick insect walking: A modified slippery surface setup with optomotor stimulation and electrical monitoring of tarsal contact. J. Neurosci. Methods 2006, 158, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Durr, V. The behavioural transition from straight to curve walking: Kinetics of leg movement parameters and the initiation of turning. J. Exp. Biol. 2005, 208, 2237–2252. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H. What mechanisms coordinate leg movement in walking arthropods? Trends Neurosci. 1990, 13, 15–21. [Google Scholar] [CrossRef]
- Cofer, D.; Cymbalyuk, G.; Reid, J.; Zhu, Y.; Heitler, W.J.; Edwards, D.H. AnimatLab: A 3D graphics environment for neuromechanical simulations. J. Neurosci. Methods 2010, 187, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Hunt, A.J.; Quinn, R.D. Design process and tools for dynamic neuromechanical models and robot controllers. Biol. Cybern. 2017, 111, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. Bull. Math. Biol. 1990, 52, 25–71. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V. The Heat of Shortening and the Dynamic Constants of Muscle. Proc. R. Soc. B Biol. Sci. 1938, 126, 136–195. [Google Scholar] [CrossRef]
- Shadmehr, R.; Arbib, M.A. A mathematical analysis of the force-stiffness characteristics of muscles in control of a single joint system. Biol. Cybern. 1992, 66, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Hooper, S.L.; Guschlbauer, C.; Blumel, M.; Rosenbaum, P.; Gruhn, M.; Akay, T.; Buschges, A. Neural Control of Unloaded Leg Posture and of Leg Swing in Stick Insect, Cockroach, and Mouse Differs from That in Larger Animals. J. Neurosci. 2009, 29, 4109–4119. [Google Scholar] [CrossRef] [PubMed]
- Ache, J.M.; Matheson, T. Passive Joint Forces Are Tuned to Limb Use in Insects and Drive Movements without Motor Activity. Curr. Biol. 2013, 23, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Zill, S.N.; Chaudhry, S.; Exter, A.; Büschges, A.; Schmitz, J. Positive force feedback in development of substrate grip in the stick insect tarsus. Arthropod Struct. Dev. 2014, 43, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Paskarbeit, J.; Otto, M.; Schilling, M.; Schneider, A. Stick(y) Insects—Evaluation of Static Stability for Bio-inspired Leg Coordination in Robotics. In Proceedings of the Conference on Biomimetic and Biohybrid Systems, Edinburgh, UK, 19–22 July 2016; Volume 1, pp. 239–250. [Google Scholar]
- Ramdya, P.; Thandiackal, R.; Cherney, R.; Asselborn, T.; Benton, R.; Ijspeert, A.J.; Floreano, D. Climbing favours the tripod gait over alternative faster insect gaits. Nat. Commun. 2017, 8, 14494. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Hunt, A.J.; Quinn, R.D. A functional subnetwork approach to designing synthetic nervous systems that control legged robot locomotion. Front. Neurorobot. 2017, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bässler, D.; Büschges, A.; Meditz, S.; Bässler, U. Correlation between muscle structure and filter characteristics of the muscle-joint system in three orthopteran insect species. J. Exp. Biol. 1996, 199, 2169–2183. [Google Scholar]
- Wolf, H. Inhibitory motoneurons in arthropod motor control: Organisation, function, evolution. J. Comp. Physiol. A 2014, 200, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Hooper, S.L.; Guschlbauer, C.; von Uckermann, G.; Buschges, A. Different Motor Neuron Spike Patterns Produce Contractions With Very Similar Rises in Graded Slow Muscles. J. Neurophysiol. 2007, 97, 1428–1444. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanz, M. Chapter 3 P.I.D. control: Structure. In EECS 475 Applied Control; Case Western Reserve University: Cleveland, OH, USA, 2016. [Google Scholar]
- Dürr, V.; Schmitz, J.; Cruse, H. Behaviour-based modelling of hexapod locomotion: Linking biology and technical application. Arthropod Struct. Dev. 2004, 33, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Mantziaris, C.; Bockemühl, T.; Holmes, P.; Borgmann, A.; Daun, S.; Büschges, A. Intra- and intersegmental influences among central pattern generating networks in the walking system of the stick insect. J. Neurophysiol. 2017, 118, 2296–2310. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H.; Riemenschneider, D.; Stammer, W. Control of body position of a stick insect standing on uneven surfaces. Biol. Cybern. 1989, 61, 71–77. [Google Scholar] [CrossRef]
- Cruse, H.; Schmitz, J.; Braun, U.; Schweins, A. Control of body height in a stick insect walking on a treadwheel. J. Exp. Biol. 1993, 181, 141–155. [Google Scholar]
- Murray, R.M.; Li, Z.; Sastry, S.S. A Mathematical Introduction to Robotic Manipulation; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Blumel, M.; Hooper, S.L.; Guschlbauer, C.; White, W.E.; Buschges, A. Determining all parameters necessary to build Hill-type muscle models from experiments on single muscles. Biol. Cybern. 2012, 106, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Guschlbauer, C.; Scharstein, H.; Büschges, A. The extensor tibiae muscle of the stick insect: Biomechanical properties of an insect walking leg muscle. J. Exp. Biol. 2007, 210, 1092–1108. [Google Scholar] [CrossRef] [PubMed]
- Blümel, M.; Guschlbauer, C.; Daun-Gruhn, S.; Hooper, S.L.; Büschges, A. Hill-type muscle model parameters determined from experiments on single muscles show large animal-to-animal variation. Biol. Cybern. 2012, 106, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Hooper, S.L.; Guschlbauer, C.; Blümel, M.; von Twickel, A.; Hobbs, K.H.; Thuma, J.B.; Büschges, A. Muscles: Non-Linear Transformers of Motor Neuron Activity; Springer Series in Computational Neuroscience; Springer: New York, NY, USA, 2016; pp. 163–194. [Google Scholar]
- Rassier, D.; MacIntosh, B.; Herzog, W. Length dependence of active force production in skeletal muscle. J. Appl. Physiol. 1999, 86, 1445–1457. [Google Scholar] [PubMed]
- Schroer, R.; Boggess, M.; Bachmann, R.; Quinn, R.; Ritzmann, R. Comparing cockroach and Whegs robot body motions. In Proceedings of the IEEE International Conference on Robotics and Automation, New Orleans, LA, USA, 26 April–1 May 2004; Volume 4, pp. 3288–3293. [Google Scholar]
- Hughes, G.M. The Co-ordination of Insect Movements. J. Exp. Biol. 1952, 29, 267–285. [Google Scholar]
- Szczecinski, N.S. Massively Distributed Neuromorphic Control for Legged Robots Modeled after Insect Stepping. Master’s Thesis, Case Western Reserve University, Cleveland, OH, USA, 2013. [Google Scholar]
- Rubeo, S.E. Control of Simulated Cockroach Using Synthethic Nervous Systems. Master’s Thesis, Case Western Reserve University, Cleveland, OH, USA, 2017. [Google Scholar]
- Rutter, B.L.; Lewinger, W.A.; Blumel, M.; Buschges, A.; Quinn, R.D. Simple Muscle Models Regularize Motion in a Robotic Leg with Neurally-Based Step Generation. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 630–635. [Google Scholar]
- Szczecinski, N.S.; Getsy, A.P.; Bosse, J.W.; Martin, J.P.; Ritzmann, R.E.; Quinn, R.D. MantisBot Uses Minimal Descending Commands to Pursue Prey as Observed in Tenodera Sinensis. In Biomimetic and Biohybrid Systems; Springer International Publishing: Cham, Switzerland, 2016; pp. 329–340. [Google Scholar]
- Bender, J.A.; Simpson, E.M.; Tietz, B.R.; Daltorio, K.A.; Quinn, R.D.; Ritzmann, R.E. Kinematic and behavioral evidence for a distinction between trotting and ambling gaits in the cockroach Blaberus discoidalis. J. Exp. Biol. 2011, 214, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Golowasch, J.; Goldman, M.S.; Abbott, L.F.; Marder, E. Failure of Averaging in the Construction of a Conductance-Based Neuron Model. J. Neurophysiol. 2002, 87, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Cruse, H. Which Parameters Control the Leg Movement of a Walking Insect? I. Velocity Control during the Stance Phase. J. Exp. Biol. 1985, 116, 343–355. [Google Scholar]
- Gruhn, M.; von Uckermann, G.; Westmark, S.; Wosnitza, A.; Buschges, A.; Borgmann, A. Control of Stepping Velocity in the Stick Insect Carausius morosus. J. Neurophysiol. 2009, 102, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Quinn, R.D. MantisBot Changes Stepping Speed by Entraining CPGs to Positive Velocity Feedback. In Biomimetic and Biohybrid Systems; Springer: Cham, Switzerland, 2017; pp. 440–452. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Actuated Joints | ||
|---|---|---|
| Front Leg | Middle Leg | Hind Leg |
| ThC2 | ThC2 | ThC2 |
| ThC1 | ThC1 | ThC1 |
| ThC3 | CTr | CTr |
| CTr | TrF | TrF |
| FTi | FTi | FTi |
| Influence Number | Direction of Influence | Effect of Influence |
|---|---|---|
| Influence 1 | Rostral | Inhibits the start of swing phase when the posterior leg is in its swing phase |
| Influence 2 | Rostral and contralateral | Excites the start of swing phase when the posterior leg enters its stance phase |
| Influence 3 | Caudal and contralateral | Excites the start of swing phase while the anterior leg is in its stance phase, increasing in strength over time |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubeo, S.; Szczecinski, N.; Quinn, R. A Synthetic Nervous System Controls a Simulated Cockroach. Appl. Sci. 2018, 8, 6. https://doi.org/10.3390/app8010006
Rubeo S, Szczecinski N, Quinn R. A Synthetic Nervous System Controls a Simulated Cockroach. Applied Sciences. 2018; 8(1):6. https://doi.org/10.3390/app8010006
Chicago/Turabian StyleRubeo, Scott, Nicholas Szczecinski, and Roger Quinn. 2018. "A Synthetic Nervous System Controls a Simulated Cockroach" Applied Sciences 8, no. 1: 6. https://doi.org/10.3390/app8010006
APA StyleRubeo, S., Szczecinski, N., & Quinn, R. (2018). A Synthetic Nervous System Controls a Simulated Cockroach. Applied Sciences, 8(1), 6. https://doi.org/10.3390/app8010006