Enhancement of Neuroprotective Effects of Spirulina platensis Extract from a High-Pressure Homogenization Process

Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of the Samples
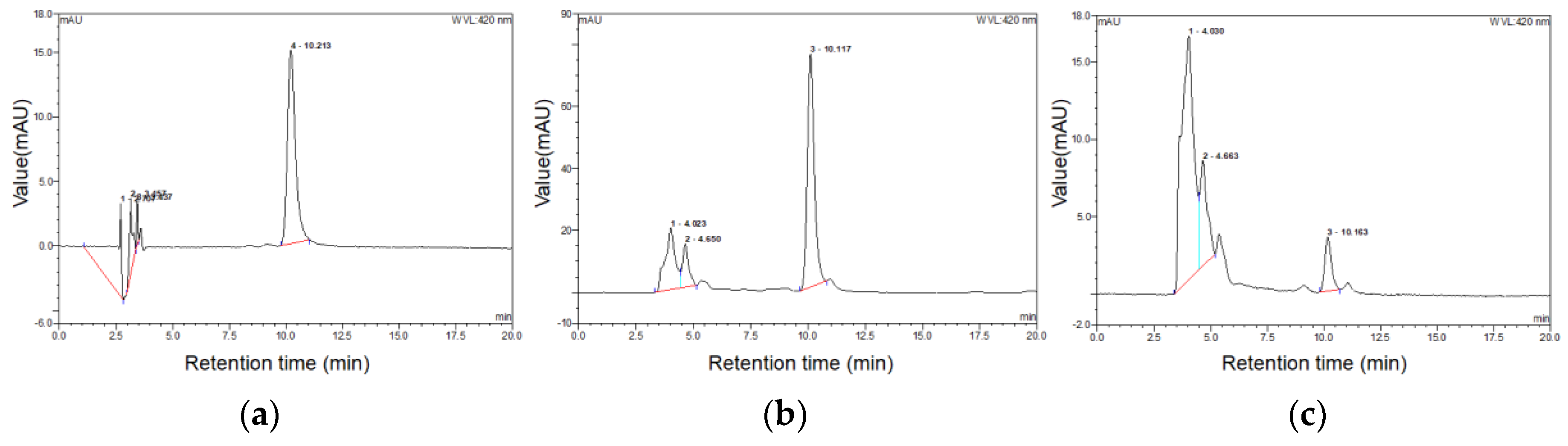
2.2. Determination of the Chlorophyll a Content of the Extracts
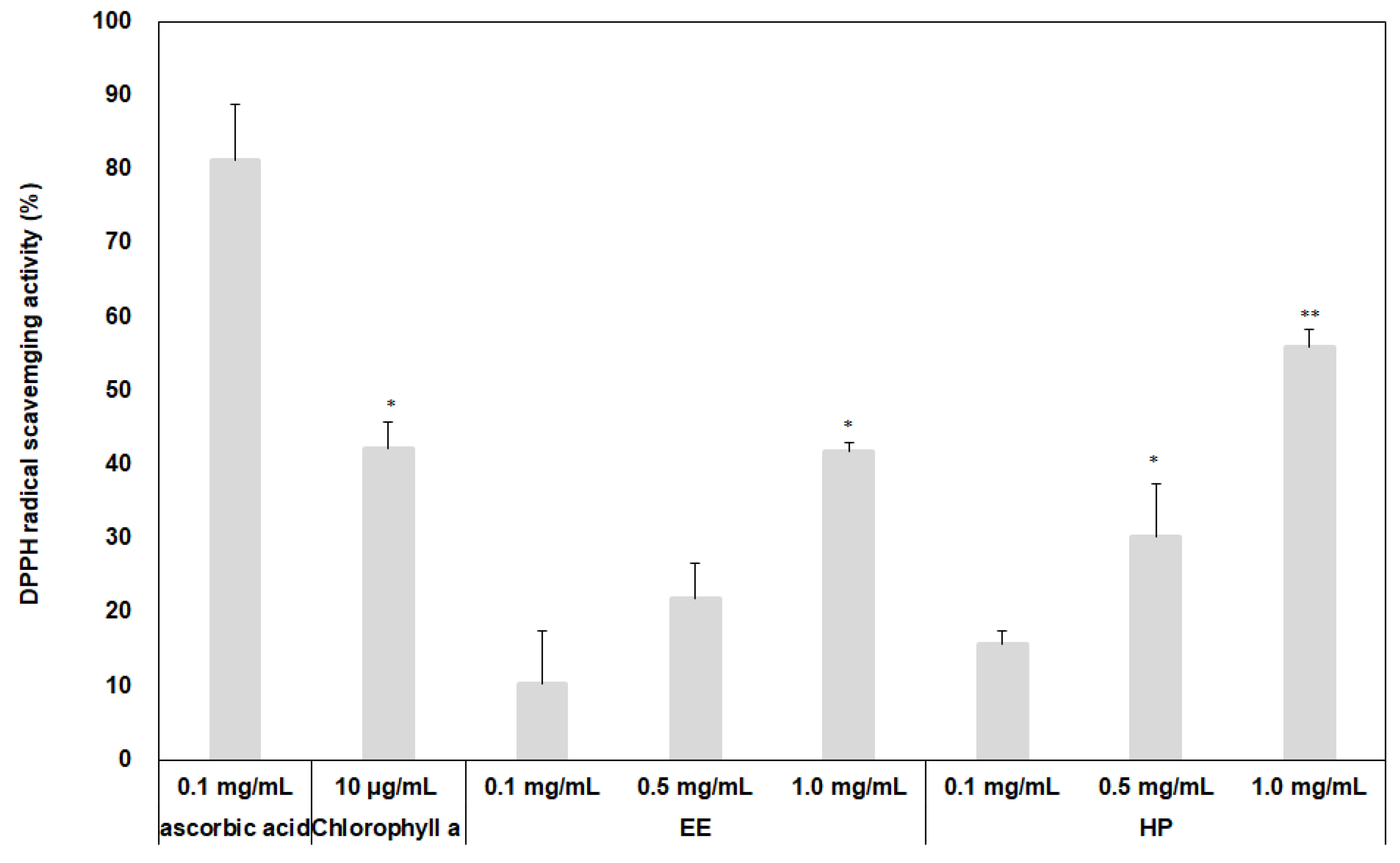
2.3. Measurement of Antioxidant Activity of the Extracts
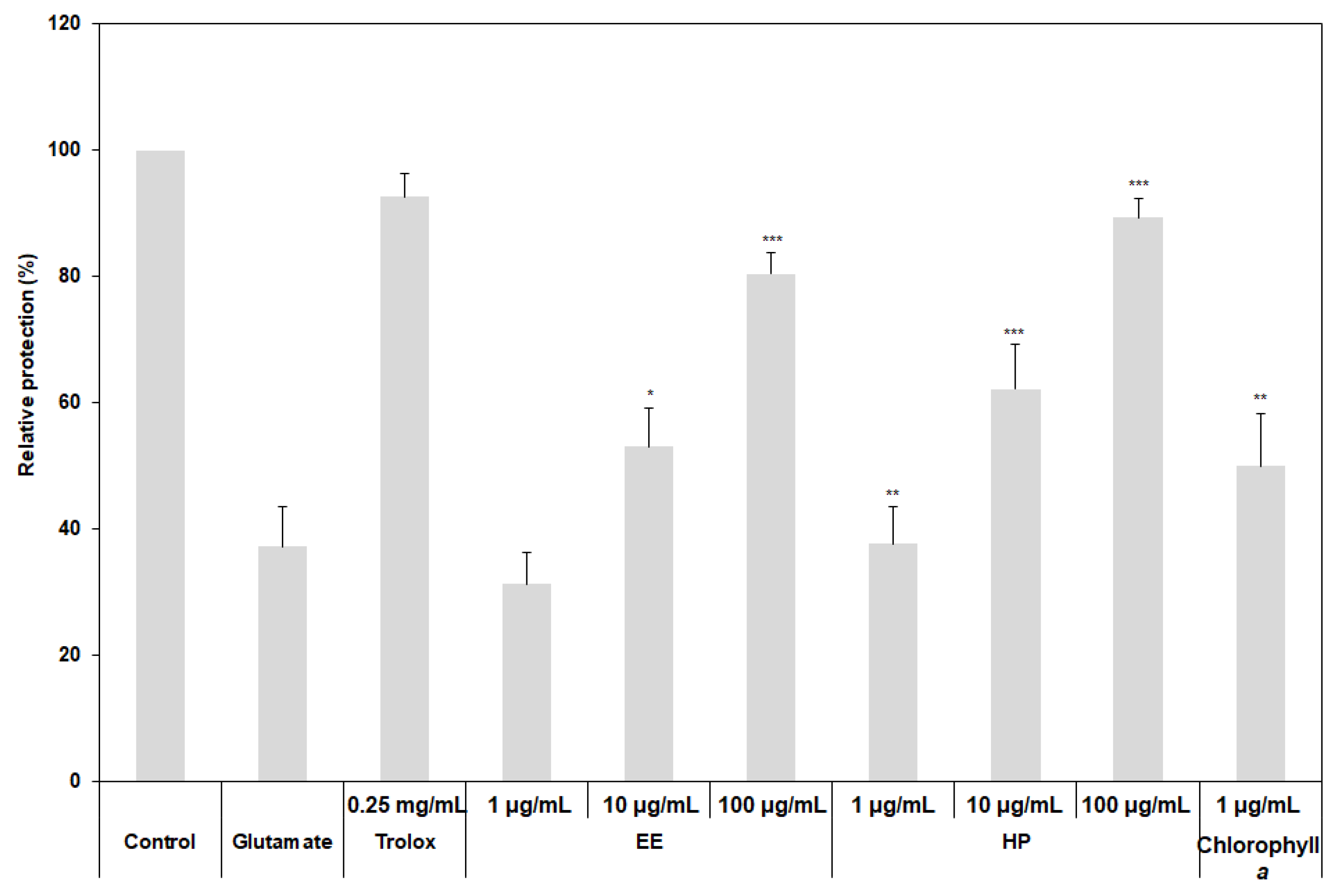
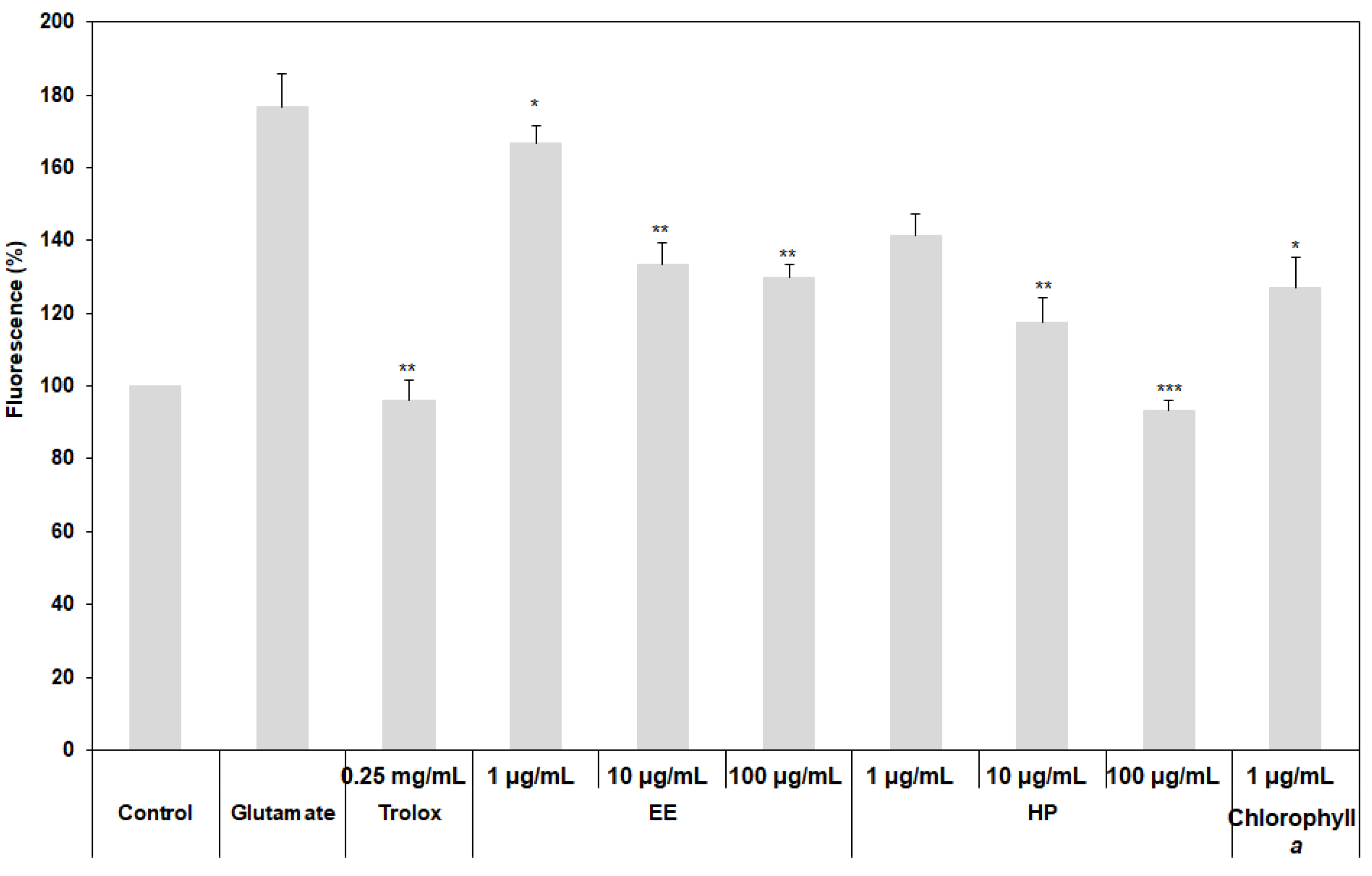
2.4. Measurement of Neuroprotective Activity of the Extracts
2.5. Statistical Analysis
3. Results
3.1. Comparison of Chlorophyll a Contents of Extracts and Morphology of Spirulina with and without High-Pressure Homogenization
3.2. Measurement of Antioxidant Effects of the Extracts Using Two Different Extraction Processes
3.3. Neuroprotective Activities of the Extracts from Different Extraction Processes
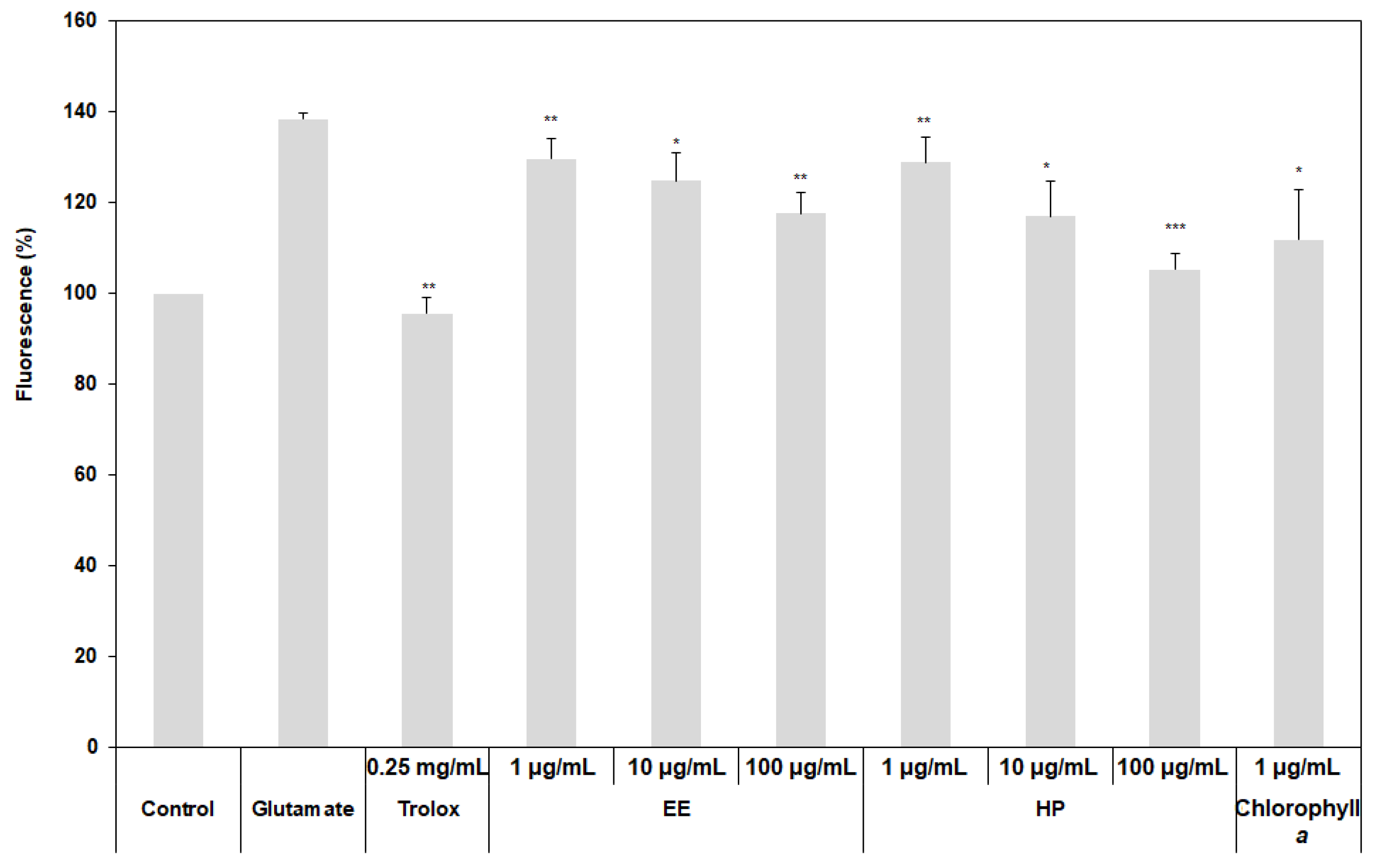
3.4. Inhibition of Oxidative Stress on HT22 Cells by the Extracts
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Khan, Z.; Bhadouria, P.; Bisen, P.S. Nutritional and therapeutic potential of Spirulina. Curr. Pharm. Biotechnol. 2005, 6, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Kay, R.A. Microalgae as food and supplement. Crit. Rev. Food Sci. Nutr. 1991, 30, 555–783. [Google Scholar] [CrossRef] [PubMed]
- Gad, A.S.; Khadrawy, Y.A.; El-Nekeety, A.A.; Mohamed, S.R.; Hassan, N.S.; Abdel-Wahhab, M.A. Antioxidant activity and hepatoprotective effects of whey protein and Spirulina in rats. Nutrition 2011, 27, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, S.H.; Kim, J.S.; Han, J.A.; Seo, H.J.; Lim, H.J.; Choi, S.Y. Studies on Simultaneous Determination of Chlorophyll a and b, Pheophorbide a, and β-Carotene in Chlorella and Spirulina Products. J. Food Hyg. Saf. 2005, 20, 141–146. [Google Scholar]
- Yamada, T.; Sakaguchi, K. Comparative studies on Chlorella cell walls: Induction of protoplast formation. Arch. Microbiol. 1982, 132, 10–13. [Google Scholar] [CrossRef]
- Silveira, S.T.; Burkert, J.F.M.; Costa, J.A.V.; Burkert, C.A.V.; Kalil, S.J. Optimization of phycocyanin extraction from Spirulina platensis using factorial design. Bioresour. Technol. 2007, 98, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Hosikian, A.; Lim, S.; Halim, R.; Danquah, M.K. Chlorophyll extraction from microalgae: A review on the process engineering aspects. Int. J. Chem. Eng. 2010, 2010, 1–11. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Shahbazizadeh, S.; Khosravi-Darani, K.; Mozafari, M.R. Spirulina paltensis: Food and Function. Curr. Nutr. Food Sci. 2013, 9, 189–193. [Google Scholar] [CrossRef]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell. Int. 2013, 13, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Danesi, E.D.G.; Rangel-Yagui, C.O.; Carvalho, J.C.M.; Sato, S. Effect of reducing the light intensity on the growth and production of chlorophyll by Spirulina platensis. Biomass Bioenergy 2004, 26, 329–335. [Google Scholar] [CrossRef]
- Bhat, S.R. Chlorophyll: The wonder pigment. Sci. Rep. 2005, 42, 29–32. [Google Scholar]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Furuki, T.; Maeda, S.; Imajo, S.; Hiroi, T.; Amaya, T.; Hirokawa, T.; Ito, K.; Nozawa, H. Rapid and selective extraction of phycocyanin from Spirulina platensis with ultrasonic cell disruption. J. Appl. Psychol. 2003, 15, 319–324. [Google Scholar] [CrossRef]
- Leonelli, C.; Mason, T.J. Microwave and ultrasonic processing: Now a realistic option for industry. Chem. Eng. Process 2010, 49, 885–900. [Google Scholar] [CrossRef]
- Fernandez-Avila, C.; Trujillo, A.J. Enhanced stability of emulsions treated by Ultra-High Pressure Homogenization for delivering conjugated linoleic acid in Caco-2 cells. Food. Hydrocoll. 2017, 71, 271–281. [Google Scholar] [CrossRef]
- Yap, B.H.J.; Dumsday, G.J.; Scales, P.J.; Martin, G.J.O. Energy evaluation of algal cell disruption by high pressure homogenization. Bioresour. Technol. 2015, 184, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Samarasinghe, N.; Fernando, S.; Lacey, R.; Faulkner, W.B. Algal cell rupture using high pressure homogenization as a prelude to oil extraction. Renew. Energy 2012, 48, 300–308. [Google Scholar] [CrossRef]
- Williams, S.T.; Davies, F.L. Use of a Scanning Electron Microscope for the Examination of Actinomycetes. Microbiology 1967, 48, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Petrović, S.M.; Savić, S.R.; Marković, D.Z.; Petronijević, Ž.B. In vitro studies of temperature and pH influence on chlorophyll degradation by horseradish peroxidase: Spectroscopic and HPLC studies. Hem. Ind. 2014, 68, 233–239. [Google Scholar] [CrossRef]
- Dietz, B.M.; Kang, Y.H.; Liu, G.; Eggler, A.L.; Yao, P.; Chadwick, L.R.; Pauli, G.F.; Farnsworth, N.R.; Mesecar, A.D.; Breemen, R.B.; et al. Xanthohumol isolated from Humulus lupulus inhibits menadione-induced DNA damage through induction of quinone reductase. Chem. Res. Toxicol. 2005, 18, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Elmlinger, M.W.; Kriebel, M.; Ziegler, D. Neuroprotective and Anti-Oxidative Effects of the Hemodialysate Actovegin on Primary Rat Neurons in Vitro. Neuromol. Med. 2011, 13, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Weon, J.B.; Ma, C.J. Neuroprotective activity of phytosterols isolated from Artemisia apiacea. Korean J. Pharmacogn. 2014, 45, 214–219. [Google Scholar]
- Rehni, A.K.; Pantlya, H.S.; Shri, R.; Singh, M. Effect of chlorophyll and aqueous extracts of Bacopa monniera and Valeriana wallichii on ischaemia and reperfusion-induced cerebral injury in mice. Indian J. Exp. Biol. 2007, 45, 764–769. [Google Scholar] [PubMed]
- Chernomorsky, S.A.; Segelman, A.B. Biological activities of chlorophyll derivatives. N. J. Med. 1988, 85, 669–673. [Google Scholar]
- Rangel-Yagui, C.O.; Danesi, E.D.G.; Carvalho, J.C.M.; Sato, S. Chlorophyll production from Spirulina platensis: Cultivation with urea addition by fed-batch process. Bioresour. Technol. 2004, 92, 133–141. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Pignolet, O.; Silvestre, F.; Vaca-Garcia, C.; Pontalier, P.Y. Influence of microalgae cell wall characteristics on protein extractability and determination of nitrogen-to-protein conversion factors. J. Appl. Phycol. 2013, 25, 523–529. [Google Scholar] [CrossRef]
- Ahmed, I.; John, A.; Vijayasarathy, C.; Robin, M.A.; Raza, H. Differential modulation of growth and glutathione metabolism in cultured rat astrocytes by 4-hydroxynonenal and green tea polyphenol, epigallocatechin-3-gallate. Neurotoxicology 2002, 23, 289–300. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Jones, D.P. Glutathione depletion enforces the mitochondrial permeability transition and causes cell death in Bcl-2 overexpressing HL60 cells. FASEB J. 2002, 16, 1263–1265. [Google Scholar] [CrossRef] [PubMed]
- Weon, J.B.; Lee, B.; Yun, B.R.; Lee, J.; Ma, J.Y.; Ma, C.J. Neuroprotective and cognitive enhancing activity of the fermented Bozhougyiqi-Tang. Pharmacogn. Mag. 2014, 10, 249–255. [Google Scholar] [CrossRef]
- Kim, E.S.; Weon, J.B.; Yun, B.R.; Lee, J.; Eom, M.R.; Oh, K.H.; Ma, C.J. Cognitive Enhancing and Neuroprotective Effect of the Embryo of the Nelumbo nucifera Seed. Evid. Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction Process | Chlorophyll a Contents (mg/g) |
|---|---|
| EE 1 | 5.27 ± 1.73 |
| HP-1 2 | 4.24 ± 2.61 * |
| HP-2 3 | 9.85 ± 1.41 *** |
| HP-3 4 | 6.32 ± 3.09 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.Y.; Lee, H.Y. Enhancement of Neuroprotective Effects of Spirulina platensis Extract from a High-Pressure Homogenization Process. Appl. Sci. 2018, 8, 634. https://doi.org/10.3390/app8040634
Choi WY, Lee HY. Enhancement of Neuroprotective Effects of Spirulina platensis Extract from a High-Pressure Homogenization Process. Applied Sciences. 2018; 8(4):634. https://doi.org/10.3390/app8040634
Chicago/Turabian StyleChoi, Woon Yong, and Hyeon Yong Lee. 2018. "Enhancement of Neuroprotective Effects of Spirulina platensis Extract from a High-Pressure Homogenization Process" Applied Sciences 8, no. 4: 634. https://doi.org/10.3390/app8040634
APA StyleChoi, W. Y., & Lee, H. Y. (2018). Enhancement of Neuroprotective Effects of Spirulina platensis Extract from a High-Pressure Homogenization Process. Applied Sciences, 8(4), 634. https://doi.org/10.3390/app8040634