Changes in Serotonin Modulation of Glutamate Currents in Pyramidal Offspring Cells of Rats Treated With 5-MT during Gestation

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
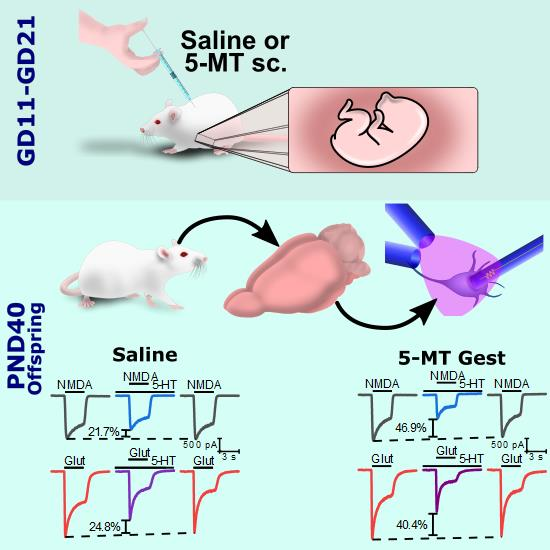
2.1. Animals
2.2. Reagent
2.3. Acute-Dissociation Procedure
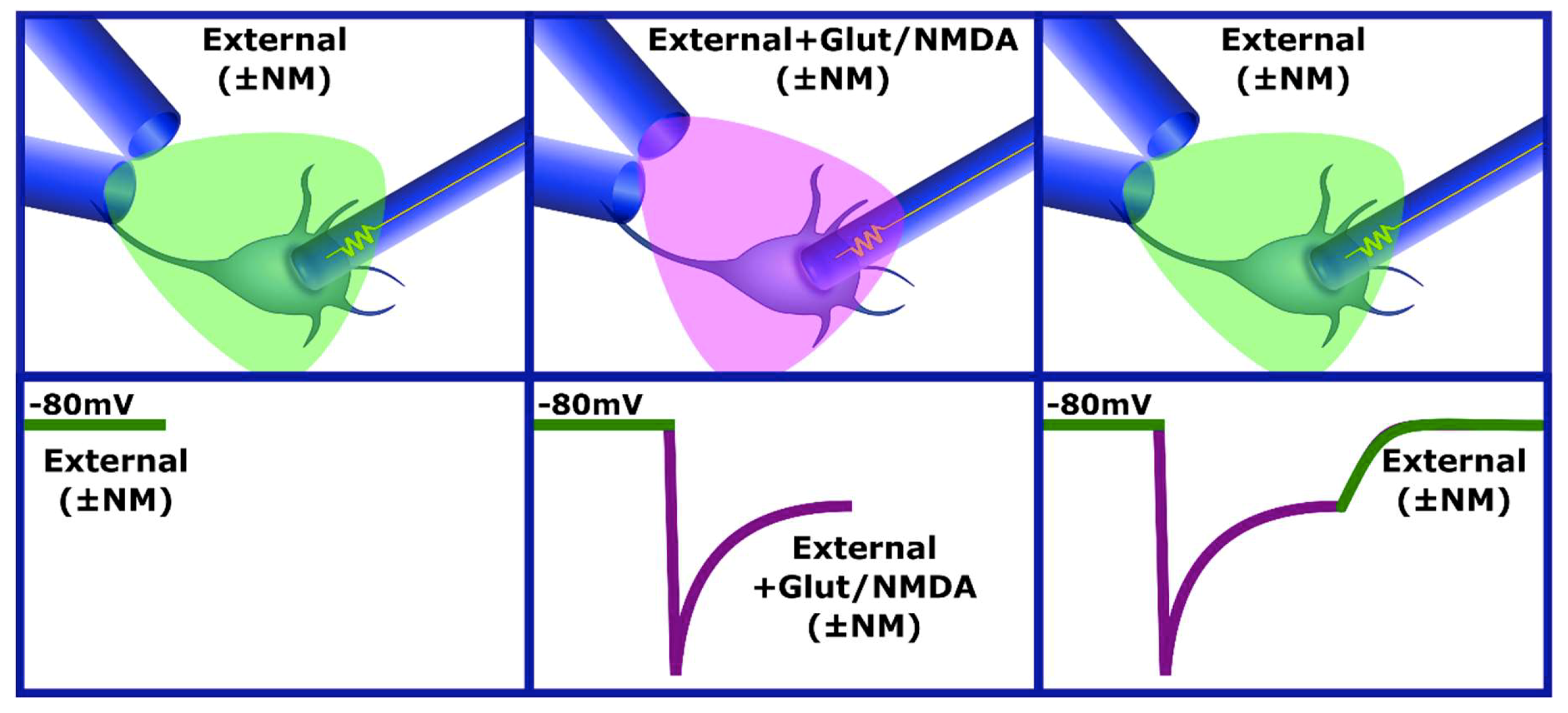
2.4. Whole-Cell Patch-Clamp Technique
2.5. Drug Application
2.6. Statistical Analysis
3. Results
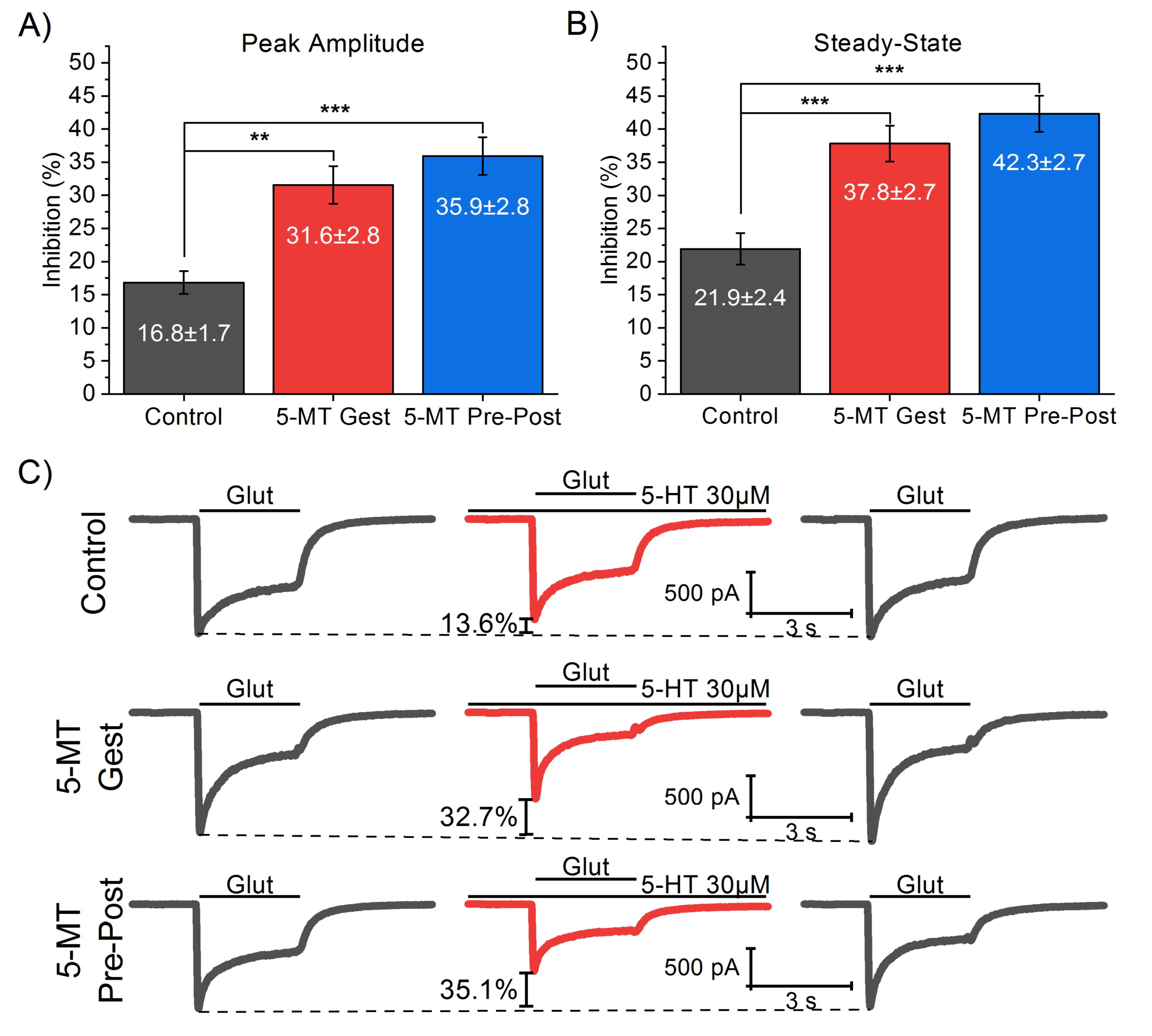
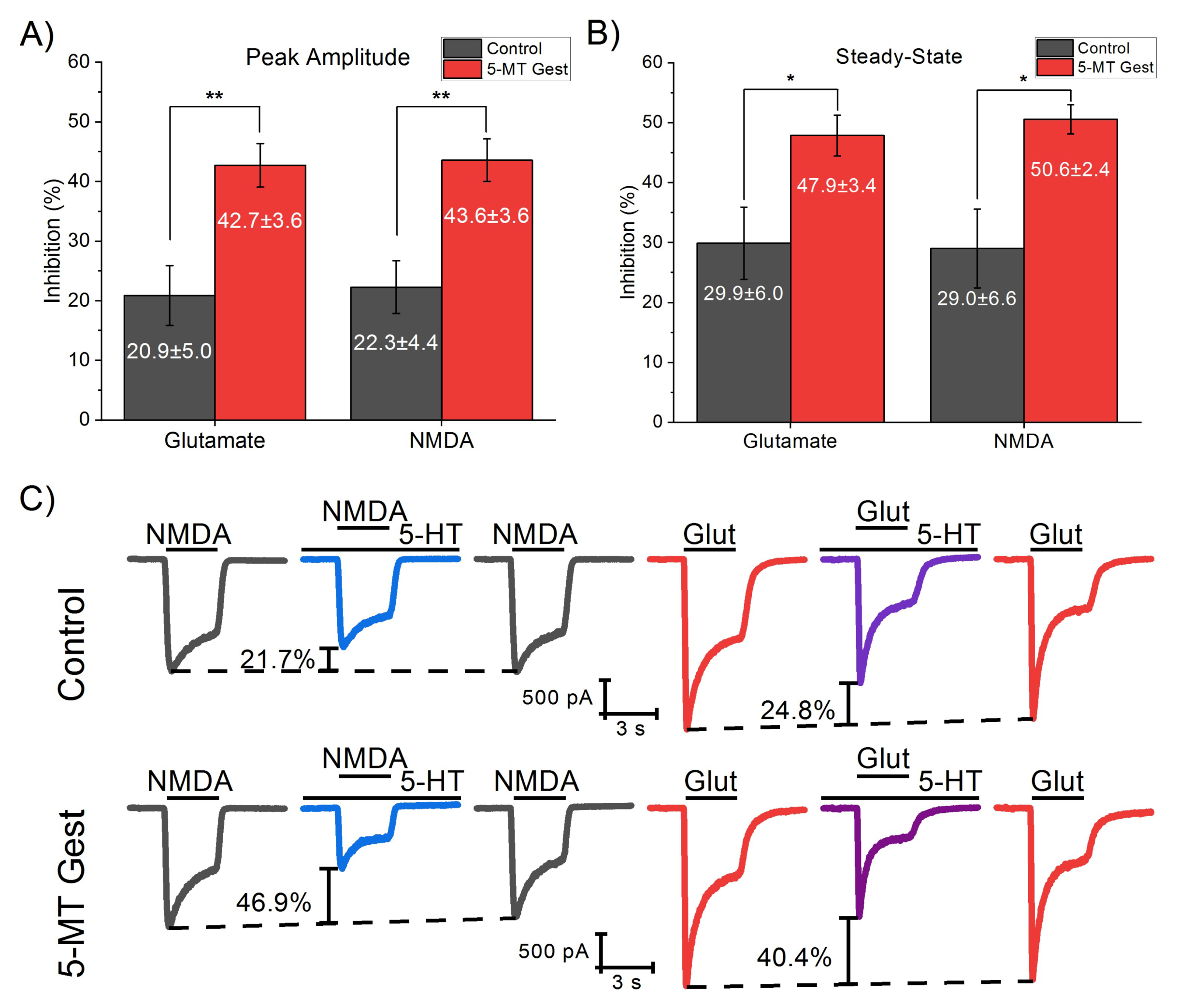
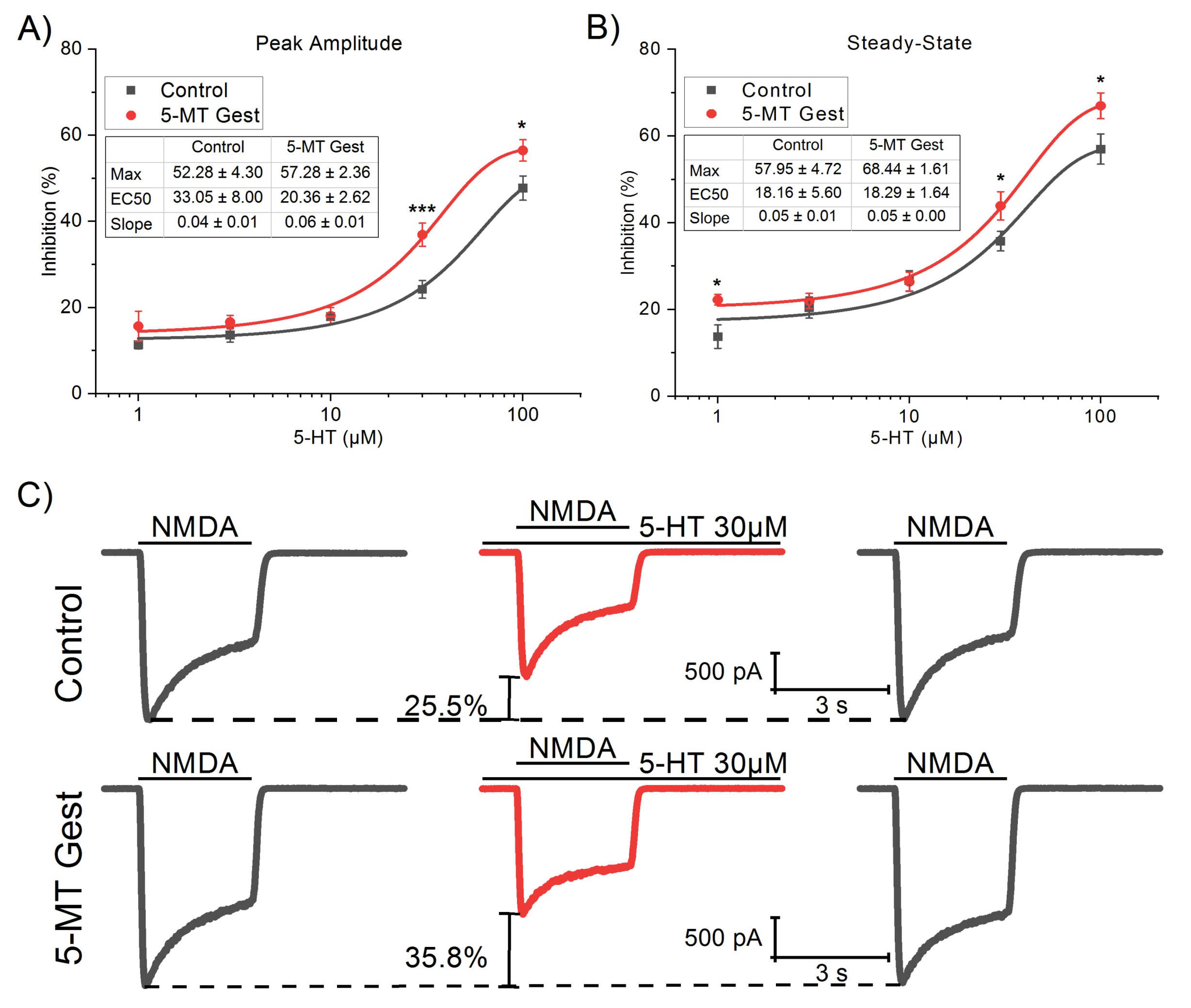
3.1. 5-MT Treatment from GD11 to GD21 Increases the 5-HT Effect on IGlut
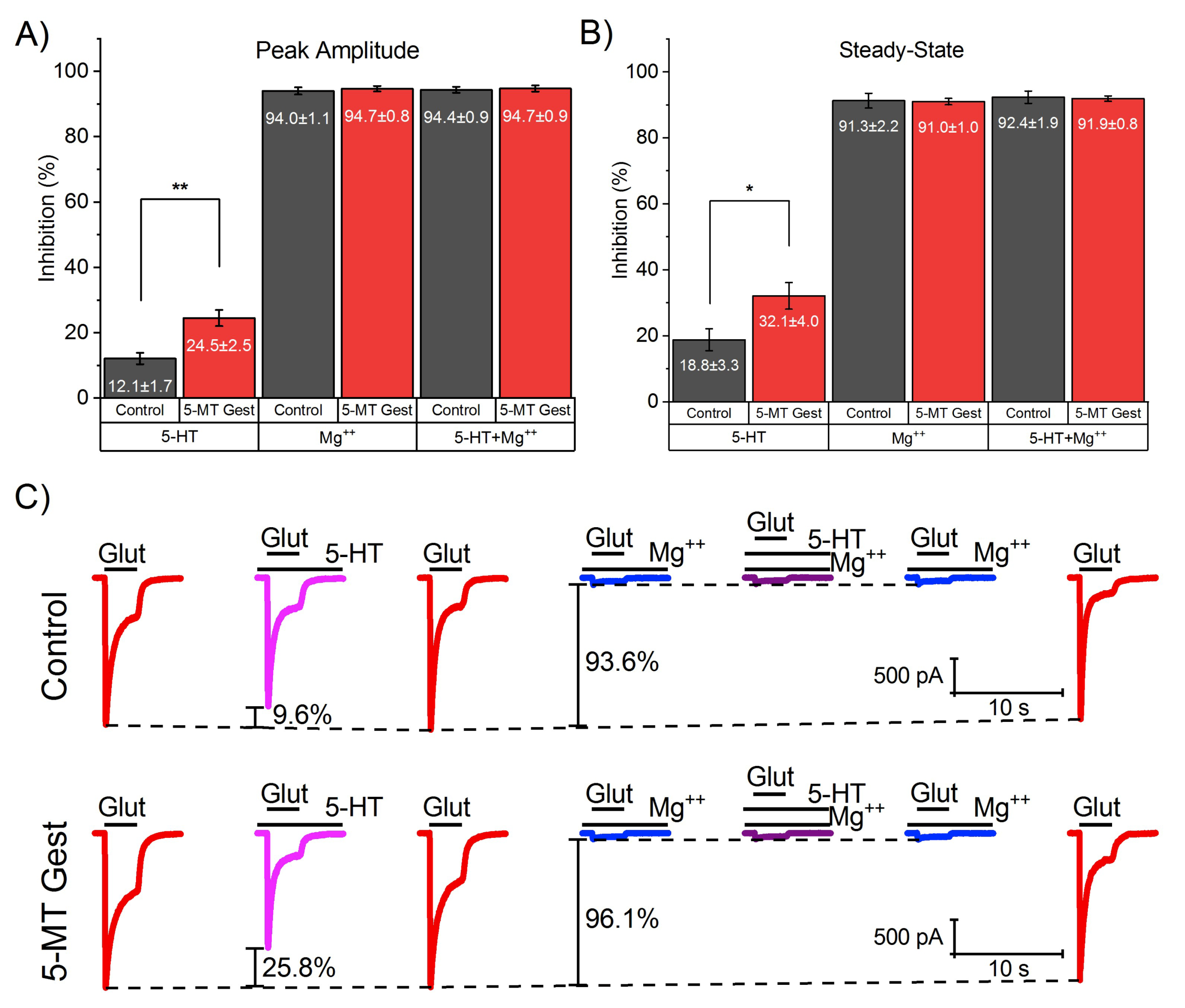
3.2. 5-HT Reduces the NMDA Receptor-Mediated Component of IGlut
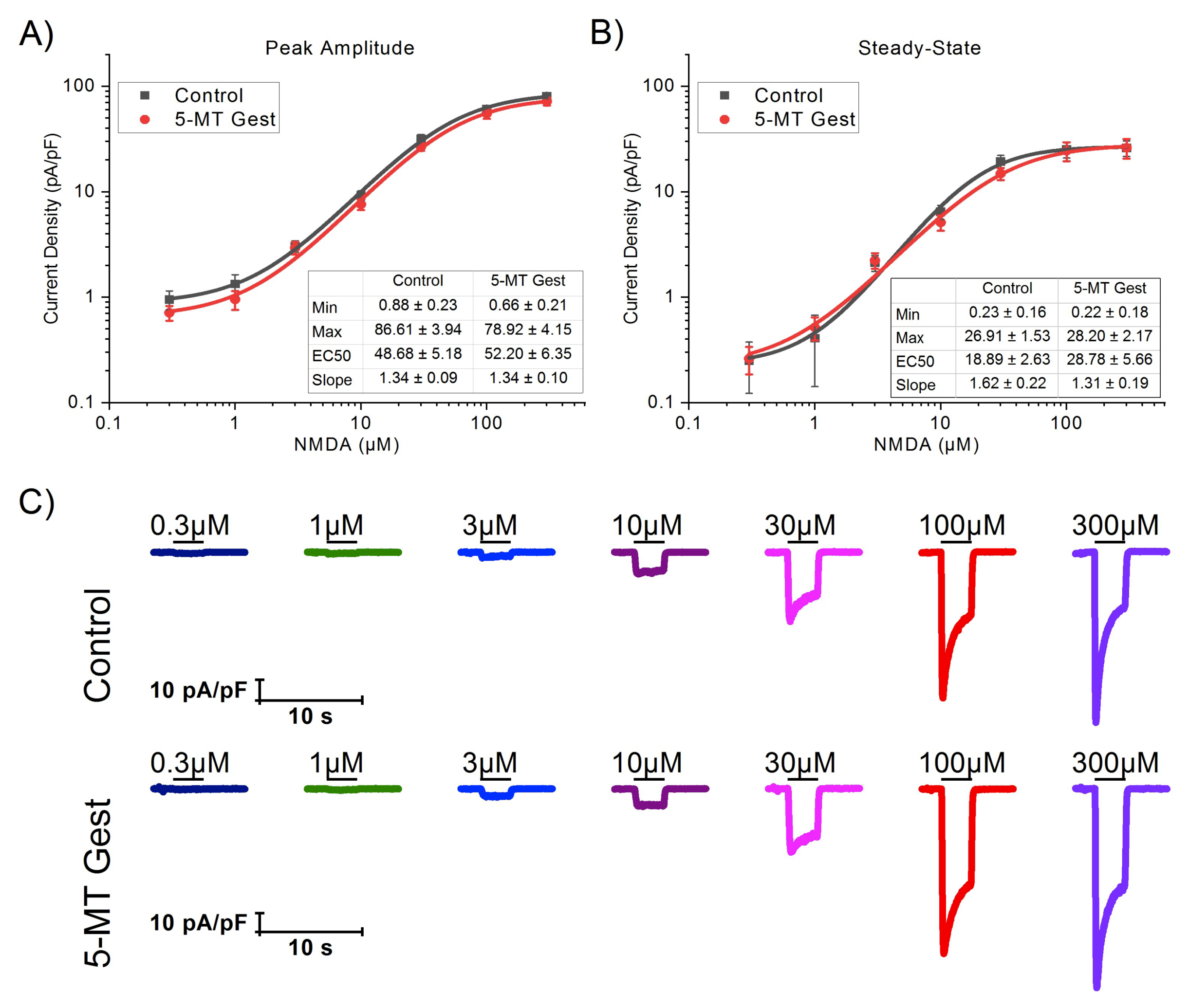
3.3. 5-MT Treatment Increases the Modulatory Effect of 5-HT on INMDA
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beyond Neurotransmission: Neuromodulation and Its Importance for Information Processing, 1st ed.; Katz, P.S. (Ed.) Oxford University Press: New York, NY, USA, 1999; ISBN 978-0-19-852424-3. [Google Scholar]
- Kupfermann, I. Modulatory actions of neurotransmitters. Annu. Rev. Neurosci. 1979, 2, 447–465. [Google Scholar] [CrossRef]
- Levitan, I.B. Modulation of ION channels in neurons and other cells. Annu. Rev. Neurosci. 1988, 11, 119–136. [Google Scholar] [CrossRef]
- Lopez, H.S.; Brown, A.M. Neuromodulation. Curr. Opin. Neurobiol. 1992, 2, 317–322. [Google Scholar] [CrossRef]
- Marder, E.; Thirumalai, V. Cellular, synaptic and network effects of neuromodulation. Neural Netw. 2002, 15, 479–493. [Google Scholar] [CrossRef]
- Nadim, F.; Bucher, D. Neuromodulation of neurons and synapses. Curr. Opin. Neurobiol. 2014, 29, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Zoli, M.; Agnati, L.F. Wiring and volume transmission in the central nervous system: The concept of closed and open synapses. Prog. Neurobiol. 1996, 49, 363–380. [Google Scholar] [CrossRef]
- Cassel, J.-C. Experimental studies on the role(s) of serotonin in learning and memory functions. In Handbook of Behavioral Neuroscience; Müller, C.P., Jacobs, B.L., Eds.; Handbook of the Behavioral Neurobiology of Serotonin; Elsevier: Amsterdam, The Netherlands, 2010; Volume 21, pp. 429–447. [Google Scholar]
- Duman, E.A.; Canli, T. Social behavior and serotonin. In Handbook of Behavioral Neuroscience; Jacobs, C.P.M.B.L., Ed.; Handbook of the Behavioral Neurobiology of Serotonin; Elsevier: Amsterdam, The Netherlands, 2010; Volume 21, pp. 449–456. [Google Scholar]
- Hensler, J.G. Serotonin in mood and emotion. In Handbook of Behavioral Neuroscience; Jacobs, C.P.M.B.L., Ed.; Handbook of the Behavioral Neurobiology of Serotonin; Elsevier: Amsterdam, The Netherlands, 2010; Volume 21, pp. 367–378. [Google Scholar]
- Aghajanian, G.K.; Marek, G.J. Serotonin model of schizophrenia: Emerging role of glutamate mechanisms. Brain Res. Brain Res. Rev. 2000, 31, 302–312. [Google Scholar] [CrossRef]
- Albert, P.R.; Vahid-Ansari, F.; Luckhart, C. Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: Pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Front. Behav. Neurosci. 2014, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Eggers, A.E. A serotonin hypothesis of schizophrenia. Med. Hypotheses 2013, 80, 791–794. [Google Scholar] [CrossRef]
- Lin, S.-H.; Lee, L.-T.; Yang, Y.K. Serotonin and mental disorders: A concise review on molecular neuroimaging evidence. Clin. Psychopharmacol. Neurosci. 2014, 12, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y. Neuropaediatric and neuroarchaeology: Understanding development to correct brain disorders. Acta Paediatr. 2013, 102, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Frederick, A.L.; Stanwood, G.D. Drugs, biogenic amine targets and the developing brain. DNE 2009, 31, 7–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herlenius, E.; Lagercrantz, H. Development of neurotransmitter systems during critical periods. Exp. Neurol. 2004, 190, 8–21. [Google Scholar] [CrossRef]
- Ruediger, T.; Bolz, J. Neurotransmitters and the development of neuronal circuits. In Axon Growth and Guidance; Advances in Experimental Medicine and, Biology; Bagnard, D., Ed.; Springer: New York, NY, USA, 2007; pp. 104–114. ISBN 978-0-387-76715-4. [Google Scholar]
- Sundström, E.; Kölare, S.; Souverbie, F.; Samuelsson, E.B.; Pschera, H.; Lunell, N.O.; Seiger, A. Neurochemical differentiation of human bulbospinal monoaminergic neurons during the first trimester. Brain Res. Dev. Brain Res. 1993, 75, 1–12. [Google Scholar] [CrossRef]
- Lauder, J.M. Ontogeny of the serotonergic system in the rat: Serotonin as a developmental signal. Ann. N. Y. Acad. Sci. 1990, 600, 297–313, discussion 314. [Google Scholar] [CrossRef]
- Rubenstein, J.L. Development of serotonergic neurons and their projections. Biol. Psychiatry 1998, 44, 145–150. [Google Scholar] [CrossRef]
- Buznikov, G.A.; Lambert, W.H.; Lauder, J.M. Serotonin and serotonin-like substances as regulators of early embryogenesis and morphogenesis. Cell Tissue Res. 2001, 305, 177–186. [Google Scholar] [CrossRef]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Booij, L.; Tremblay, R.E.; Szyf, M.; Benkelfat, C. Genetic and early environmental influences on the serotonin system: Consequences for brain development and risk for psychopathology. J. Psychiatry Neurosci. 2015, 40, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Brummelte, S.; Mc Glanaghy, E.; Bonnin, A.; Oberlander, T.F. Developmental changes in serotonin signaling: Implications for early brain function, behavior and adaptation. Neuroscience 2017, 342, 212–231. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Oberlander, T.F. Fetal serotonin signaling: Setting pathways for early childhood development and behavior. J. Adolesc. Health 2012, 51, S9–S16. [Google Scholar] [CrossRef]
- Whitaker-Azmitia, P.M. Serotonin and brain development: Role in human developmental diseases. Brain Res. Bull. 2001, 56, 479–485. [Google Scholar] [CrossRef]
- Shah, R.; Courtiol, E.; Castellanos, F.X.; Teixeira, C.M. Abnormal serotonin levels during perinatal development lead to behavioral deficits in adulthood. Front. Behav. Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suri, D.; Teixeira, C.M.; Cagliostro, M.K.C.; Mahadevia, D.; Ansorge, M.S. Monoamine-sensitive developmental periods impacting adult emotional and cognitive behaviors. Neuropsychopharmacology 2015, 40, 88–112. [Google Scholar] [CrossRef] [Green Version]
- Vitalis, T.; Cases, O.; Passemard, S.; Callebert, J.; Parnavelas, J.G. Embryonic depletion of serotonin affects cortical development. Eur. J. Neurosci. 2007, 26, 331–344. [Google Scholar] [CrossRef]
- González-Burgos, I.; Angel-Meza, A.R.D.; Barajas-López, G.; Feria-Velasco, A. Tryptophan restriction causes long-term plastic changes in corticofrontal pyramidal neurons. Int. J. Dev. Neurosci. 1996, 14, 673–679. [Google Scholar] [CrossRef]
- Dooley, A.E.; Pappas, I.S.; Parnavelas, J.G. Serotonin promotes the survival of cortical glutamatergic neuronsin vitro. Exp. Neurol. 1997, 148, 205–214. [Google Scholar] [CrossRef]
- Riccio, O.; Potter, G.; Walzer, C.; Vallet, P.; Szabó, G.; Vutskits, L.; Kiss, J.Z.; Dayer, A.G. Excess of serotonin affects embryonic interneuron migration through activation of the serotonin receptor 6. Mol. Psychiatry 2009, 14, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Riccio, O.; Jacobshagen, M.; Golding, B.; Vutskits, L.; Jabaudon, D.; Hornung, J.P.; Dayer, A.G. Excess of serotonin affects neocortical pyramidal neuron migration. Transl. Psychiatry 2011, 1, e47. [Google Scholar] [CrossRef]
- Ansorge, M.S.; Morelli, E.; Gingrich, J.A. Inhibition of serotonin but not norepinephrine transport during development produces delayed, persistent perturbations of emotional behaviors in mice. J. Neurosci. 2008, 28, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Sprowles, J.L.N.; Hufgard, J.R.; Gutierrez, A.; Bailey, R.A.; Jablonski, S.A.; Williams, M.T.; Vorhees, C.V. Differential effects of perinatal exposure to antidepressants on learning and memory, acoustic startle, anxiety, and open-field activity in Sprague-Dawley rats. Int. J. Dev. Neurosci. 2017, 61, 92–111. [Google Scholar] [CrossRef] [PubMed]
- Lauder, J.M.; Liu, J.; Grayson, D.R. In utero exposure to serotonergic drugs alters neonatal expression of 5-HT(1A) receptor transcripts: A quantitative RT-PCR study. Int. J. Dev. Neurosci. 2000, 18, 171–176. [Google Scholar] [CrossRef]
- Madden, A.M.K.; Zup, S.L. Effects of developmental hyperserotonemia on juvenile play behavior, oxytocin and serotonin receptor expression in the hypothalamus are age and sex dependent. Physiol. Behav. 2014, 128, 260–269. [Google Scholar] [CrossRef]
- Blažević, S.; Hranilović, D. Expression of 5HT-related genes after perinatal treatment with 5HT agonists. Transl.Neurosci. 2013, 4, 165–171. [Google Scholar] [CrossRef]
- Cases, O.; Vitalis, T.; Seif, I.; Maeyer, E.D.; Sotelo, C.; Gaspar, P. Lack of barrels in the somatosensory cortex of monoamine oxidase A–deficient mice: Role of a serotonin excess during the critical period. Neuron 1996, 16, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Vitalis, T.; Cases, O.; Callebert, J.; Launay, J.-M.; Price, D.J.; Seif, I.; Gaspar, P. Effects of monoamine oxidase A inhibition on barrel formation in the mouse somatosensory cortex: Determination of a sensitive developmental period. J. Comp. Neurol. 1998, 393, 169–184. [Google Scholar] [CrossRef]
- Janušonis, S.; Gluncic, V.; Rakic, P. Early serotonergic projections to Cajal-Retzius cells: Relevance for cortical development. J. Neurosci. 2004, 24, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Rebello, T.J.; Yu, Q.; Goodfellow, N.M.; Cagliostro, M.K.C.; Teissier, A.; Morelli, E.; Demireva, E.Y.; Chemiakine, A.; Rosoklija, G.B.; Dwork, A.J.; et al. Postnatal day 2 to 11 constitutes a 5-HT-sensitive period impacting adult mPFC function. J. Neurosci. 2014, 34, 12379–12393. [Google Scholar] [CrossRef]
- Fuster, J.M. The Prefrontal Cortex, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 978-0-12-373644-4. [Google Scholar]
- Catts, V.S.; Fung, S.J.; Long, L.E.; Joshi, D.; Vercammen, A.; Allen, K.M.; Fillman, S.G.; Moore, L.; Rothmond, D.; Sinclair, D.; et al. Rethinking schizophrenia in the context of normal neurodevelopment. Front. Cell. Neurosci. 2013, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, A.; Conn, P.J. The hippocampo-prefrontal pathway: A possible therapeutic target for negative and cognitive symptoms of schizophrenia. Futur. Neurol. 2015, 10, 115–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deserno, L.; Sterzer, P.; Wüstenberg, T.; Heinz, A.; Schlagenhauf, F. Reduced prefrontal-parietal effective connectivity and working memory deficits in schizophrenia. J. Neurosci. 2012, 32, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.P.; Witteveen, J.S.; Middelman, A.; van Hulten, J.A.; Martens, G.J.M.; Homberg, J.R.; Kolk, S.M. Perturbed developmental serotonin signaling affects prefrontal catecholaminergic innervation and cortical integrity. Mol. Neurobiol. 2019, 56, 1405–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumback, A.C.; Ellwood, I.T.; Kjaerby, C.; Iafrati, J.; Robinson, S.; Lee, A.T.; Patel, T.; Nagaraj, S.; Davatolhagh, F.; Sohal, V.S. Identifying specific prefrontal neurons that contribute to autism-associated abnormalities in physiology and social behavior. Mol. Psychiatry 2018, 23, 2078–2089. [Google Scholar] [CrossRef]
- Willsey, A.J.; Sanders, S.J.; Li, M.; Dong, S.; Tebbenkamp, A.T.; Muhle, R.A.; Reilly, S.K.; Lin, L.; Fertuzinhos, S.; Miller, J.A.; et al. Coexpression networks implicate human midfetal deep cortical Projection neurons in the pathogenesis of autism. Cell 2013, 155, 997–1007. [Google Scholar] [CrossRef] [Green Version]
- Goeden, N.; Velasquez, J.; Arnold, K.A.; Chan, Y.; Lund, B.T.; Anderson, G.M.; Bonnin, A. Maternal inflammation disrupts fetal neurodevelopment via increased placental output of serotonin to the fetal brain. J. Neurosci. 2016, 36, 6041–6049. [Google Scholar] [CrossRef] [Green Version]
- Peters, D.A. Prenatal stress increases the behavioral response to serotonin agonists and alters open field behavior in the rat. Pharmacol. Biochem. Behav. 1986, 25, 873–877. [Google Scholar] [CrossRef]
- Peters, D.A. Maternal stress increases fetal brain and neonatal cerebral cortex 5-hydroxytryptamine synthesis in rats: A possible mechanism by which stress influences brain development. Pharmacol. Biochem. Behav. 1990, 35, 943–947. [Google Scholar] [CrossRef]
- St-Pierre, J.; Laurent, L.; King, S.; Vaillancourt, C. Effects of prenatal maternal stress on serotonin and fetal development. Placenta 2016, 48, S66–S71. [Google Scholar] [CrossRef]
- Lattimore, K.A.; Donn, S.M.; Kaciroti, N.; Kemper, A.R.; Neal, C.R.; Vazquez, D.M. Selective Serotonin Reuptake Inhibitor (SSRI) use during pregnancy and effects on the fetus and newborn: A meta-analysis. J. Perinatol. 2005, 25, 595–604. [Google Scholar] [CrossRef]
- Olivier, J.D.; Åkerud, H.; Kaihola, H.; Pawluski, J.L.; Skalkidou, A.; Högberg, U.; Sundström Poromaa, I. The effects of maternal depression and maternal selective serotonin reuptake inhibitor exposure on offspring. Front. Cell. Neurosci. 2013, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasquez, J.C.; Goeden, N.; Bonnin, A. Placental serotonin: Implications for the developmental effects of SSRIs and maternal depression. Front. Cell. Neurosci. 2013, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker-Azmitia, P.M. Behavioral and cellular consequences of increasing serotonergic activity during brain development: A role in autism? Int. J. Dev. Neurosci. 2005, 23, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bargas, J.; Howe, A.; Eberwine, J.; Cao, Y.; Surmeier, D.J. Cellular and molecular characterization of Ca2+ currents in acutely isolated, adult rat neostriatal neurons. J. Neurosci. 1994, 14, 6667–6686. [Google Scholar] [CrossRef] [PubMed]
- Flores-Hernández, J.; Cepeda, C.; Hernández-Echeagaray, E.; Calvert, C.R.; Jokel, E.S.; Fienberg, A.A.; Greengard, P.; Levine, M.S. Dopamine enhancement of NMDA currents in dissociated medium-sized striatal neurons: Role of D1 receptors and DARPP-32. J. Neurophysiol. 2002, 88, 3010–3020. [Google Scholar] [CrossRef] [PubMed]
- Flores-Hernández, J.; Hernandez, S.; Snyder, G.L.; Yan, Z.; Fienberg, A.A.; Moss, S.J.; Greengard, P.; Surmeier, D.J. D(1) dopamine receptor activation reduces GABA(A) receptor currents in neostriatal neurons through a PKA/DARPP-32/PP1 signaling cascade. J. Neurophysiol. 2000, 83, 2996–3004. [Google Scholar] [CrossRef] [Green Version]
- Santana, N.; Artigas, F. Laminar and cellular distribution of monoamine receptors in rat medial prefrontal cortex. Front. Neuroanat. 2017, 11, 87. [Google Scholar] [CrossRef]
- Parsons, C.G.; Danysz, W.; Hesselink, M.; Hartmann, S.; Lorenz, B.; Wollenburg, C.; Quack, G. Modulation of NMDA receptors by glycine — introduction to some basic aspects and recent developments. Amino Acids 1998, 14, 207–216. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Shemer, A.V.; Azmitia, E.C.; Whitaker-Azmitia, P.M. Dose-related effects of prenatal 5-methoxytryptamine (5-MT) on development of serotonin terminal density and behavior. Brain Res. Dev. Brain Res. 1991, 59, 59–63. [Google Scholar] [CrossRef]
- Whitaker-Azmitia, P.M.; Azmitia, E.C. Autoregulation of fetal serotonergic neuronal development: Role of high affinity serotonin receptors. Neurosci. Lett. 1986, 67, 307–312. [Google Scholar]
- Martin, M.M.; Liu, Y.; Wang, Z. Developmental exposure to a serotonin agonist produces subsequent behavioral and neurochemical changes in the adult male prairie vole. Physiol. Behav. 2012, 105, 529–535. [Google Scholar]
- Kahne, D.; Tudorica, A.; Borella, A.; Shapiro, L.; Johnstone, F.; Huang, W.; Whitaker-Azmitia, P.M. Behavioral and magnetic resonance spectroscopic studies in the rat hyperserotonemic model of autism. Physiol. Behav. 2002, 75, 403–410. [Google Scholar]
- McNamara, I.M.; Borella, A.W.; Bialowas, L.A.; Whitaker-Azmitia, P.M. Further studies in the developmental hyperserotonemia model (DHS) of autism: Social, behavioral and peptide changes. Brain Res. 2008, 1189, 203–214. [Google Scholar]
- Carlsson, M.L. Hypothesis: Is infantile autism a hypoglutamatergic disorder? Relevance of glutamate – serotonin interactions for pharmacotherapy. J. Neural Transm. 1998, 105, 525–535. [Google Scholar]
- Sceniak, M.P.; Fedder, K.N.; Wang, Q.; Droubi, S.; Babcock, K.; Patwardhan, S.; Wright-Zornes, J.; Pham, L.; Sabo, S.L. An autism-associated mutation in GluN2B prevents NMDA receptor trafficking and interferes with dendrite growth. J. Cell Sci. 2019, 132, jcs232892. [Google Scholar] [PubMed] [Green Version]
- Liu, S.; Zhou, L.; Yuan, H.; Vieira, M.; Sanz-Clemente, A.; Badger, J.D.; Lu, W.; Traynelis, S.F.; Roche, K.W. A rare variant identified within the GluN2B C-terminus in a patient with autism affects NMDA receptor surface expression and spine density. J. Neurosci. 2017, 37, 4093–4102. [Google Scholar] [PubMed]
- Yuen, E.Y.; Jiang, Q.; Chen, P.; Gu, Z.; Feng, J.; Yan, Z. Serotonin 5-HT1A receptors regulate NMDA receptor channels through a microtubule-dependent mechanism. J. Neurosci. 2005, 25, 5488–5501. [Google Scholar] [PubMed] [Green Version]
- Field, T.; Diego, M.; Hernandez-Reif, M.; Figueiredo, B.; Deeds, O.; Ascencio, A.; Schanberg, S.; Kuhn, C. Prenatal dopamine and neonatal behavior and biochemistry. Infant Behav. Dev. 2008, 31, 590–593. [Google Scholar] [PubMed] [Green Version]
- McCarthy, D.; Lueras, P.; Bhide, P.G. Elevated dopamine levels during gestation produce region-specific decreases in neurogenesis and subtle deficits in neuronal numbers. Brain Res. 2007, 1182, 11–25. [Google Scholar]
- Money, K.M.; Stanwood, G.D. Developmental origins of brain disorders: Roles for dopamine. Front. Cell. Neurosci. 2013, 7, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laplante, F.; Nakagawasai, O.; Srivastava, L.K.; Quirion, R. Alterations in behavioral responses to a cholinergic agonist in post-pubertal rats with neonatal ventral hippocampal lesions: Relationship to changes in muscarinic receptor levels. Neuropsychopharmacol 2005, 30, 1076–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, R.; Ross, R.G. Prenatal choline and the development of schizophrenia. Shanghai Arch. Psychiatry 2015, 27, 90–102. [Google Scholar] [PubMed]
- Yang, S.N.; Yang, J.-M.; Wu, J.-N.; Kao, Y.-H.; Hsieh, W.-Y.; Chao, C.C.; Tao, P.-L. Prenatal exposure to morphine alters kinetic properties of NMDA receptor-mediated synaptic currents in the hippocampus of rat offspring. Hippocampus 2000, 10, 654–662. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Amplitude Inhibition (%) | Steady-State Inhibition (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| MeanDiff | SEM | q Value | p (<|q|) | MeanDiff | SEM | q Value | p (<|q|) | |
| 5-MT Gest Vs. Control | 14.73 | 3.72 | 5.6 | ** | 15.91 | 3.79 | 5.94 | *** |
| 5-MT Pre-Post Vs. Control | 19.10 | 3.48 | 7.77 | *** | 20.41 | 3.54 | 8.15 | *** |
| 5-MT Pre-Post Vs. 5-MT Gest | 4.37 | 3.65 | 1.69 | ns. | 4.50 | 3.72 | 1.71 | ns. |
| NMDA(µM) | Peak Amplitude (pA/pF) | Steady-State (pA/pF) | ||
|---|---|---|---|---|
| Control | 5-MT Gest | Control | 5-MT Gest | |
| 0.3 | −0.95 ± 0.20 (9) | −0.71 ± 0.11 (7) | −0.25 ± 0.13 (9) | −0.26 ± 0.08 (7) |
| 1 | −1.34 ± 0.30 (9) | −0.95 ± 0.19 (9) | −0.41 ± 0.27 (9) | −0.52 ± 0.13 (9) |
| 3 | −3.05 ± 0.38 (8) | −2.95 ± 0.40 (9) | −2.13 ± 0.37 (8) | −2.24 ± 0.38 (9) |
| 10 | −9.26 ± 0.97 (9) | −7.61 ± 0.88 (9) | −6.47 ± 1.00 (9) | −5.12 ± 0.84 (9) |
| 30 | −31.85 ± 3.08 (9) | −26.74 ± 2.22 (9) | −19.42 ± 2.76 (9) | −14.90 ± 1.95 (9) |
| 100 | −61.18 ± 4.27 (9) | −55.34 ± 6.05 (9) | −25.25 ± 4.36 (9) | −24.54 ± 5.00 (9) |
| 300 | −80.80 ± 3.88 (9) | −72.21 ± 6.24 (9) | −26.11 ± 4.35 (9) | −26.20 ± 5.57 (9) |
| 5-HT (µM) | Peak Amplitude Inhibition (%) | Steady-State Inhibition (%) | ||||
|---|---|---|---|---|---|---|
| Control | 5-MT Gest | p ( < |t|) | Control | 5-MT Gest | p ( < |t|) | |
| 1 | 11.33 ± 0.90 (13) | 15.68 ± 3.51 (7) | ns. | 13.73 ± 2.69 (13) | 22.24 ± 1.22 (7) | * |
| 3 | 13.64 ± 1.65 (9) | 16.63 ± 1.57 (13) | ns. | 20.44 ± 2.45 (9) | 21.97 ± 1.73 (13) | ns. |
| 10 | 17.88 ± 0.50 (14) | 18.05 ± 1.94 (11) | ns. | 26.64 ± 2.33 (14) | 26.38 ± 2.23 (11) | ns. |
| 30 | 24.23 ± 2.06 (16) | 36.92 ± 2.69 (16) | *** | 35.77 ± 2.29 (16) | 46.94 ± 3.29 (16) | * |
| 100 | 47.75 ± 2.81 (12) | 56.51 ± 2.48 (14) | * | 56.99 ± 3.48 (12) | 66.98 ± 2.98 (14) | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Carballo, G.; Ruíz-Luna, E.A.; López-López, G.; Manjarrez, E.; Flores-Hernández, J. Changes in Serotonin Modulation of Glutamate Currents in Pyramidal Offspring Cells of Rats Treated With 5-MT during Gestation. Brain Sci. 2020, 10, 221. https://doi.org/10.3390/brainsci10040221
Hernández-Carballo G, Ruíz-Luna EA, López-López G, Manjarrez E, Flores-Hernández J. Changes in Serotonin Modulation of Glutamate Currents in Pyramidal Offspring Cells of Rats Treated With 5-MT during Gestation. Brain Sciences. 2020; 10(4):221. https://doi.org/10.3390/brainsci10040221
Chicago/Turabian StyleHernández-Carballo, Gustavo, Evelyn A. Ruíz-Luna, Gustavo López-López, Elias Manjarrez, and Jorge Flores-Hernández. 2020. "Changes in Serotonin Modulation of Glutamate Currents in Pyramidal Offspring Cells of Rats Treated With 5-MT during Gestation" Brain Sciences 10, no. 4: 221. https://doi.org/10.3390/brainsci10040221
APA StyleHernández-Carballo, G., Ruíz-Luna, E. A., López-López, G., Manjarrez, E., & Flores-Hernández, J. (2020). Changes in Serotonin Modulation of Glutamate Currents in Pyramidal Offspring Cells of Rats Treated With 5-MT during Gestation. Brain Sciences, 10(4), 221. https://doi.org/10.3390/brainsci10040221