Cerebellar Transcranial Direct Current Stimulation Improves Maximum Isometric Force Production during Isometric Barbell Squats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
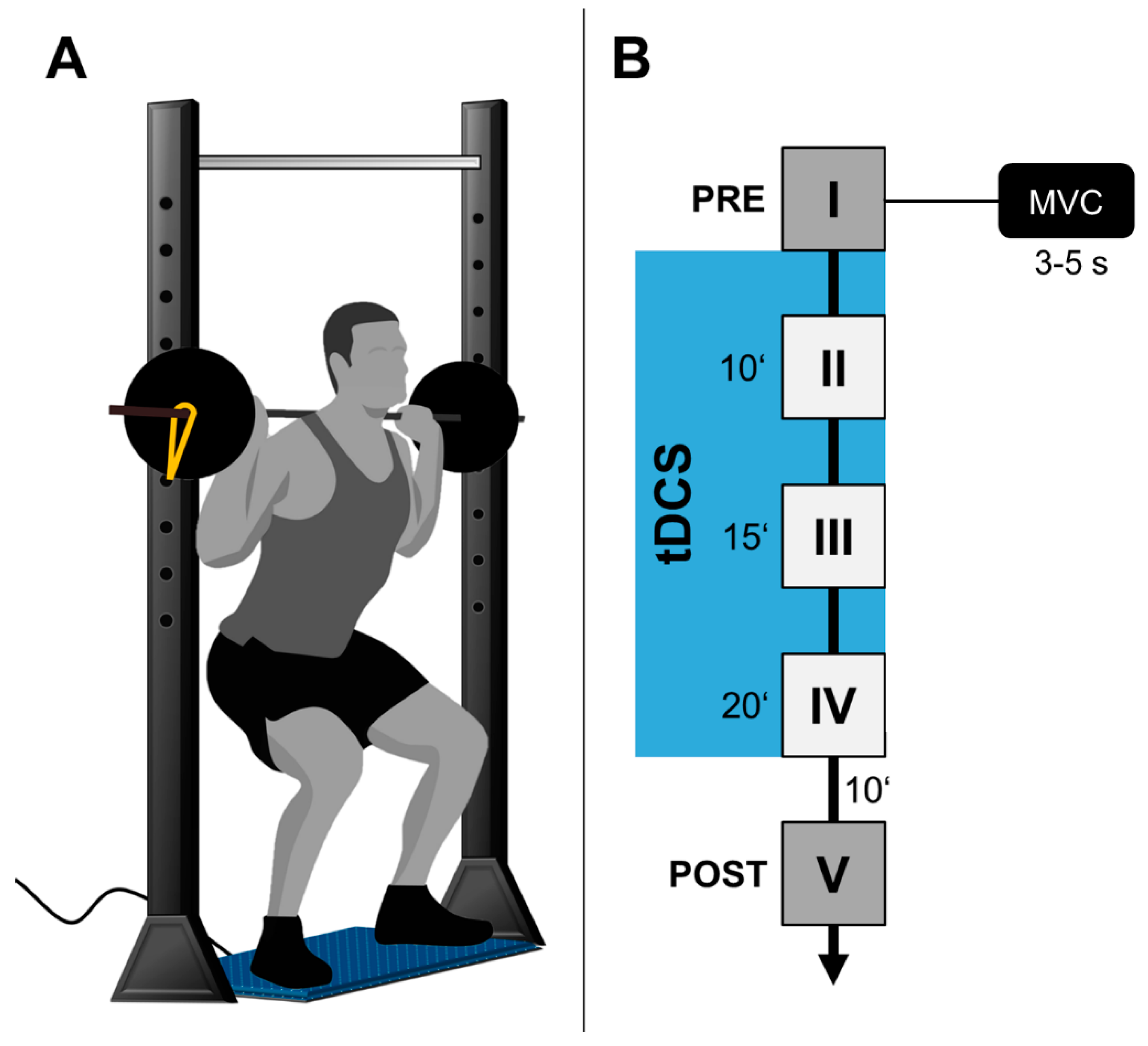
2.3. Behavioral Task (iBS)
2.4. Transcranial Direct Current Stimulation
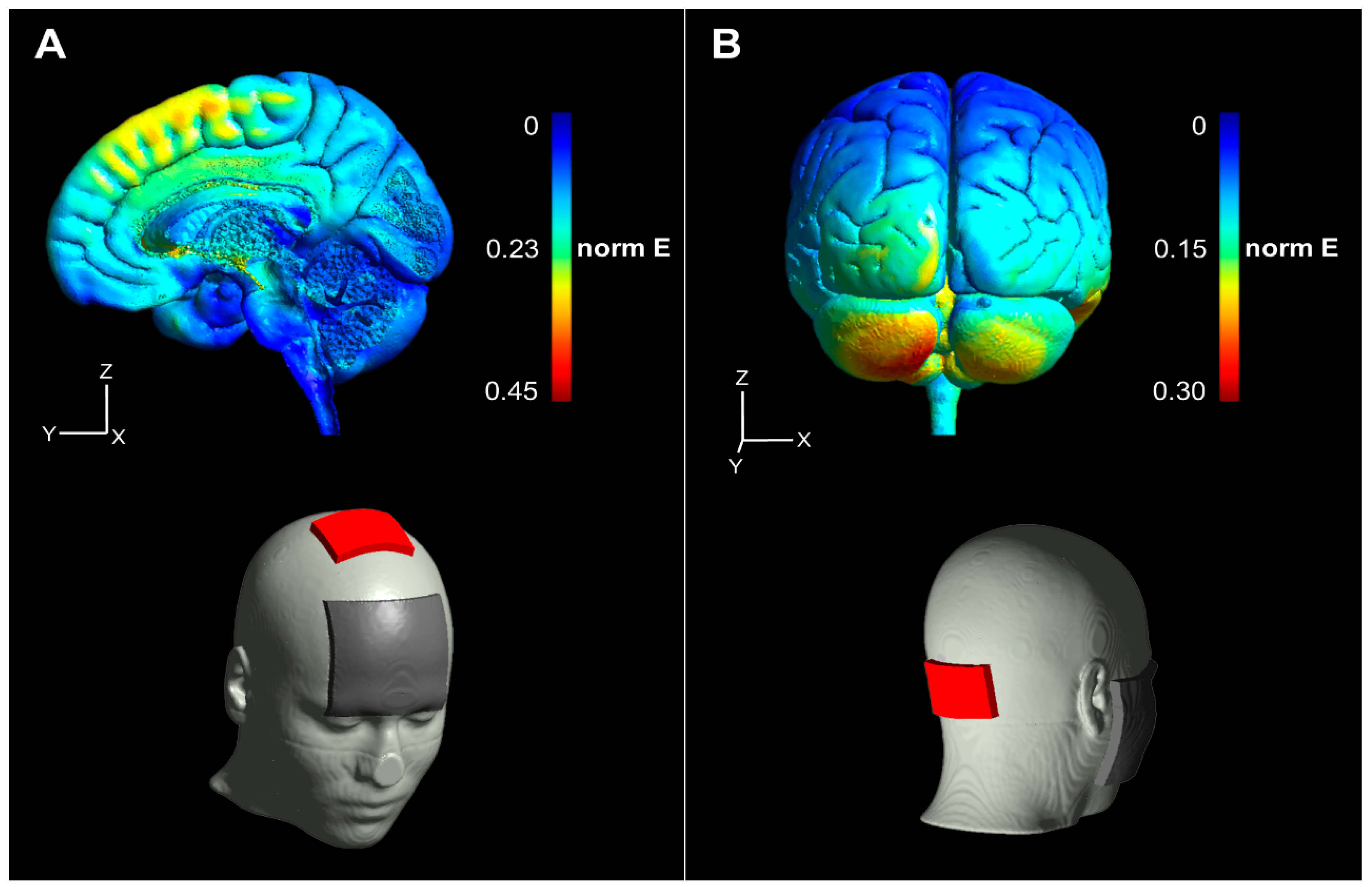
2.5. tDCS Current Flow Simulation
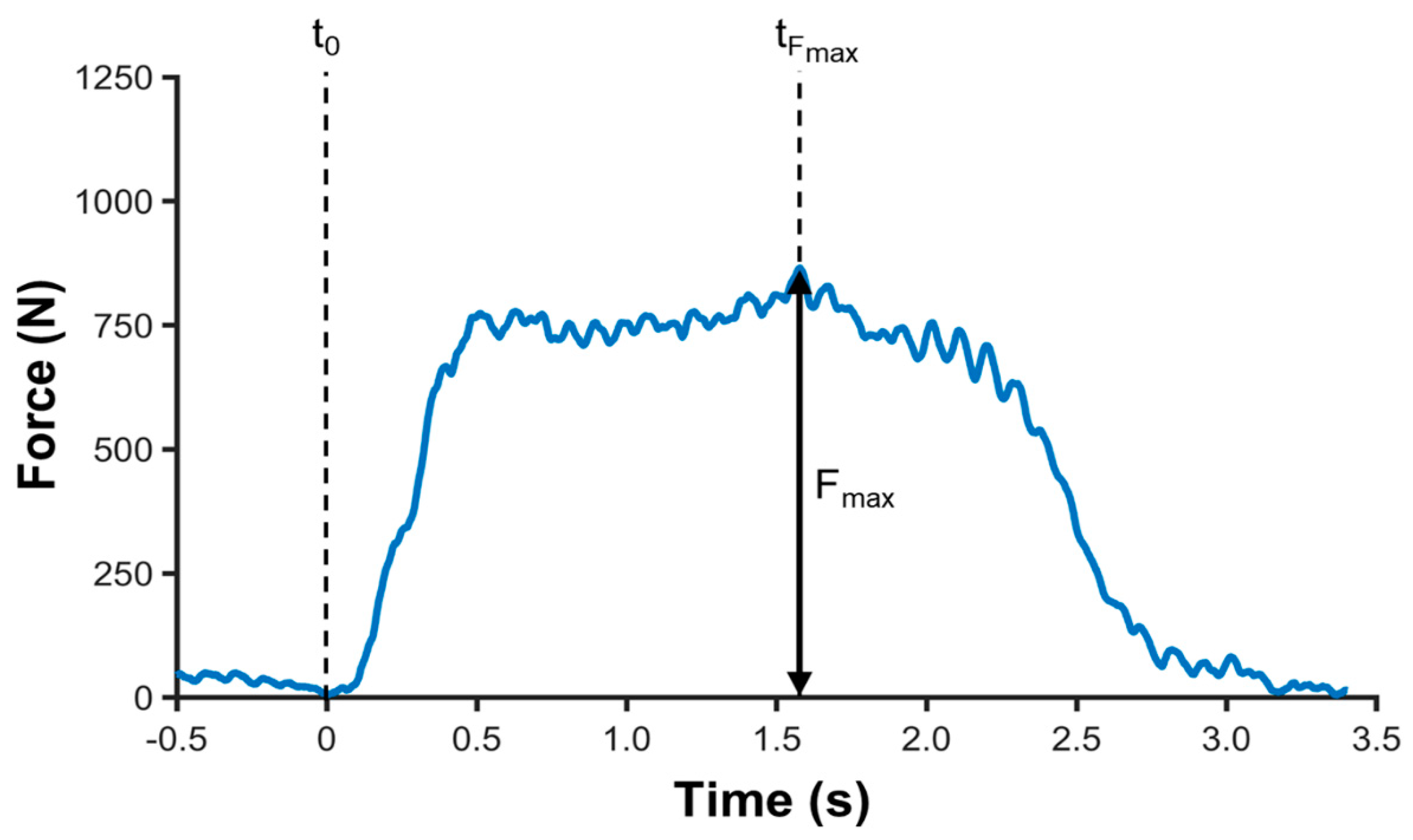
2.6. Data Analysis
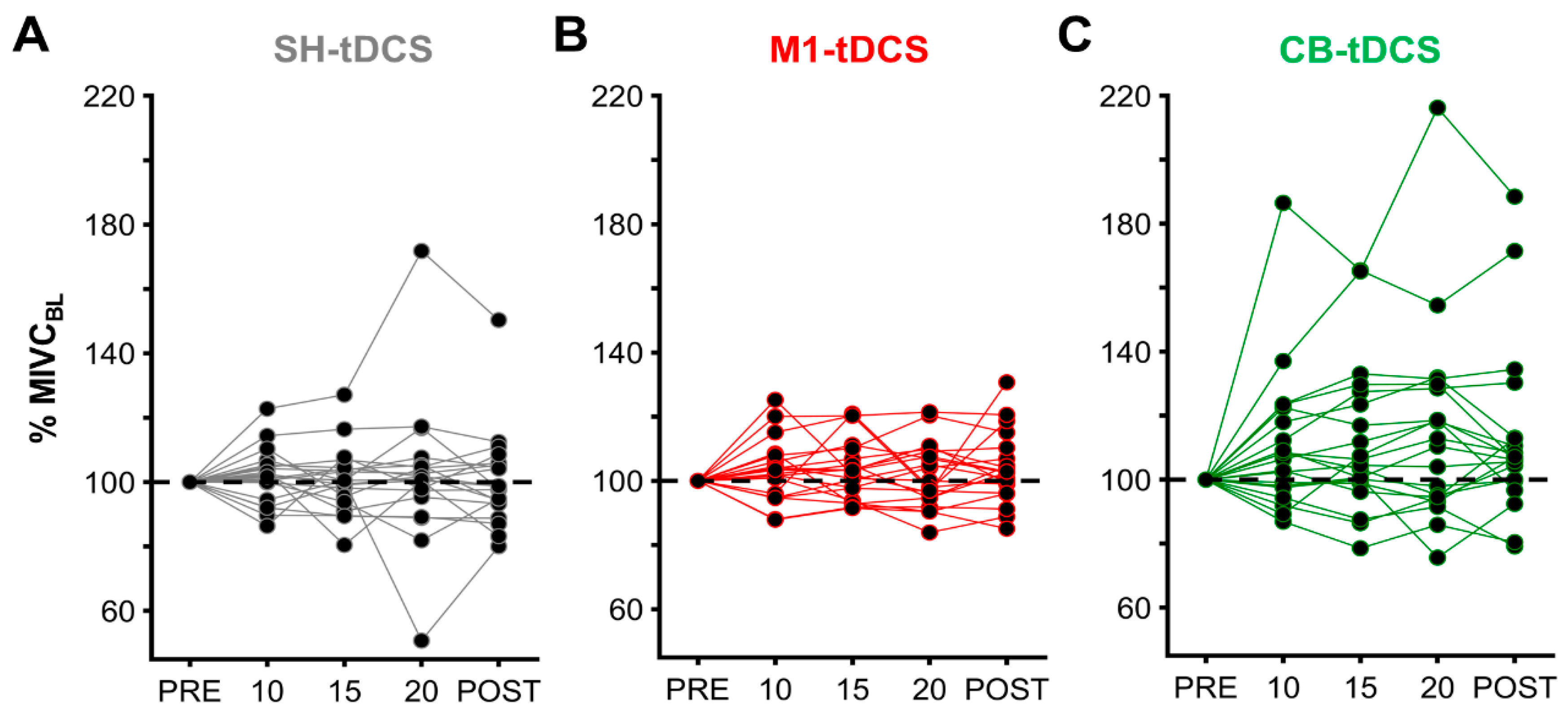
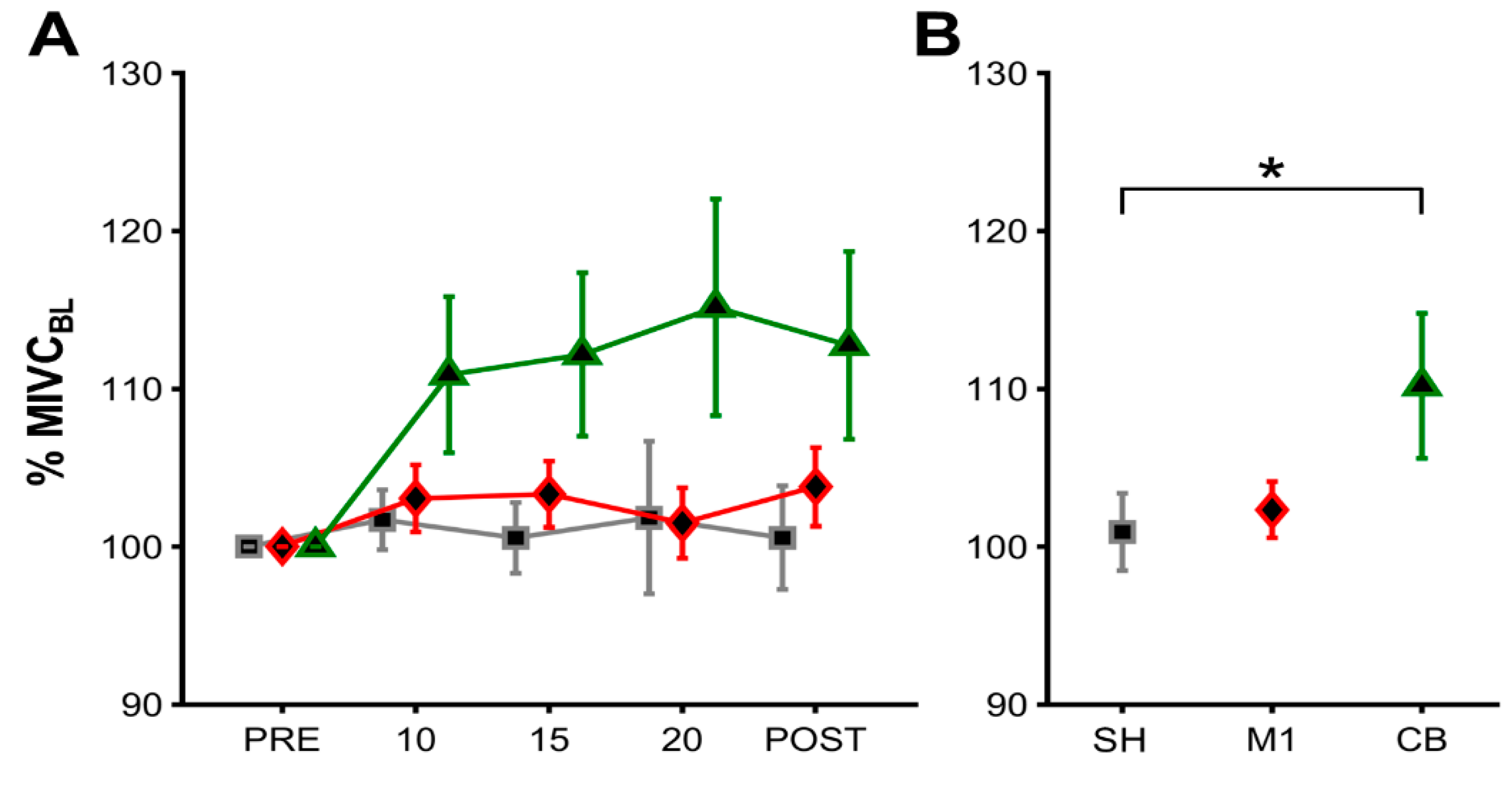
3. Results
MIVC During iBS
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beckham, G.; Mizuguchi, S.; Carter, C.; Sato, K.; Ramsey, M.; Lamont, H.; Hornsby, G.; Haff, G.; Stone, M. Relationships of isometric mid-thigh pull variables to weightlifting performance. J. Sports Med. Phys. Fit. 2013, 53, 573–581. [Google Scholar]
- Izquierdo, M.; Aguado, X.; Gonzalez, R.; Lopez, J.; Häkkinen, K. Maximal and explosive force production capacity and balance performance in men of different ages. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 79, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, K.; Alen, M.; Kraemer, W.; Gorostiaga, E.; Izquierdo, M.; Rusko, H.; Mikkola, J.; Häkkinen, A.; Valkeinen, H.; Kaarakainen, E. Neuromuscular adaptations during concurrent strength and endurance training versus strength training. Eur. J. Appl. Physiol. 2003, 89, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Grodd, W.; Hulsmann, E.; Lotze, M.; Wildgruber, D.; Erb, M. Sensorimotor mapping of the human cerebellum: fMRI evidence of somatotopic organization. Hum. Brain Mapp. 2001, 13, 55–73. [Google Scholar] [CrossRef]
- Wiestler, T.; McGonigle, D.J.; Diedrichsen, J. Integration of sensory and motor representations of single fingers in the human cerebellum. J. Neurophysiol. 2011, 105, 3042–3053. [Google Scholar] [CrossRef] [Green Version]
- Spraker, M.B.; Corcos, D.M.; Kurani, A.S.; Prodoehl, J.; Swinnen, S.P.; Vaillancourt, D.E. Specific cerebellar regions are related to force amplitude and rate of force development. Neuroimage 2012, 59, 1647–1656. [Google Scholar] [CrossRef] [Green Version]
- Dettmers, C.; Fink, G.R.; Lemon, R.N.; Stephan, K.M.; Passingham, R.E.; Silbersweig, D.; Holmes, A.; Ridding, M.C.; Brooks, D.J.; Frackowiak, R.S. Relation between cerebral activity and force in the motor areas of the human brain. J. Neurophysiol. 1995, 74, 802–815. [Google Scholar] [CrossRef]
- Lattari, E.; Oliveira, B.R.R.; Monteiro Junior, R.S.; Marques Neto, S.R.; Oliveira, A.J.; Maranhao Neto, G.A.; Machado, S.; Budde, H. Acute effects of single dose transcranial direct current stimulation on muscle strength: A systematic review and meta-analysis. PLoS ONE 2018, 13, e0209513. [Google Scholar] [CrossRef] [Green Version]
- Cogiamanian, F.; Marceglia, S.; Ardolino, G.; Barbieri, S.; Priori, A. Improved isometric force endurance after transcranial direct current stimulation over the human motor cortical areas. Eur. J. Neurosci. 2007, 26, 242–249. [Google Scholar] [CrossRef]
- Lattari, E.; Rosa Filho, B.J.; Fonseca Junior, S.J.; Murillo-Rodriguez, E.; Rocha, N.; Machado, S.; Maranhao Neto, G.A. Effects on Volume Load and Ratings of Perceived Exertion in Individuals Advanced Weight-Training After Transcranial Direct Current Stimulation. J. Strength Cond. Res. 2018, 34, 89–96. [Google Scholar] [CrossRef]
- Lattari, E.; Andrade, M.L.; Filho, A.S.; Moura, A.M.; Neto, G.M.; Silva, J.G.; Rocha, N.B.; Yuan, T.F.; Arias-Carrion, O.; Machado, S. Can Transcranial Direct Current Stimulation Improve the Resistance Strength and Decrease the Rating Perceived Scale in Recreational Weight-Training Experience? J. Strength Cond. Res. 2016, 30, 3381–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmoula, A.; Baudry, S.; Duchateau, J. Anodal transcranial direct current stimulation enhances time to task failure of a submaximal contraction of elbow flexors without changing corticospinal excitability. Neuroscience 2016, 322, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Vargas, V.Z.; Baptista, A.F.; Pereira, G.O.C.; Pochini, A.C.; Ejnisman, B.; Santos, M.B.; Joao, S.M.A.; Hazime, F.A. Modulation of Isometric Quadriceps Strength in Soccer Players With Transcranial Direct Current Stimulation: A Crossover Study. J. Strength Cond. Res. 2018, 32, 1336–1341. [Google Scholar] [CrossRef]
- Tanaka, S.; Hanakawa, T.; Honda, M.; Watanabe, K. Enhancement of pinch force in the lower leg by anodal transcranial direct current stimulation. Exp. Brain Res. 2009, 196, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Angius, L.; Pageaux, B.; Hopker, J.; Marcora, S.M.; Mauger, A.R. Transcranial direct current stimulation improves isometric time to exhaustion of the knee extensors. Neuroscience 2016, 339, 363–375. [Google Scholar] [CrossRef]
- Lattari, E.; Campos, C.; Lamego, M.K.; Passos de Souza, S.L.; Neto, G.M.; Rocha, N.B.; Jose de Oliveira, A.; Carpenter, S.; Machado, S. Can transcranial direct current stimulation improve muscle power in individuals with advanced resistance training experience? J. Strength Cond. Res. 2017. [Google Scholar] [CrossRef]
- Ciccone, A.B.; Deckert, J.A.; Schlabs, C.R.; Tilden, M.J.; Herda, T.J.; Gallagher, P.M.; Weir, J.P. Transcranial Direct Current Stimulation of the Temporal Lobe Does Not Affect High Intensity Work Capacity. J. Strength Cond. Res. 2018. [Google Scholar] [CrossRef]
- Maeda, K.; Yamaguchi, T.; Tatemoto, T.; Kondo, K.; Otaka, Y.; Tanaka, S. Transcranial Direct Current Stimulation Does Not Affect Lower Extremity Muscle Strength Training in Healthy Individuals: A Triple-Blind, Sham-Controlled Study. Front. Neurosci. 2017, 11, 179. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527 Pt 3, 633–639. [Google Scholar] [CrossRef]
- Hendy, A.M.; Kidgell, D.J. Anodal-tDCS applied during unilateral strength training increases strength and corticospinal excitability in the untrained homologous muscle. Exp. Brain Res. 2014, 232, 3243–3252. [Google Scholar] [CrossRef]
- Jacobs, J.V.; Horak, F.B. Cortical control of postural responses. J. Neural Transm. (Vienna) 2007, 114, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M. Historical review of the significance of the cerebellum and the role of Purkinje cells in motor learning. Ann. N. Y. Acad. Sci. 2002, 978, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Ioffe, M.E.; Chernikova, L.A.; Ustinova, K.I. Role of cerebellum in learning postural tasks. Cerebellum 2007, 6, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.A.; Keating, J.G.; Goodkin, H.P.; Bastian, A.J.; Thach, W.T. Throwing while looking through prisms. I. Focal olivocerebellar lesions impair adaptation. Brain 1996, 119 Pt 4, 1183–1198. [Google Scholar] [CrossRef]
- Timmann, D.; Brandauer, B.; Hermsdorfer, J.; Ilg, W.; Konczak, J.; Gerwig, M.; Gizewski, E.R.; Schoch, B. Lesion-symptom mapping of the human cerebellum. Cerebellum 2008, 7, 602–606. [Google Scholar] [CrossRef]
- Topka, H.; Konczak, J.; Schneider, K.; Boose, A.; Dichgans, J. Multijoint arm movements in cerebellar ataxia: Abnormal control of movement dynamics. Exp. Brain Res. 1998, 119, 493–503. [Google Scholar] [CrossRef]
- Bastian, A.J.; Mink, J.W.; Kaufman, B.A.; Thach, W.T. Posterior vermal split syndrome. Ann. Neurol. 1998, 44, 601–610. [Google Scholar] [CrossRef]
- Thach, W.T. Timing of activity in cerebellar dentate nucleus and cerebral motor cortex during prompt volitional movement. Brain Res. 1975, 88, 233–241. [Google Scholar] [CrossRef]
- Goodkin, H.P.; Thach, W.T. Cerebellar control of constrained and unconstrained movements. I. Nuclear inactivation. J. Neurophysiol. 2003, 89, 884–895. [Google Scholar] [CrossRef]
- Jayaram, G.; Tang, B.; Pallegadda, R.; Vasudevan, E.V.; Celnik, P.; Bastian, A. Modulating locomotor adaptation with cerebellar stimulation. J. Neurophysiol. 2012, 107, 2950–2957. [Google Scholar] [CrossRef] [Green Version]
- Poortvliet, P.; Hsieh, B.; Cresswell, A.; Au, J.; Meinzer, M. Cerebellar transcranial direct current stimulation improves adaptive postural control. Clin. Neurophysiol. 2018, 129, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandvliet, S.B.; Meskers, C.G.M.; Kwakkel, G.; van Wegen, E.E.H. Short-Term Effects of Cerebellar tDCS on Standing Balance Performance in Patients with Chronic Stroke and Healthy Age-Matched Elderly. Cerebellum 2018, 17, 575–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, K.M.; Enders, A.; Thier, W.; Batsikadze, G.; Ludolph, N.; Ilg, W.; Timmann, D. Cerebellar tDCS does not improve learning in a complex whole body dynamic balance task in young healthy subjects. PLoS ONE 2016, 11, e0163598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, C.J.; Bailey, J.A.; Zatorre, R.J.; Penhune, V.B. Early musical training and white-matter plasticity in the corpus callosum: Evidence for a sensitive period. J. Neurosci. 2013, 33, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Vollmann, H.; Ragert, P.; Conde, V.; Villringer, A.; Classen, J.; Witte, O.W.; Steele, C.J. Instrument specific use-dependent plasticity shapes the anatomical properties of the corpus callosum: A comparison between musicians and non-musicians. Front. Behav. Neurosci. 2014, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, S.; Shao, J.; Wang, H. Sample Size Calculations in Clinical Research, Chapman & Hall; CRC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Rosner, B. Fundamentals of Biostatistics. Cengage Learning; Inc., Kentucky: Boston, MA, USA, 2010. [Google Scholar]
- Ferrucci, R.; Cortese, F.; Priori, A. Cerebellar tDCS: How to do it. Cerebellum 2015, 14, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Knoch, D.; Nitsche, M.A.; Fischbacher, U.; Eisenegger, C.; Pascual-Leone, A.; Fehr, E. Studying the neurobiology of social interaction with transcranial direct current stimulation—The example of punishing unfairness. Cereb. Cortex 2008, 18, 1987–1990. [Google Scholar] [CrossRef] [Green Version]
- Nitsche, M.A.; Doemkes, S.; Karakose, T.; Antal, A.; Liebetanz, D.; Lang, N.; Tergau, F.; Paulus, W. Shaping the effects of transcranial direct current stimulation of the human motor cortex. J. Neurophysiol. 2007, 97, 3109–3117. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, S.; Stinear, J.W. Focal and bi-directional modulation of lower limb motor cortex using anodal transcranial direct current stimulation. Brain Stimul. 2010, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Laczo, B.; Antal, A.; Rothkegel, H.; Paulus, W. Increasing human leg motor cortex excitability by transcranial high frequency random noise stimulation. Restor. Neurol. Neurosci. 2014, 32, 403–410. [Google Scholar] [CrossRef]
- Kaminski, E.; Steele, C.J.; Hoff, M.; Gundlach, C.; Rjosk, V.; Sehm, B.; Villringer, A.; Ragert, P. Transcranial direct current stimulation (tDCS) over primary motor cortex leg area promotes dynamic balance task performance. Clin. Neurophysiol. 2016, 127, 2455–2462. [Google Scholar] [CrossRef] [PubMed]
- Taubert, M.; Stein, T.; Kreutzberg, T.; Stockinger, C.; Hecker, L.; Focke, A.; Ragert, P.; Villringer, A.; Pleger, B. Remote Effects of Non-Invasive Cerebellar Stimulation on Error Processing in Motor Re-Learning. Brain Stimul. 2016, 9, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Van Selst, M.; Jolicoeur, P. A solution to the effect of sample size on outlier elimination. Q. J. Exp. Psychol. Sect. A 1994, 47, 631–650. [Google Scholar] [CrossRef]
- Miller, J. Reaction time analysis with outlier exclusion: Bias varies with sample size. Q. J. Exp. Psychol. 1991, 43, 907–912. [Google Scholar] [CrossRef]
- Pollet, T.V.; van der Meij, L. To Remove or not to Remove: The Impact of Outlier Handling on Significance Testing in Testosterone Data. Adapt. Hum. Behav. Physiol. 2017, 3, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, D.T.; Norton, J.A.; Roy, F.D.; Gorassini, M.A. Effects of transcranial direct current stimulation on the excitability of the leg motor cortex. Exp. Brain Res. 2007, 182, 281–287. [Google Scholar] [CrossRef]
- Sasaki, N.; Abo, M.; Hara, T.; Yamada, N.; Niimi, M.; Kakuda, W. High-frequency rTMS on leg motor area in the early phase of stroke. Acta Neurol. Belg. 2017, 117, 189–194. [Google Scholar] [CrossRef]
- Jamil, A.; Batsikadze, G.; Kuo, H.I.; Labruna, L.; Hasan, A.; Paulus, W.; Nitsche, M.A. Systematic evaluation of the impact of stimulation intensity on neuroplastic after-effects induced by transcranial direct current stimulation. J. Physiol. 2017, 595, 1273–1288. [Google Scholar] [CrossRef]
- Kan, B.; Dundas, J.E.; Nosaka, K. Effect of transcranial direct current stimulation on elbow flexor maximal voluntary isometric strength and endurance. Appl. Physiol. Nutr. Metab. 2013, 38, 734–739. [Google Scholar] [CrossRef]
- Neuling, T.; Wagner, S.; Wolters, C.H.; Zaehle, T.; Herrmann, C.S. Finite-Element Model Predicts Current Density Distribution for Clinical Applications of tDCS and tACS. Front. Psychiatry 2012, 3, 83. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, C.; Ranganathan, R.; Kantak, S.S.; Dhaher, Y.Y.; Rymer, W.Z. Anodal transcranial direct current stimulation alters elbow flexor muscle recruitment strategies. Brain Stimul. 2014, 7, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Galea, J.M.; Jayaram, G.; Ajagbe, L.; Celnik, P. Modulation of cerebellar excitability by polarity-specific noninvasive direct current stimulation. J. Neurosci. 2009, 29, 9115–9122. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.K.; de Albuquerque, L.L.; Pantovic, M.; Fischer, K.M.; Guadagnoli, M.A.; Riley, Z.A.; Poston, B. Cerebellar Transcranial Direct Current Stimulation Enhances Motor Learning in a Complex Overhand Throwing Task. Cerebellum 2019, 18, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Cantarero, G.; Spampinato, D.; Reis, J.; Ajagbe, L.; Thompson, T.; Kulkarni, K.; Celnik, P. Cerebellar direct current stimulation enhances on-line motor skill acquisition through an effect on accuracy. J. Neurosci. 2015, 35, 3285–3290. [Google Scholar] [CrossRef] [PubMed]
- Jongkees, B.J.; Immink, M.A.; Boer, O.D.; Yavari, F.; Nitsche, M.A.; Colzato, L.S. The Effect of Cerebellar tDCS on Sequential Motor Response Selection. Cerebellum 2019, 18, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carolan, B.; Cafarelli, E. Adaptations in coactivation after isometric resistance training. J. Appl. Physiol. 1992, 73, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, O.M.; Jones, D.A. The role of learning and coordination in strength training. Eur. J. Appl. Physiol. Occup. Physiol. 1986, 55, 100–105. [Google Scholar] [CrossRef]
- Hallett, M.; Shahani, B.T.; Young, R.R. EMG analysis of patients with cerebellar deficits. J. Neurol. Neurosurg. Psychiatry 1975, 38, 1163–1169. [Google Scholar] [CrossRef] [Green Version]
- Hore, J.; Flament, D. Changes in motor cortex neural discharge associated with the development of cerebellar limb ataxia. J. Neurophysiol. 1988, 60, 1285–1302. [Google Scholar] [CrossRef]
- Diener, H.C.; Dichgans, J. Pathophysiology of cerebellar ataxia. Mov. Disord. 1992, 7, 95–109. [Google Scholar] [CrossRef]
- Grimaldi, G.; Oulad Ben Taib, N.; Manto, M.; Bodranghien, F. Marked reduction of cerebellar deficits in upper limbs following transcranial cerebello-cerebral DC stimulation: Tremor reduction and re-programming of the timing of antagonist commands. Front. Syst. Neurosci. 2014, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galea, J.M.; Vazquez, A.; Pasricha, N.; de Xivry, J.J.; Celnik, P. Dissociating the roles of the cerebellum and motor cortex during adaptive learning: The motor cortex retains what the cerebellum learns. Cereb. Cortex 2011, 21, 1761–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehsani, F.; Bakhtiary, A.H.; Jaberzadeh, S.; Talimkhani, A.; Hajihasani, A. Differential effects of primary motor cortex and cerebellar transcranial direct current stimulation on motor learning in healthy individuals: A randomized double-blind sham-controlled study. Neurosci. Res. 2016, 112, 10–19. [Google Scholar] [CrossRef]
- Jayaram, G.; Galea, J.M.; Bastian, A.J.; Celnik, P. Human locomotor adaptive learning is proportional to depression of cerebellar excitability. Cereb. Cortex 2011, 21, 1901–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabe, K.; Livne, O.; Gizewski, E.R.; Aurich, V.; Beck, A.; Timmann, D.; Donchin, O. Adaptation to visuomotor rotation and force field perturbation is correlated to different brain areas in patients with cerebellar degeneration. J. Neurophysiol. 2009, 101, 1961–1971. [Google Scholar] [CrossRef] [Green Version]
- Hazime, F.A.; da Cunha, R.A.; Soliaman, R.R.; Romancini, A.C.B.; Pochini, A.C.; Ejnisman, B.; Baptista, A.F. Anodal Transcranial Direct Current Stimulation (Tdcs) Increases Isometric Strength of Shoulder Rotators Muscles in Handball Players. Int. J. Sports Phys. Ther. 2017, 12, 402–407. [Google Scholar]
- Laakso, I.; Tanaka, S.; Koyama, S.; De Santis, V.; Hirata, A. Inter-subject Variability in Electric Fields of Motor Cortical tDCS. Brain Stimul. 2015, 8, 906–913. [Google Scholar] [CrossRef]
- Laakso, I.; Tanaka, S.; Mikkonen, M.; Koyama, S.; Sadato, N.; Hirata, A. Electric fields of motor and frontal tDCS in a standard brain space: A computer simulation study. Neuroimage 2016, 137, 140–151. [Google Scholar] [CrossRef]
- Wiethoff, S.; Hamada, M.; Rothwell, J.C. Variability in response to transcranial direct current stimulation of the motor cortex. Brain Stimul. 2014, 7, 468–475. [Google Scholar] [CrossRef]
- Madhavan, S.; Sriraman, A.; Freels, S. Reliability and Variability of tDCS Induced Changes in the Lower Limb Motor Cortex. Brain Sci. 2016, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Van Asseldonk, E.H.; Boonstra, T.A. Transcranial Direct Current Stimulation of the Leg Motor Cortex Enhances Coordinated Motor Output During Walking With a Large Inter-Individual Variability. Brain Stimul. 2016, 9, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-I.; Bikson, M.; Datta, A.; Minhas, P.; Paulus, W.; Kuo, M.-F.; Nitsche, M.A. Comparing cortical plasticity induced by conventional and high-definition 4 × 1 ring tDCS: A neurophysiological study. Brain Stimul. 2013, 6, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Flood, A.; Waddington, G.; Keegan, R.J.; Thompson, K.G.; Cathcart, S. The effects of elevated pain inhibition on endurance exercise performance. PeerJ 2017, 5, e3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radel, R.; Tempest, G.; Denis, G.; Besson, P.; Zory, R. Extending the limits of force endurance: Stimulation of the motor or the frontal cortex? Cortex 2017, 97, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R. Effect of different rest intervals on the exercise volume completed during squat bouts. J. Sports Sci. Med. 2005, 4, 361–366. [Google Scholar] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenville, R.; Maudrich, T.; Maudrich, D.; Villringer, A.; Ragert, P. Cerebellar Transcranial Direct Current Stimulation Improves Maximum Isometric Force Production during Isometric Barbell Squats. Brain Sci. 2020, 10, 235. https://doi.org/10.3390/brainsci10040235
Kenville R, Maudrich T, Maudrich D, Villringer A, Ragert P. Cerebellar Transcranial Direct Current Stimulation Improves Maximum Isometric Force Production during Isometric Barbell Squats. Brain Sciences. 2020; 10(4):235. https://doi.org/10.3390/brainsci10040235
Chicago/Turabian StyleKenville, Rouven, Tom Maudrich, Dennis Maudrich, Arno Villringer, and Patrick Ragert. 2020. "Cerebellar Transcranial Direct Current Stimulation Improves Maximum Isometric Force Production during Isometric Barbell Squats" Brain Sciences 10, no. 4: 235. https://doi.org/10.3390/brainsci10040235
APA StyleKenville, R., Maudrich, T., Maudrich, D., Villringer, A., & Ragert, P. (2020). Cerebellar Transcranial Direct Current Stimulation Improves Maximum Isometric Force Production during Isometric Barbell Squats. Brain Sciences, 10(4), 235. https://doi.org/10.3390/brainsci10040235