The Role of BDNF on Aging-Modulation Markers



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of BDNF Solutions
2.2. Astrocytes Isolation
2.3. Primary Cortical Neuronal Cells
2.4. In Vitro Experimental Protocol
2.5. Intestinal Barrier In Vitro Model
2.6. Blood–Brain Barrier (BBB) Experimental Model
2.7. Brain-Derived Neurotrophic Factor (BDNF) Quantification
2.8. MTT Assay
2.9. Crystal Violet Staining
2.10. ROS Production
2.11. NO Production
2.12. Mitochondrial Membrane Potential
2.13. ERK Activation Assay
2.14. Western Blot
2.15. Animal Model
2.16. Statistical Analysis
3. Results
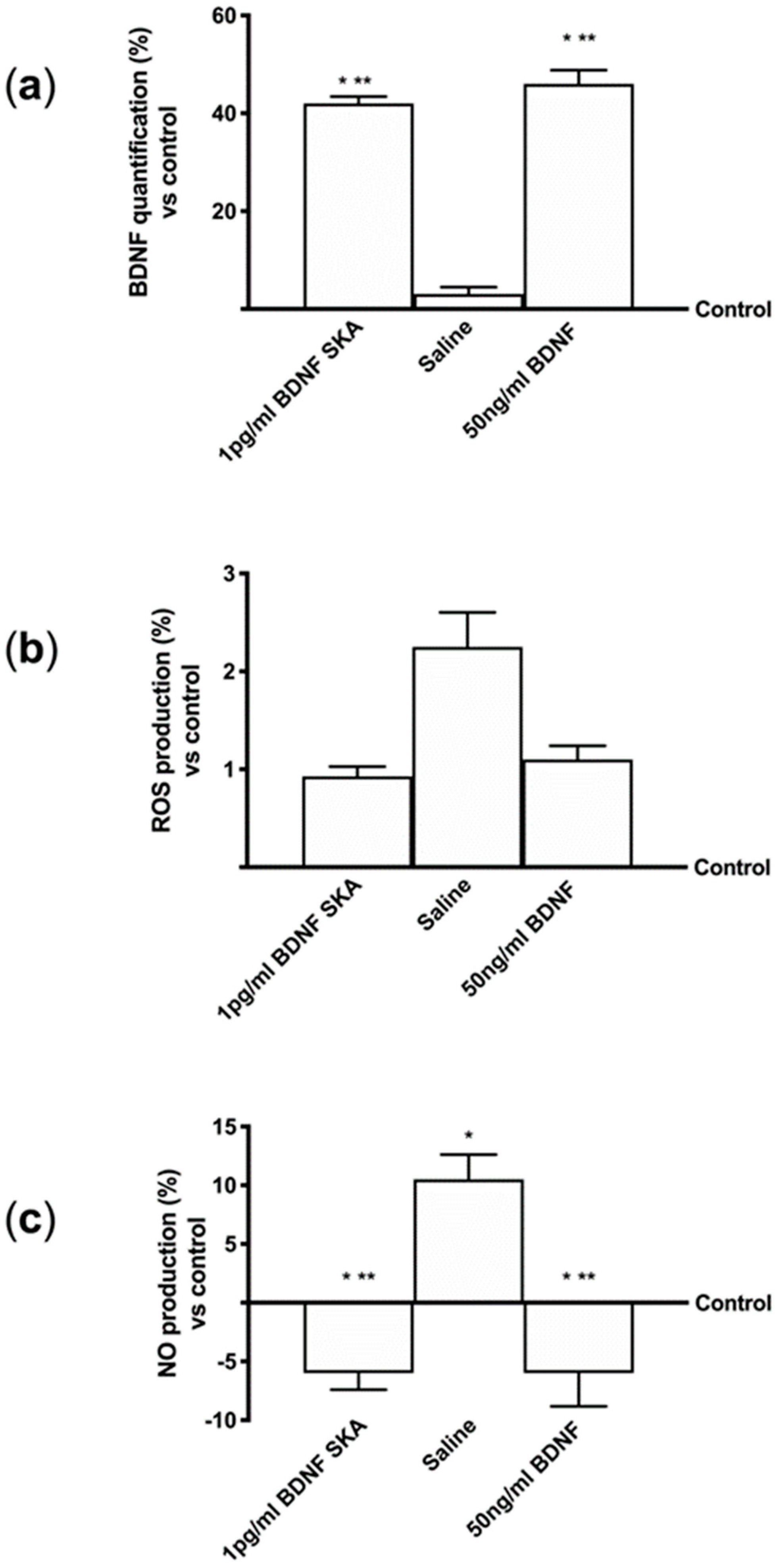
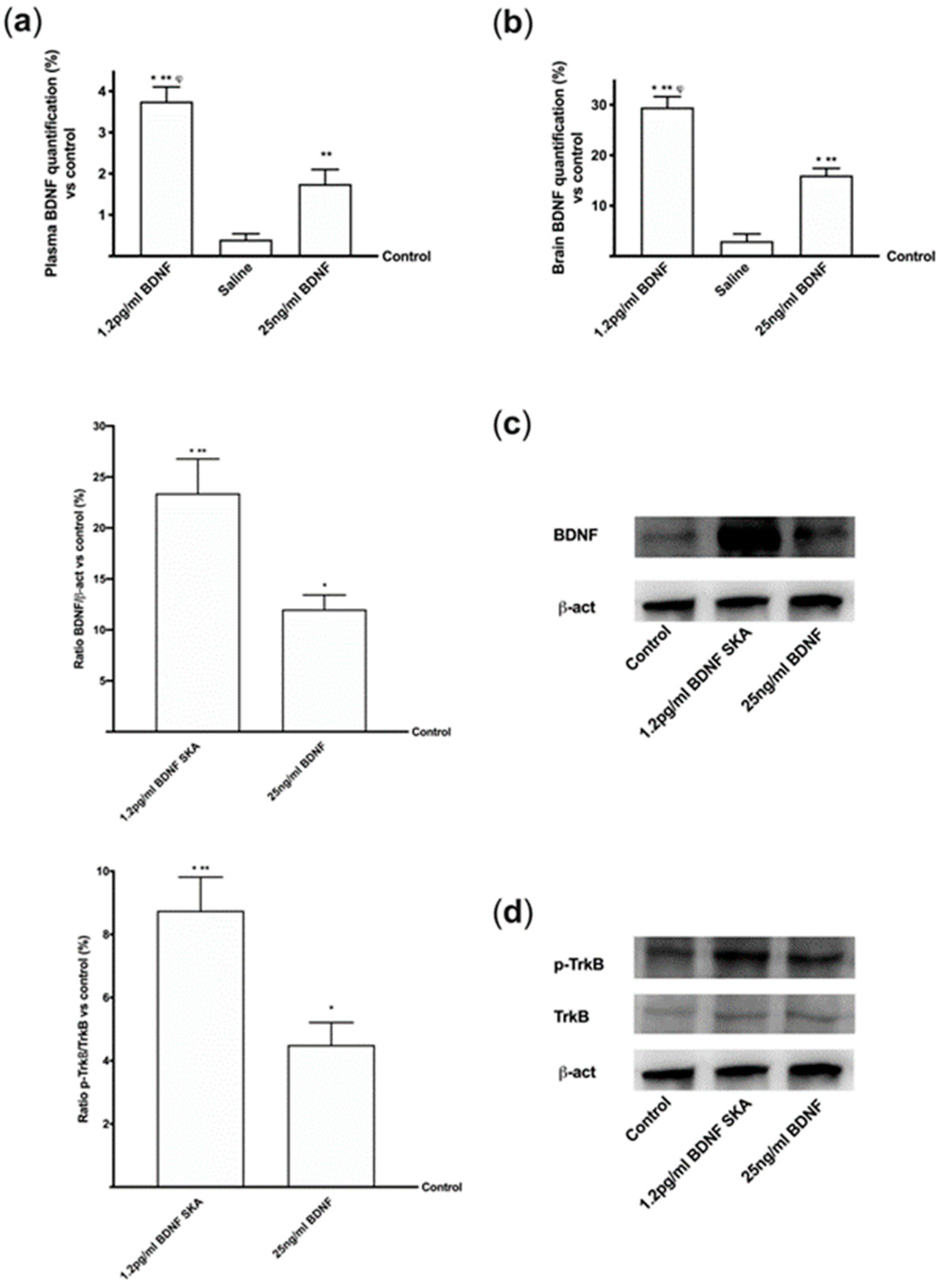
3.1. The Potential Intestinal Absorption as Evaluated In Vitro
3.2. Permeability of BDNF SKA Through Blood–Brain Barrier (BBB)
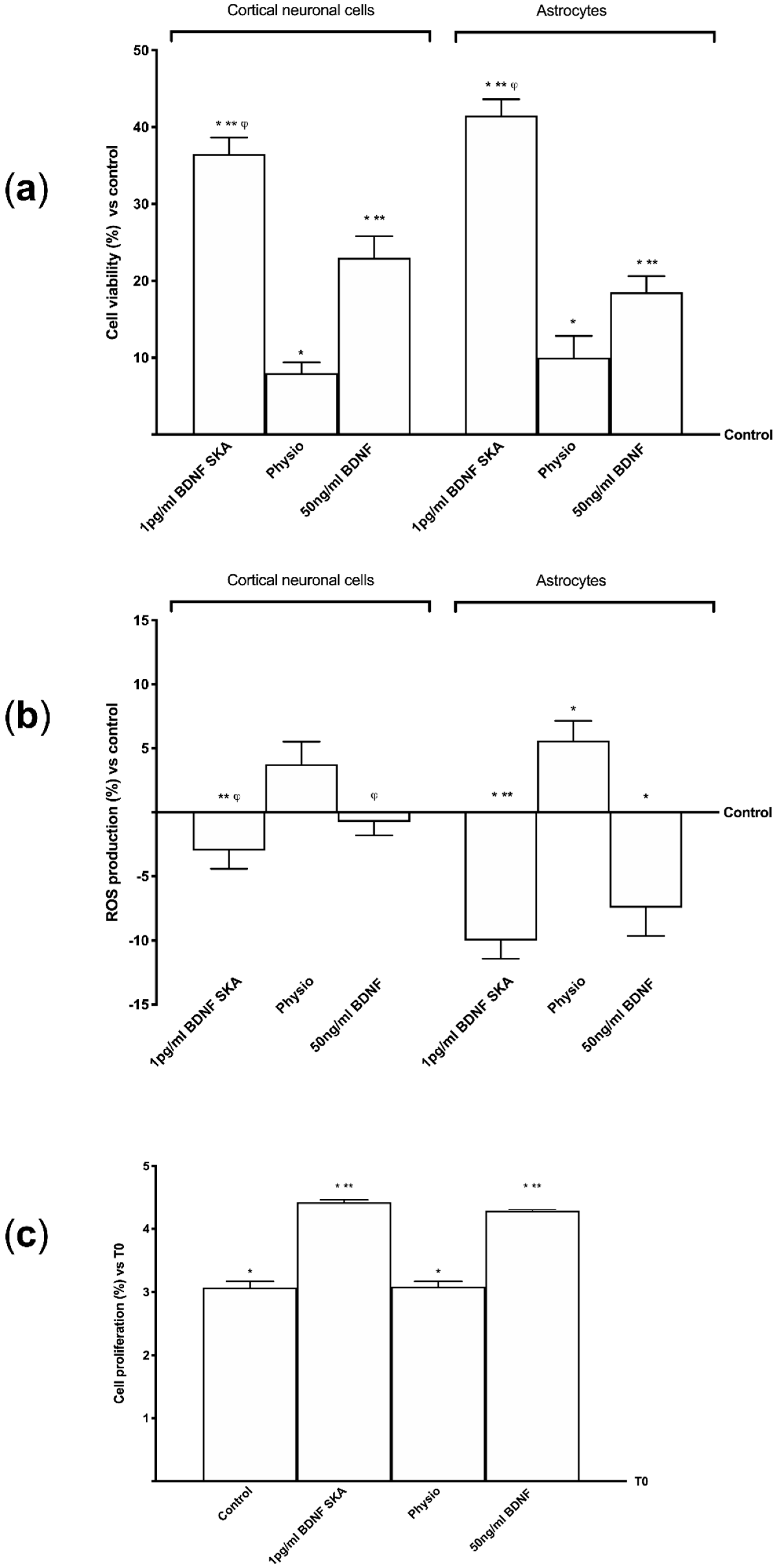
3.3. Topic Action of BDNF SKA on Monolayer Culture
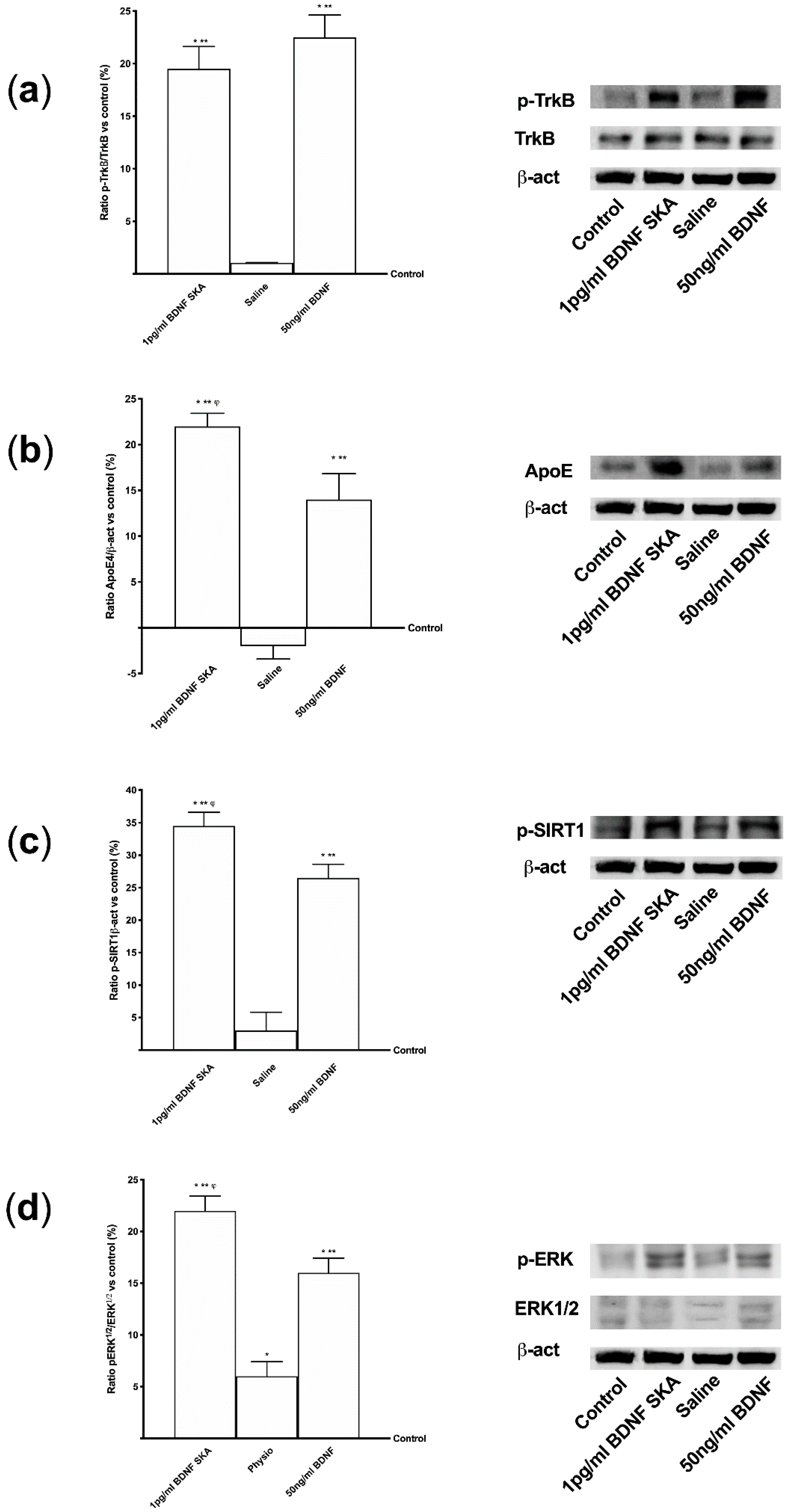
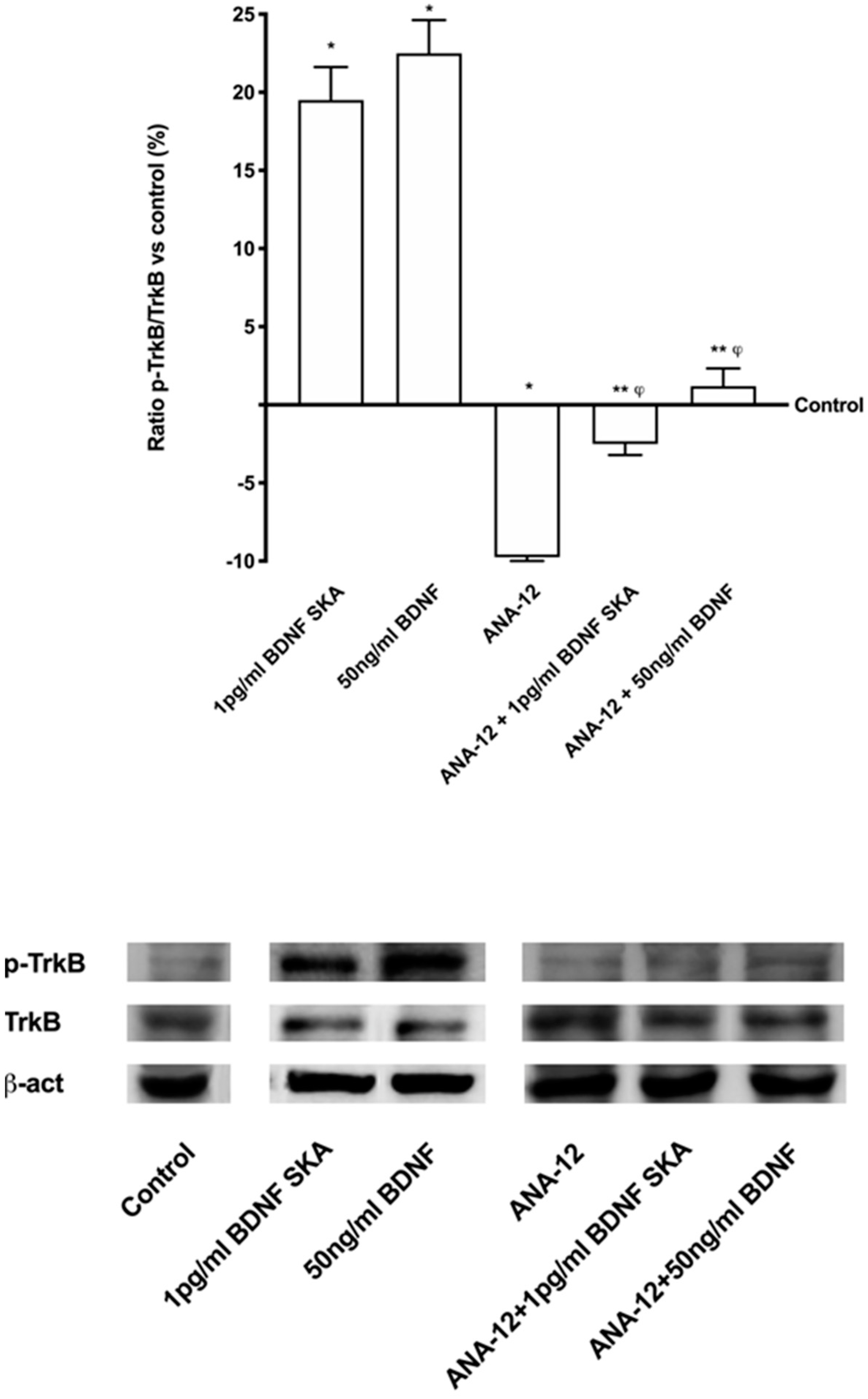
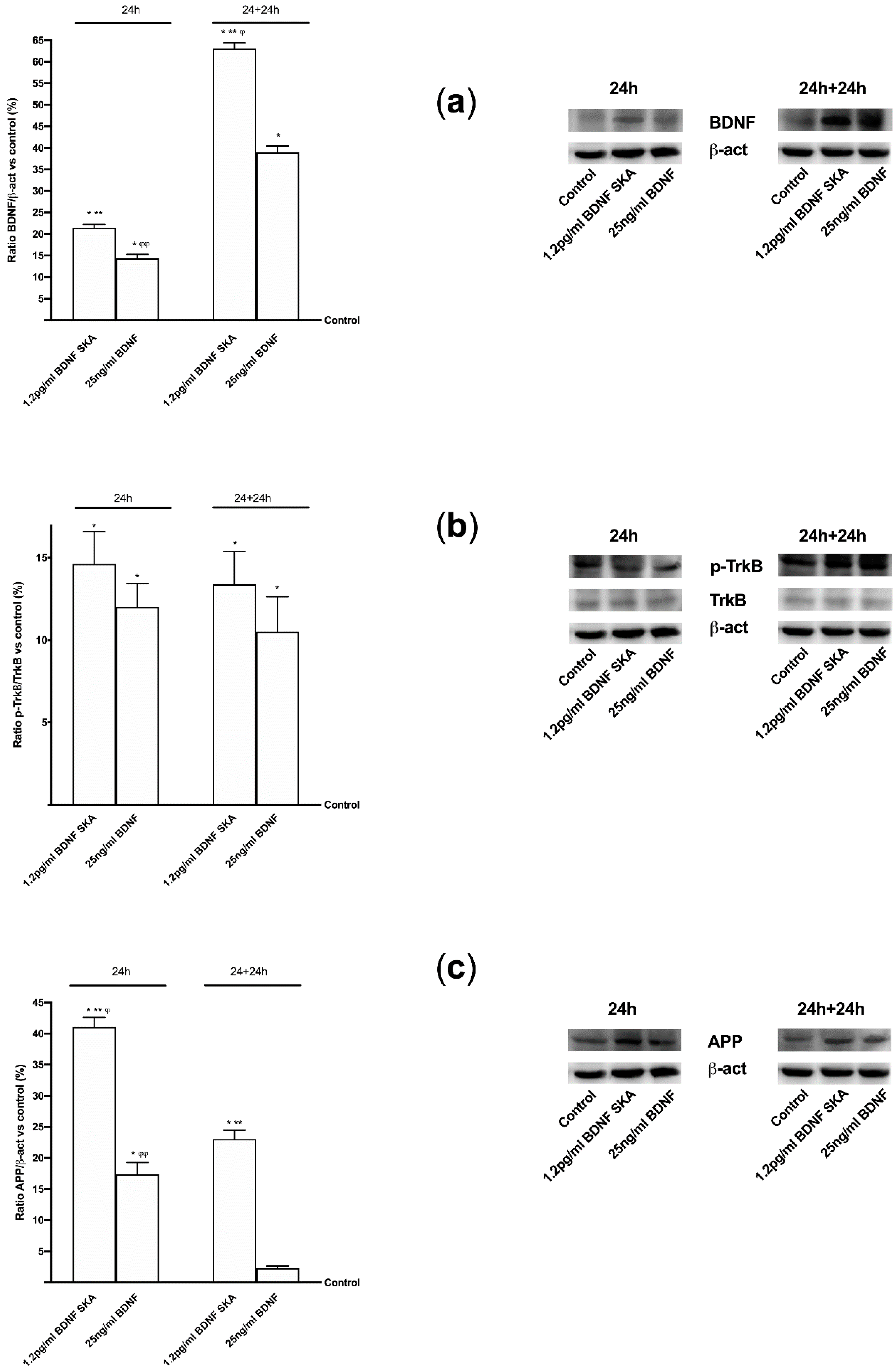
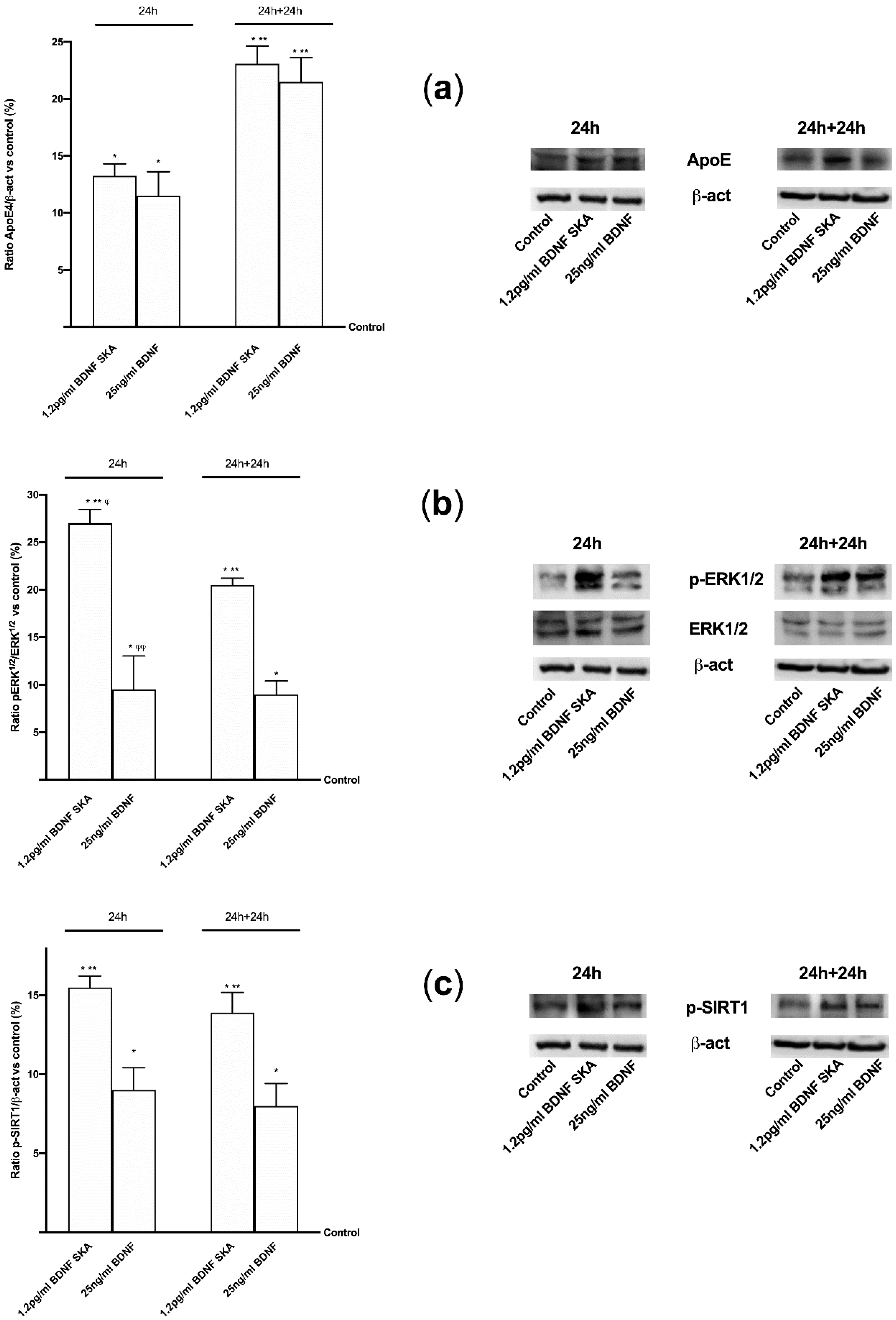
3.4. Intracellular Pathways Activated by BDNF SKA on Monolayer Culture
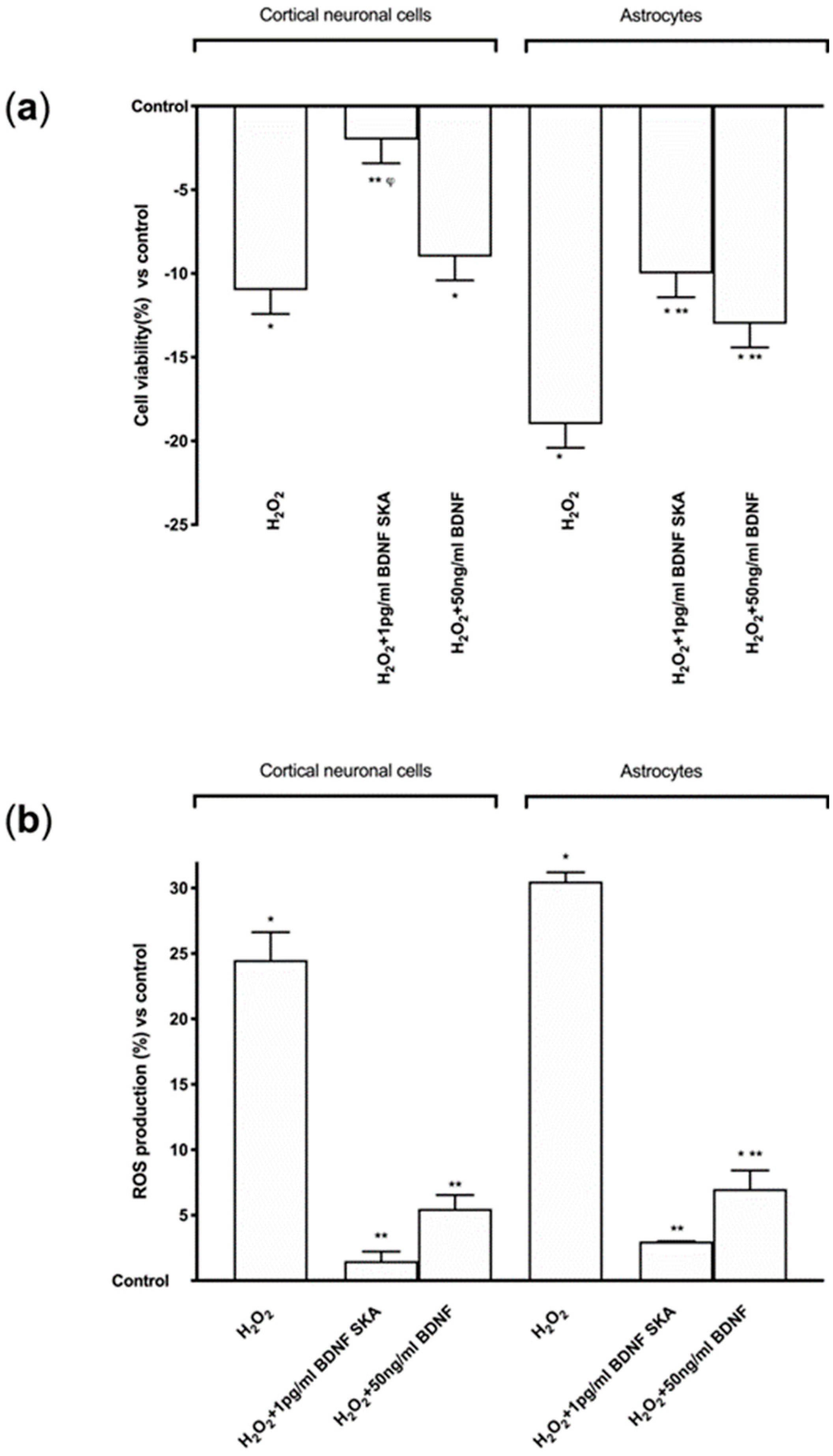
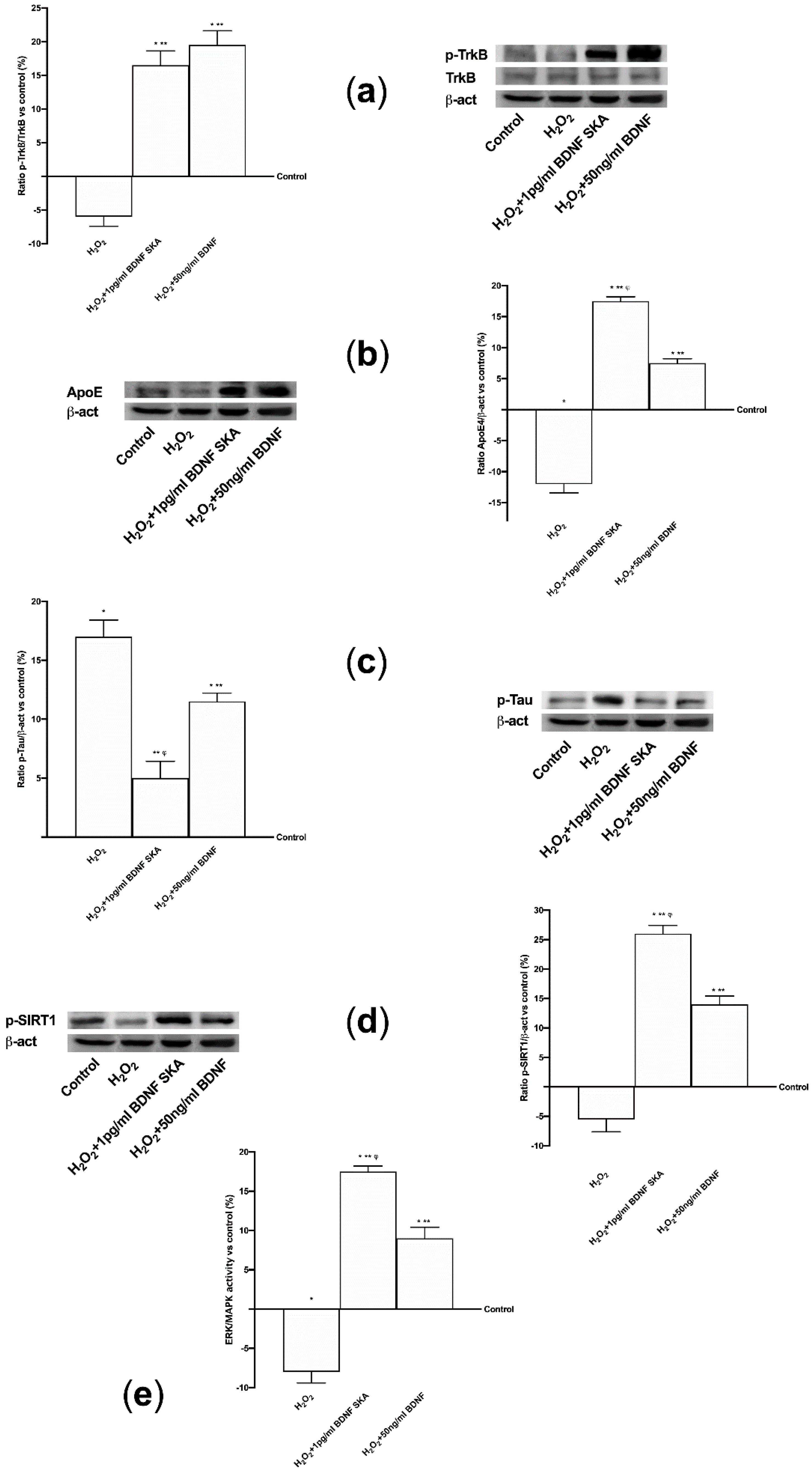
3.5. Effects of BDNF Solutions Under Oxidative Conditions
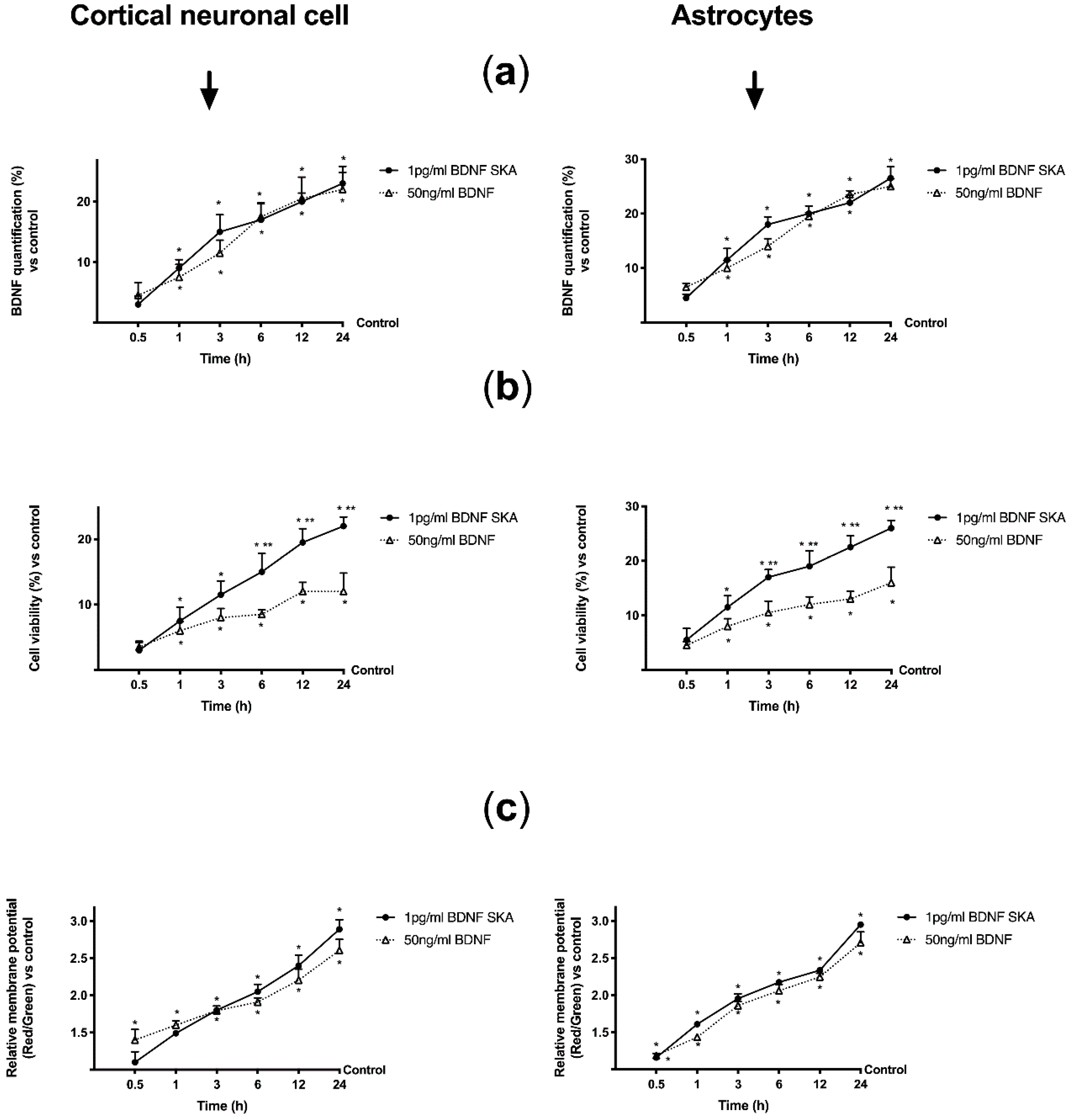
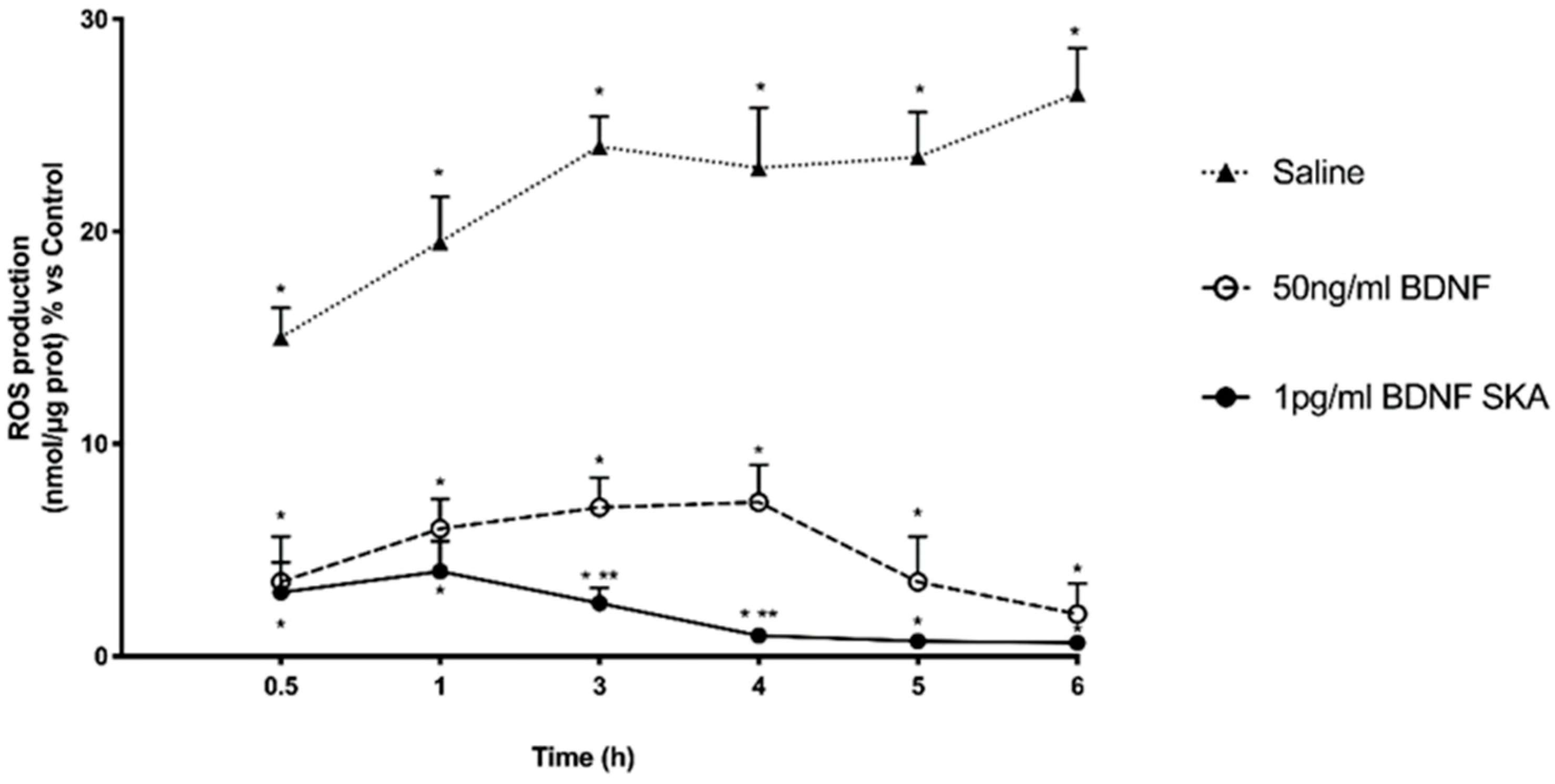
3.6. Daily Duration of the Effects of BDNF Solutions on Neurons and Astrocytes
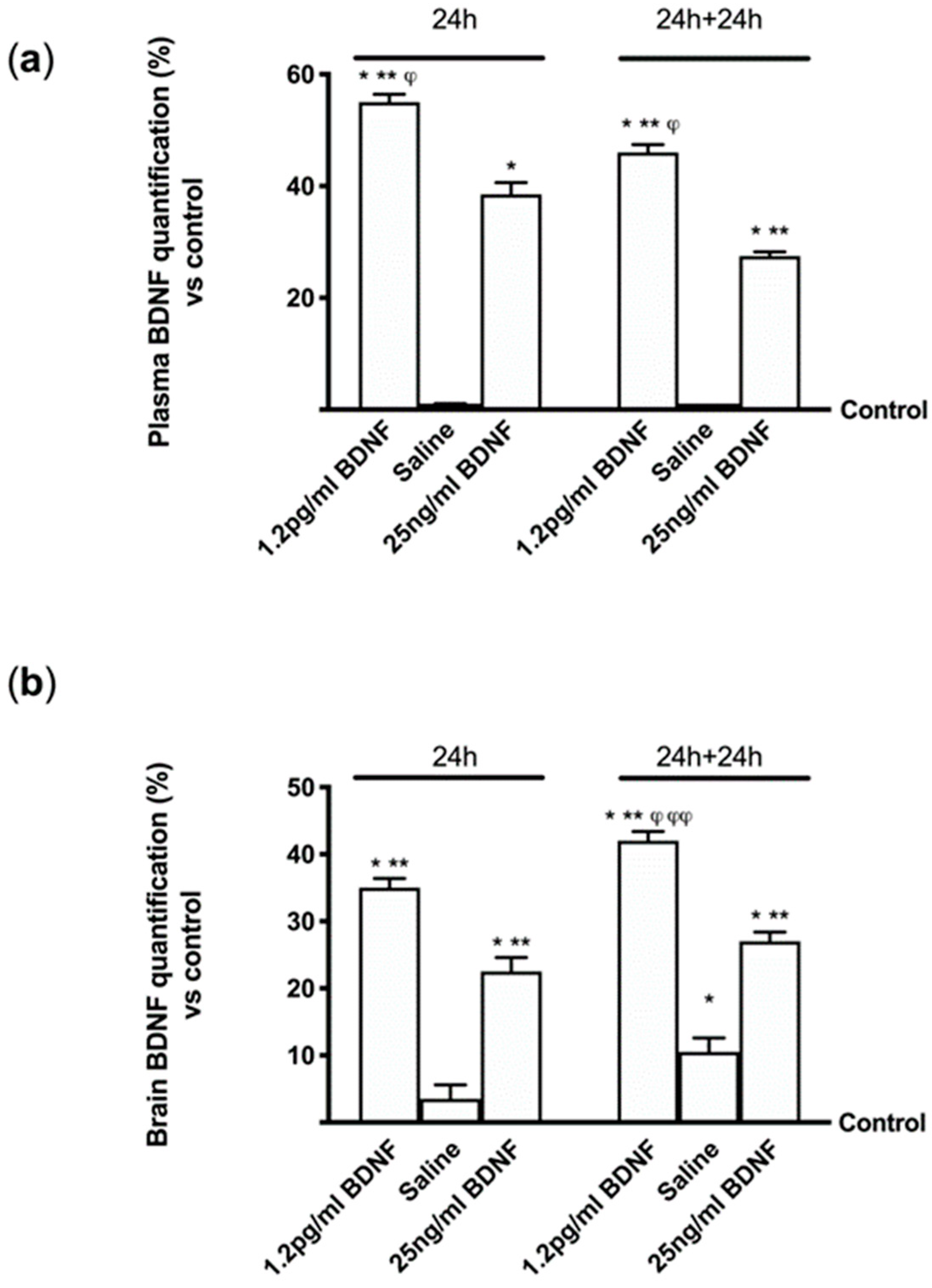
3.7. Analysis of Bioavailability of BDNF Solutions and Their Effects in Mouse Brain
3.8. Effects of BDNF Solutions in Mouse Brain During Time
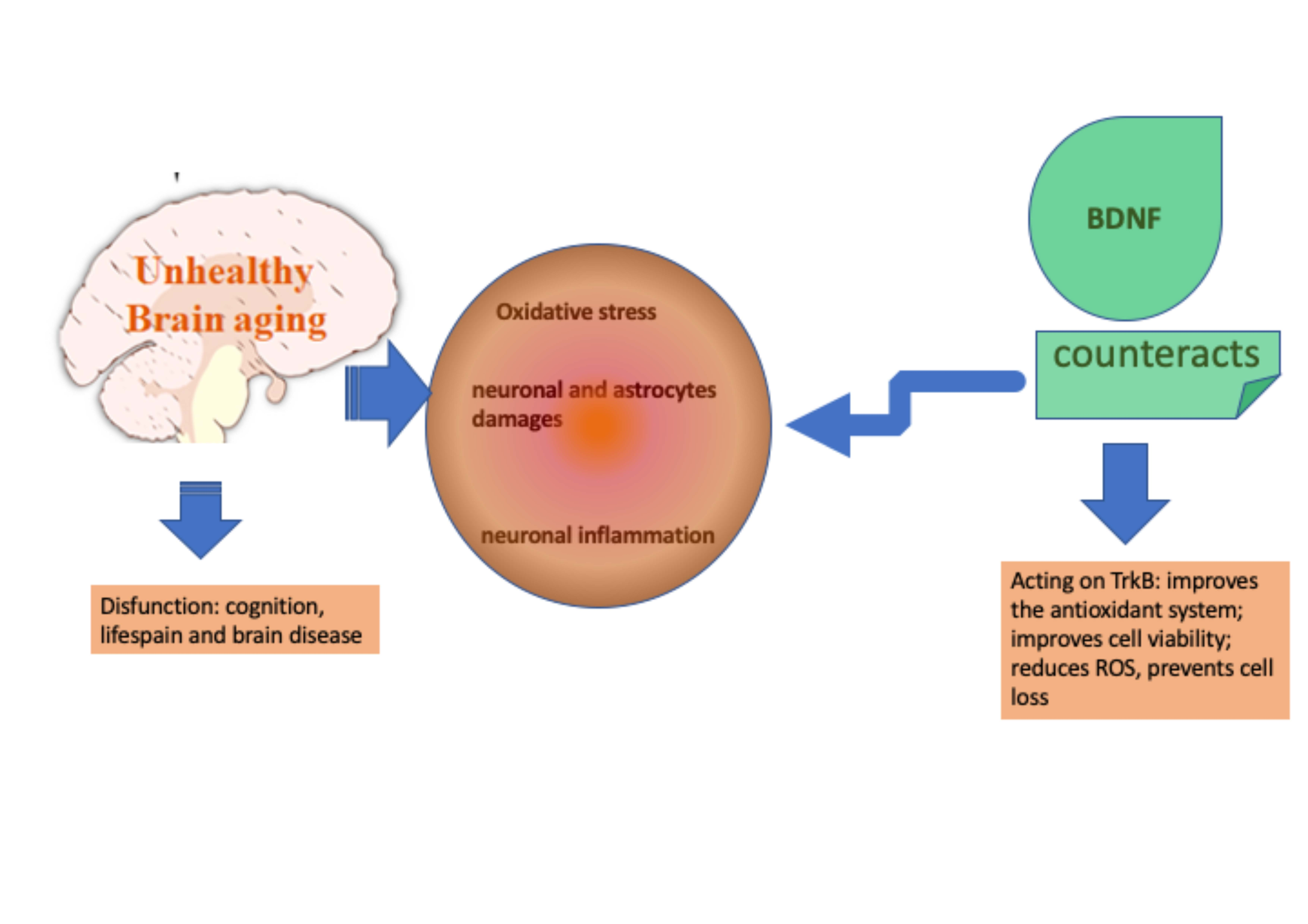
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Method: Key Phases of Rissole Preparation

Appendix B

References
- Anderson-Hanley, C.; Barcelos, N.M.; Zimmerman, E.A.; Gillen, R.W.; Dunnam, M.; Cohen, B.D.; Yerokhin, V.; Miller, K.E.; Hayes, D.J.; Arciero, P.J.; et al. The Aerobic and Cognitive Exercise Study (ACES) for Community-Dwelling Older Adults with or At-Risk for Mild Cognitive Impairment (MCI): Neuropsychological, Neurobiological and Neuroimaging Outcomes of a Randomized Clinical Trial. Front. Aging Neurosci. 2018, 10, 76. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.-M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2012, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, E.G.; Xu, B. New insights into the role of brain-derived neurotrophic factor in synaptic plasticity. Mol. Cell. Neurosci. 2009, 42, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silhol, M.; Arancibia, S.; Perrin, D.; Maurice, T.; Alliot, J.; Tapia-Arancibia, L. Effect of Aging on Brain-Derived Neurotrophic Factor, proBDNF, and Their Receptors in the Hippocampus of Lou/C Rats. Rejuvenation Res. 2008, 11, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Nockher, W.A.; Renz, H. Neurotrophins in inflammatory lung diseases: Modulators of cell differentiation and neuroimmune interactions. Cytokine Growth Factor Rev. 2003, 14, 559–578. [Google Scholar] [CrossRef]
- Maroder, M.; Bellavia, D.; Vacca, A.; Felli, M.P.; Screpanti, I. The thymus at the crossroad of neuroimmune interactions. Ann. N. Y. Acad. Sci. 2000, 917, 741–747. [Google Scholar] [CrossRef]
- Petzold, A.; Psotta, L.; Brigadski, T.; Endres, T.; Lessmann, V. Chronic BDNF deficiency leads to an age-dependent impairment in spatial learning. Neurobiol. Learn. Mem. 2015, 120, 52–60. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Almeida, R.D.; Manadas, B.; Melo, C.V.; Gomes, J.R.; Mendes, C.S.; Grãos, M.; Carvalho, R.F.; Carvalho, A.P.; Duarte, C.B. Neuroprotection by BDNF against glutamate-induced apoptotic cell death is mediated by ERK and PI3-kinase pathways. Cell Death Differ. 2005, 12, 1329–1343. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Michaelis, E.K. Selective Neuronal Vulnerability to Oxidative Stress in the Brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Munshi, S.; Banerjee, K.; Thakurta, I.G.; Sinha, M.; Bagh, M.B. Mitochondrial Dysfunction during Brain Aging: Role of Oxidative Stress and Modulation by Antioxidant Supplementation. Aging Dis. 2011, 2, 242–256. [Google Scholar] [PubMed]
- Mattson, M.P.; Magnus, T. Ageing and neuronal vulnerability. Nat. Rev. Neurosci. 2006, 7, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Karege, F.; Perret, G.; Bondolfi, G.; Schwald, M.; Bertschy, G.; Aubry, J.-M. Decreased serum brain-derived neurotrophic factor levels in major depressed patients. Psychiatry Res. Neuroimaging 2002, 109, 143–148. [Google Scholar] [CrossRef]
- Toyooka, K.; Asama, K.; Watanabe, Y.; Muratake, T.; Takahashi, M.; Someya, T.; Nawa, H. Decreased levels of brain-derived neurotrophic factor in serum of chronic schizophrenic patients. Psychiatry Res. Neuroimaging 2002, 110, 249–257. [Google Scholar] [CrossRef]
- Tapia-Arancibia, L.; Aliaga, E.; Silhol, M.; Arancibia, S. New insights into brain BDNF function in normal aging and Alzheimer disease. Brain Res. Rev. 2008, 59, 201–220. [Google Scholar] [CrossRef]
- Knusel, B.; Beck, K.D.; Winslow, J.W.; Rosenthal, A.; Burton, L.E.; Widmer, H.R. Brain-derived neurotrophic factor administration protects basal fore-brain cholinergic but not nigral dopaminergic neurons from degenerative changes after axotomy in the adult rat brain. J. Neurosci. 1992, 12, 4391–4402. [Google Scholar] [CrossRef]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2010, 14, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, F.; Racagni, G.; Riva, M.A. The expanding role of BDNF: A therapeutic target for Alzheimer’s disease? Pharmacogenom. J. 2006, 6, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Thoenen, H.; Sendtner, M. Neurotrophins: From enthusiastic expectations through sobering experiences to rational therapeutic approaches. Nat. Neurosci. 2002, 5, 1046–1050. [Google Scholar] [CrossRef]
- Gariboldi, S.; Palazzo, M.; Zanobbio, L.; Dusio, G.F.; Mauro, V.; Solimene, U.; Cardani, D.; Mantovani, M.; Rumio, C. Low dose oral administration of cytokines for treatment of allergic asthma. Pulm. Pharmacol. Ther. 2009, 22, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Uberti, F.; Morsanuto, V.; Ghirlanda, S.; Ruga, S.; Clemente, N.; Boieri, C.; Boldorini, R.; Molinari, C. Highly Diluted Acetylcholine Promotes Wound Repair in an In Vivo Model. Adv. Wound Care 2018, 7, 121–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brod, S.A.; Khan, M. Oral Administration of IFN-α is Superior to Subcutaneous Administration of IFN-α in the Suppression of Chronic Relapsing Experimental Autoimmune Encephalomyelitis. J. Autoimmun. 1996, 9, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Saba, J.; Turati, J.; Ramírez, D.; Carniglia, L.; Durand, D.; Lasaga, M.; Caruso, C. Astrocyte truncated tropomyosin receptor kinase B mediates brain-derived neurotrophic factor anti-apoptotic effect leading to neuroprotection. J. Neurochem. 2018, 146, 686–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Zhang, Y.; Wang, X.; Ma, H. Exogenous Brain-Derived Neurotrophic Factor at a 50 ng/mL Concentration has a Significant Protective Effect on Bilirubin-Induced Cerebral Cortex Neuronal Injury. Clin. Lab. 2017, 63, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Schildge, S.; Bohrer, C.; Beck, K.; Schachtrup, K. Isolation and Culture of Mouse Cortical Astrocytes. J. Vis. Exp. 2013, 71, 50079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Magrané, J. Isolation and Culture of Neurons and Astrocytes from the Mouse Brain Cortex. Adv. Struct. Saf. Stud. 2011, 793, 63–75. [Google Scholar] [CrossRef]
- Thomaz, A.; Jaeger, M.; Buendia, M.; Bambini-Junior, V.; Gregianin, L.J.; Brunetto, A.L.; Brunetto, A.T.; De Farias, C.B.; Roesler, R. BDNF/TrkB Signaling as a Potential Novel Target in Pediatric Brain Tumors: Anticancer Activity of Selective TrkB Inhibition in Medulloblastoma Cells. J. Mol. Neurosci. 2015, 59, 326–333. [Google Scholar] [CrossRef]
- Lü, L.; Li, J.; Zhu, Y.; Mak, Y.T.; Yew, D.T. H2O2-Induced Changes in Astrocytic Cultures from Control and Rapidly Aging Strains of Mouse. Int. J. Neurosci. 2008, 118, 1239–1250. [Google Scholar] [CrossRef]
- Uberti, F.; Morsanuto, V.; Ghirlanda, S.; Molinari, C. Iron Absorption from Three Commercially Available Supplements in Gastrointestinal Cell Lines. Nutrients 2017, 9, 1008. [Google Scholar] [CrossRef] [Green Version]
- DiMarco, R.L.; Hunt, D.R.; Dewi, R.E.; Heilshorn, S.C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials 2017, 129, 152–162. [Google Scholar] [CrossRef]
- Obringer, C.; Manwaring, J.; Goebel, C.; Hewitt, N.J.; Rothe, H. Suitability of the in vitro Caco-2 assay to predict the oral absorption of aromatic amine hair dyes. Toxicol. Vitr. 2016, 32, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 1, 175–185. [Google Scholar] [CrossRef]
- Zorkina, Y.A.; Volgina, N.E.; Gorlachev, G.; Mel’Nikov, P.A.; Golanov, A.V.; Potapov, A.A.; Chekhonin, V.P. Effect of γ-Irradiation on Expression of Tight and Adherens Junction Protein mRNA on In Vitro Blood–Brain Barrier Model. Bull. Exp. Boil. Med. 2014, 158, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Kulczar, C.; Lubin, K.E.; Lefebvre, S.; Miller, N.W.; Knipp, G. Development of a direct contact astrocyte-human cerebral microvessel endothelial cells blood-brain barrier coculture model. J. Pharm. Pharmacol. 2017, 69, 1684–1696. [Google Scholar] [CrossRef]
- Uberti, F.; Lattuada, D.; Morsanuto, V.; Nava, U.; Bolis, G.; Vacca, G.; Squarzanti, D.F.; Cisari, C.; Molinari, C. Vitamin D Protects Human Endothelial Cells from Oxidative Stress Through the Autophagic and Survival Pathways. J. Clin. Endocrinol. Metab. 2014, 99, 1367–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uberti, F.; Morsanuto, V.; Aprile, S.; Ghirlanda, S.; Stoppa, I.; Cochis, A.; Grosa, G.; Rimondini, L.; Molinari, C. Biological effects of combined resveratrol and vitamin D3 on ovarian tissue. J. Ovarian Res. 2017, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Cappellano, G.; Uberti, F.; Caimmi, P.P.; Pietronave, S.; Mary, D.A.; Dianzani, C.; Micalizzi, E.; Melensi, M.; Boldorini, R.; Nicosia, G.; et al. Different Expression and Function of the Endocannabinoid System in Human Epicardial Adipose Tissue in Relation to Heart Disease. Can. J. Cardiol. 2013, 29, 499–509. [Google Scholar] [CrossRef]
- Uberti, F.; Bardelli, C.; Morsanuto, V.; Ghirlanda, S.; Cochis, A.; Molinari, C. Stimulation of the Nonneuronal Cholinergic System by Highly Diluted Acetylcholine in Keratinocytes. Cells Tissues Organs 2016, 203, 215–230. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Walker, M.K.; Boberg, J.R.; Walsh, M.T.; Wolf, V.; Trujillo, A.; Duke, M.S.; Palme, R.; Felton, L.A. A less stressful alternative to oral gavage for pharmacological and toxicological studies in mice. Toxicol. Appl. Pharmacol. 2012, 260, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Bachmanov, A.; Reed, D.; Beauchamp, G.K.; Tordoff, M. Food intake, water intake, and drinking spout side preference of 28 mouse strains. Behav. Genet. 2002, 32, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. Voluntary oral administration of drugs in mice. Protoc. Exch. 2011. [Google Scholar] [CrossRef]
- Kaushal, N.; Nair, D.; Gozal, D.; Ramesh, V. Socially isolated mice exhibit a blunted homeostatic sleep response to acute sleep deprivation compared to socially paired mice. Brain Res. 2012, 1454, 65–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.; Moya-Alvarado, G.; Gonzalez-Billaut, C.; Bronfman, F.C. Cellular and molecular mechanisms regulating neuronal growth by brain-derived neurotrophic factor. Cytoskeleton (Hoboken) 2016, 73, 612–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budni, J.; Bellettini-Santos, T.; Mina, F.; Garcez, M.L.; Zugno, A.I. The involvement of BDNF, NGF and GDNF in aging and Alzheimer’s disease. Aging Dis. 2015, 6, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, T. BDNF Integrity in Ageing and Stress. MOJ Gerontol. Geriatr. 2017, 1, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, A.H.; Tuszynski, M.H. Potential therapeutic uses of BDNF in neurological and psychiatric disorders. Nat. Rev. Drug Discov. 2011, 10, 209–219. [Google Scholar] [CrossRef]
- Cummins, J.M.; Krakowka, G.S.; Thompson, C.G. Systemic effects of interferons after oral administration in animals and humans. Am. J. Veter Res. 2005, 66, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Tompkins, W.A. Immunomodulation and Therapeutic Effects of the Oral Use of Interferon-alpha: Mechanism of Action. J. Interf. Cytokine Res. 1999, 19, 817–828. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; Saal, F.S.V.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Biancotto, A.; Wank, A.; Perl, S.; Cook, W.; Olnes, M.J.; Dagur, P.K.; Fuchs, J.C.; Langweiler, M.; Wang, E.; McCoy, J.P. Baseline levels and temporal stability of 27 multiplexed serum cytokine concentrations in healthy subjects. PLoS ONE 2013, 8, e76091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinta, S.J.; Woods, G.; Rane, A.; DeMaria, M.; Campisi, J.; Andersen, J.K. Cellular senescence and the aging brain. Exp. Gerontol. 2014, 68, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-León, Y.; Pascual, A. Regulation of beta-amyloid precursor protein expression by brain-derived neurotrophic factor involves activation of both the Ras and phosphatidylinositide 3-kinase signalling pathways. J. Neurochem. 2004, 88, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-León, Y.; Pascual, A. Induction of tyrosine kinase receptor b by retinoic acid allows brain-derived neurotrophic factor-induced amyloid precursor protein gene expression in human sh-sy5y neuroblastoma cells. Neuroscience 2003, 120, 1019–1026. [Google Scholar] [CrossRef] [Green Version]
- Basak, J.M.; Verghese, P.B.; Yoon, H.; Kim, J.; Holtzman, D.M. Low-density Lipoprotein Receptor Represents an Apolipoprotein E-independent Pathway of Aβ Uptake and Degradation by Astrocytes*. J. Boil. Chem. 2012, 287, 13959–13971. [Google Scholar] [CrossRef] [Green Version]
- Belinson, H.; Lev, D.; Masliah, E.; Michaelson, D.M. Activation of the amyloid cascade in apolipoprotein E4 transgenic mice induces lysosomal activation and neurodegeneration resulting in marked cognitive deficits. J. Neurosci. 2008, 28, 4690–4701. [Google Scholar] [CrossRef] [Green Version]
- Manelli, A.M.; Bulfinch, L.C.; Sullivan, P.M.; Ladu, M.J. Abeta42 neurotoxicity in primary co-cultures: Effect of apoE isoform and Abeta conformation. Neurobiol. Aging 2006, 28, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Herskovits, A.Z.; Guarente, L. SIRT1 in neurodevelopment and brain senescence. Neuron 2014, 81, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Donmez, G.; Outeiro, T.F. SIRT1 and SIRT2: Emerging targets in neurodegeneration. EMBO Mol. Med. 2013, 5, 344–352. [Google Scholar] [CrossRef]
- De Luca, C.; Colangelo, A.M.; Alberghina, L.; Papa, M. Neuro-Immune Hemostasis: Homeostasis and Diseases in the Central Nervous System. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, H. Weber’s Law. In Encyclopaedia Britannica, 11th ed.; Cambridge University Press: Cambridge, UK, 1911. [Google Scholar]
- Droge, W. Oxidative stress and aging. Adv. Exp. Med. Biol. 2003, 543, 191–200. [Google Scholar] [PubMed]
- Jovanović, S.; Jovanović, S. Toxicity induced by cumene hydroperoxide in leech Retzius nerve cells: The protective role of glutathione. Folia Boil. 2013, 61, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, Oxidants, and Aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mufson, E.J.; Mahady, L.; Waters, D.; Counts, S.E.; Perez, S.E.; DeKosky, S.T.; Ginsberg, S.D.; Ikonomovic, M.D.; Scheff, S.; Binder, L. Hippocampal plasticity during the progression of Alzheimer’s disease. Neuroscience 2015, 309, 51–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siuda, J.; Patalong-Ogiewa, M.; Żmuda, W.; Targosz-Gajniak, M.; Niewiadomska, E.; Matuszek, I.; Jędrzejowska-Szypułka, H.; Rudzińska-Bar, M. Corrigendum to “Cognitive impairment and BDNF serum levels” [Polish, J. Neurol. Neurosurg. 51 (2017) 24–32]. Neurol. I Neurochir. Polska 2017, 51, 537. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.; Alonso, M.; Bramham, C.R.; Pozzo-Miller, L. From acquisition to consolidation: On the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn. Mem. 2002, 9, 224–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khakh, B.S.; Sofroniew, M. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stimulations | 30 min | 1 h | 3 h | 4 h | 5 h | 6 h |
|---|---|---|---|---|---|---|
| 1 pg/mL BDNF SKA | 2.91 ± 0.3 | 4.18 ± 0.1 | 3.36 ± 0.2 | 3.27 ± 0.3 | 2.02 ± 0.2 | 1.97 ± 0.3 |
| Saline | 0.11 ± 0.1 | 0.29 ± 0.1 | 0.36 ± 0.1 | 0.5 ± 0.1 | 0.57 ± 0.1 | 0.6 ± 0.1 |
| 50 ng/mL BDNF | 3 ± 0.3 | 3.98 ± 0.3 | 4 ± 0.3 | 3.2 ± 0.2 | 1.98 ± 0.2 | 1.02 ± 0.1 |
| 1 pg/mL BDNF SKA | 32.75 | 39.8 | 36.5 | 34.85 | 26.6 | 26 |
| Saline | <20 | <20 | <20 | <20 | <20 | <20 |
| 50 ng/mL BDNF | 33.2 | 38.15 | 39.8 | 33.2 | 25.8 | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinari, C.; Morsanuto, V.; Ruga, S.; Notte, F.; Farghali, M.; Galla, R.; Uberti, F. The Role of BDNF on Aging-Modulation Markers. Brain Sci. 2020, 10, 285. https://doi.org/10.3390/brainsci10050285
Molinari C, Morsanuto V, Ruga S, Notte F, Farghali M, Galla R, Uberti F. The Role of BDNF on Aging-Modulation Markers. Brain Sciences. 2020; 10(5):285. https://doi.org/10.3390/brainsci10050285
Chicago/Turabian StyleMolinari, Claudio, Vera Morsanuto, Sara Ruga, Felice Notte, Mahitab Farghali, Rebecca Galla, and Francesca Uberti. 2020. "The Role of BDNF on Aging-Modulation Markers" Brain Sciences 10, no. 5: 285. https://doi.org/10.3390/brainsci10050285
APA StyleMolinari, C., Morsanuto, V., Ruga, S., Notte, F., Farghali, M., Galla, R., & Uberti, F. (2020). The Role of BDNF on Aging-Modulation Markers. Brain Sciences, 10(5), 285. https://doi.org/10.3390/brainsci10050285