Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions on Corticospinal Excitability to Forearm Muscles during Low-Intensity Hand-Gripping



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
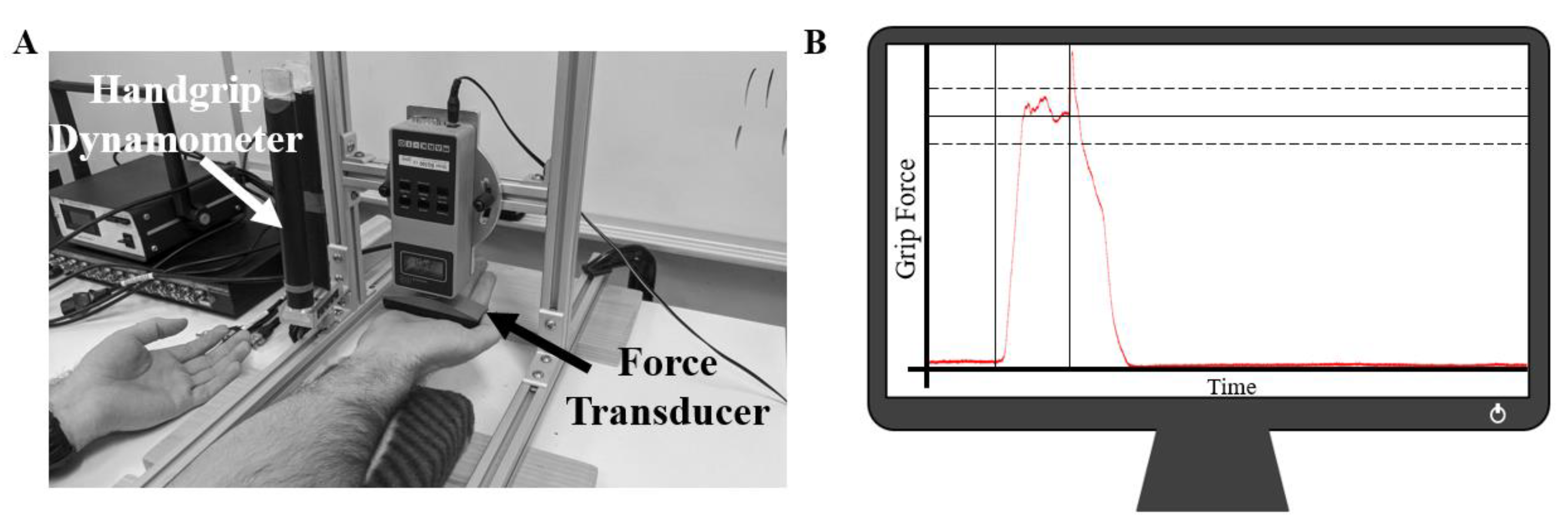
2.2. Experimental Setup
2.3. Electromyography
2.4. TMS
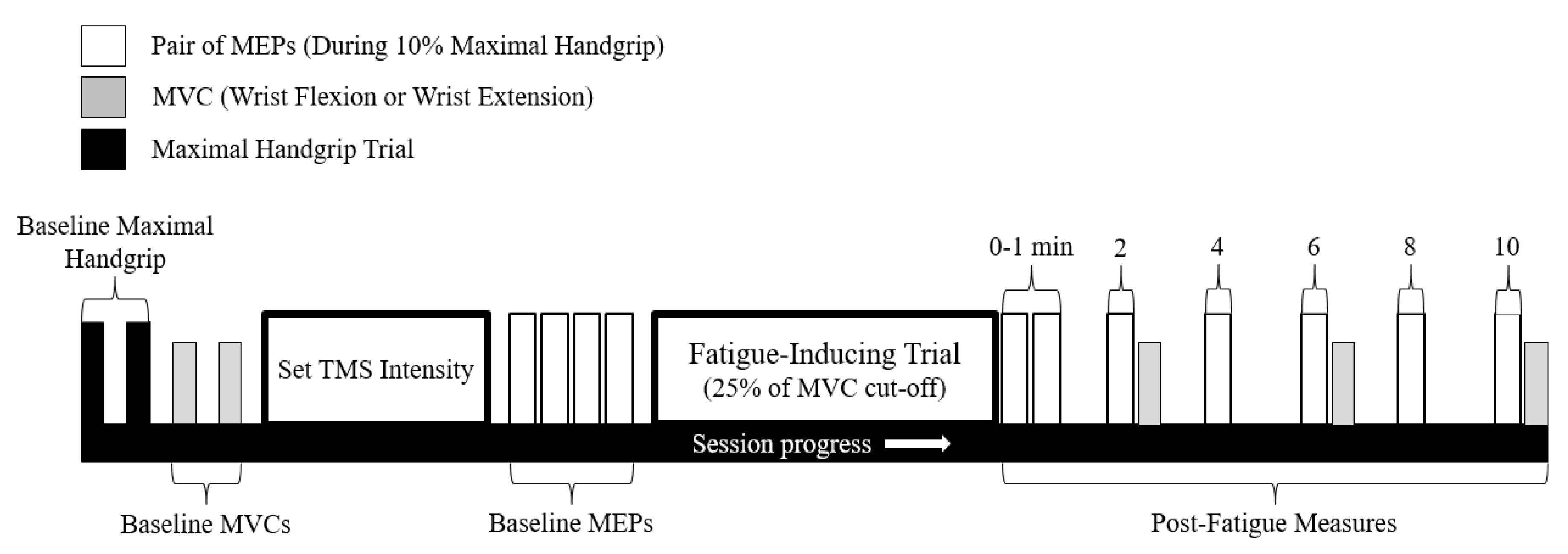
2.5. Experimental Protocol
2.6. Data Analysis
2.7. Statistics
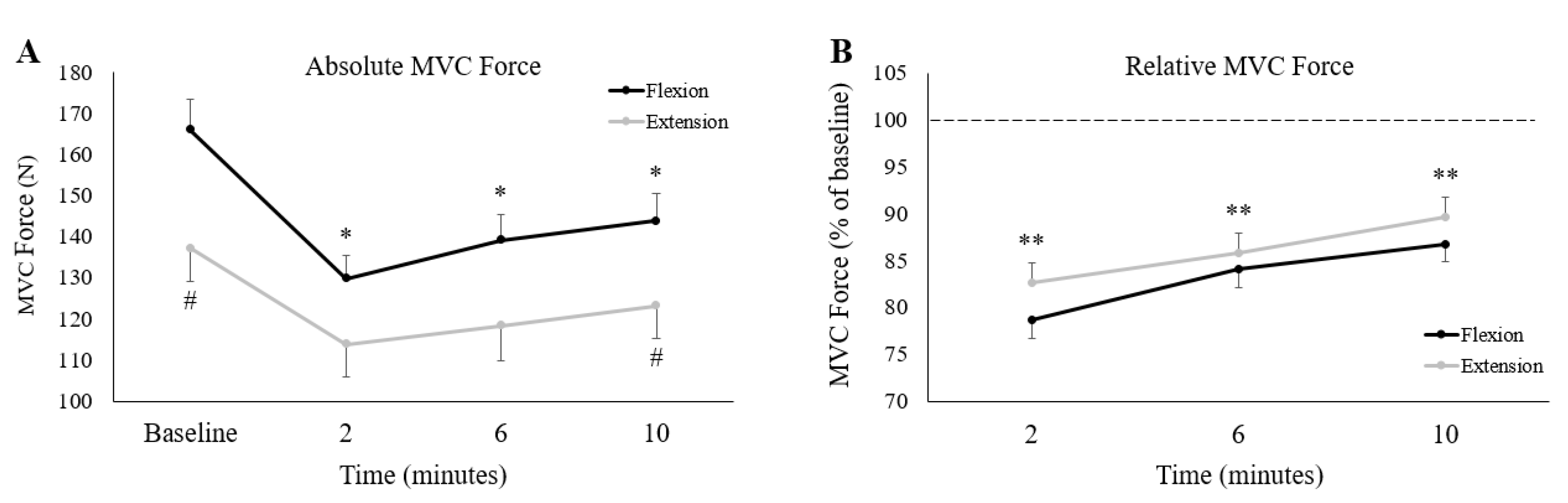
3. Results
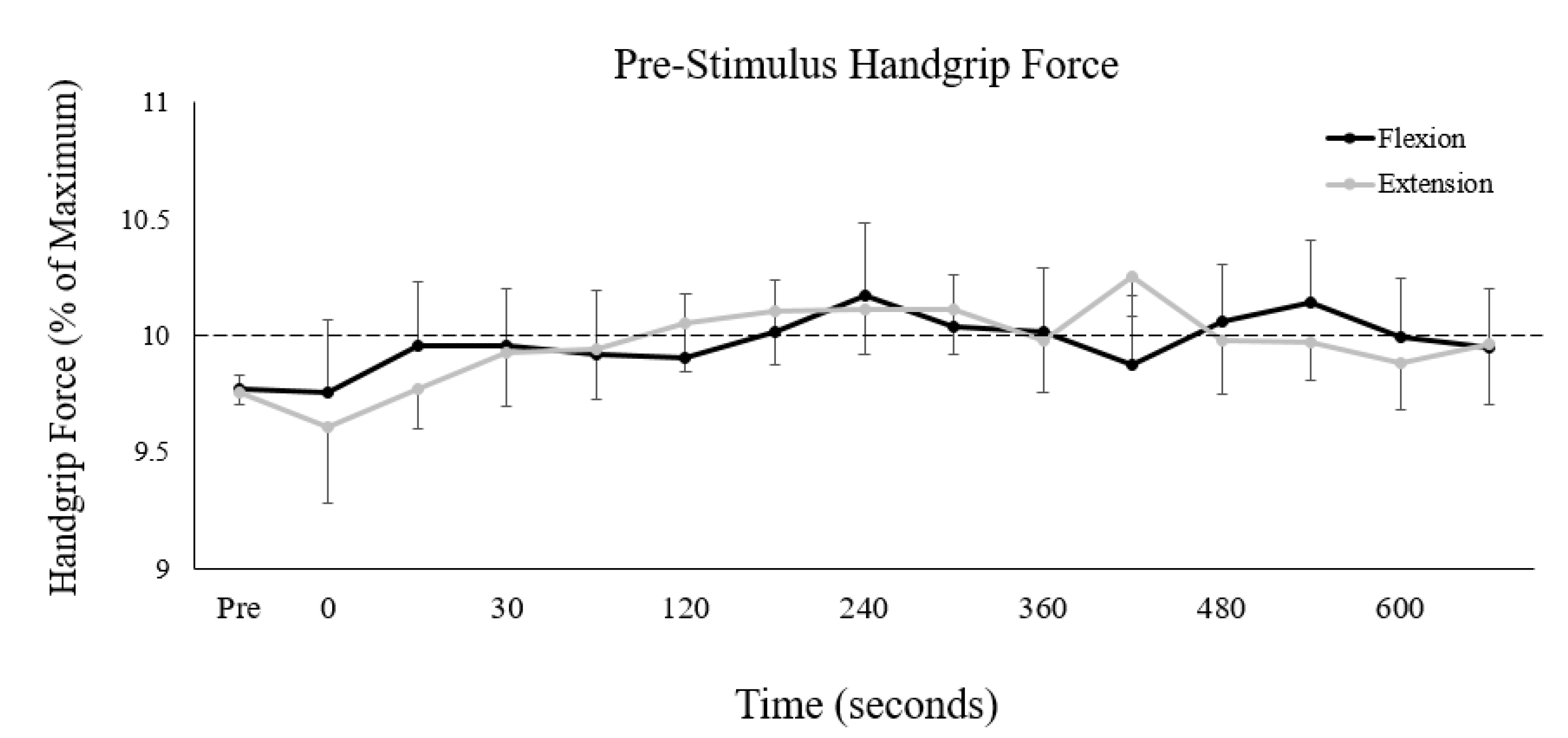
3.1. Pre-Stimulus Handgrip Force
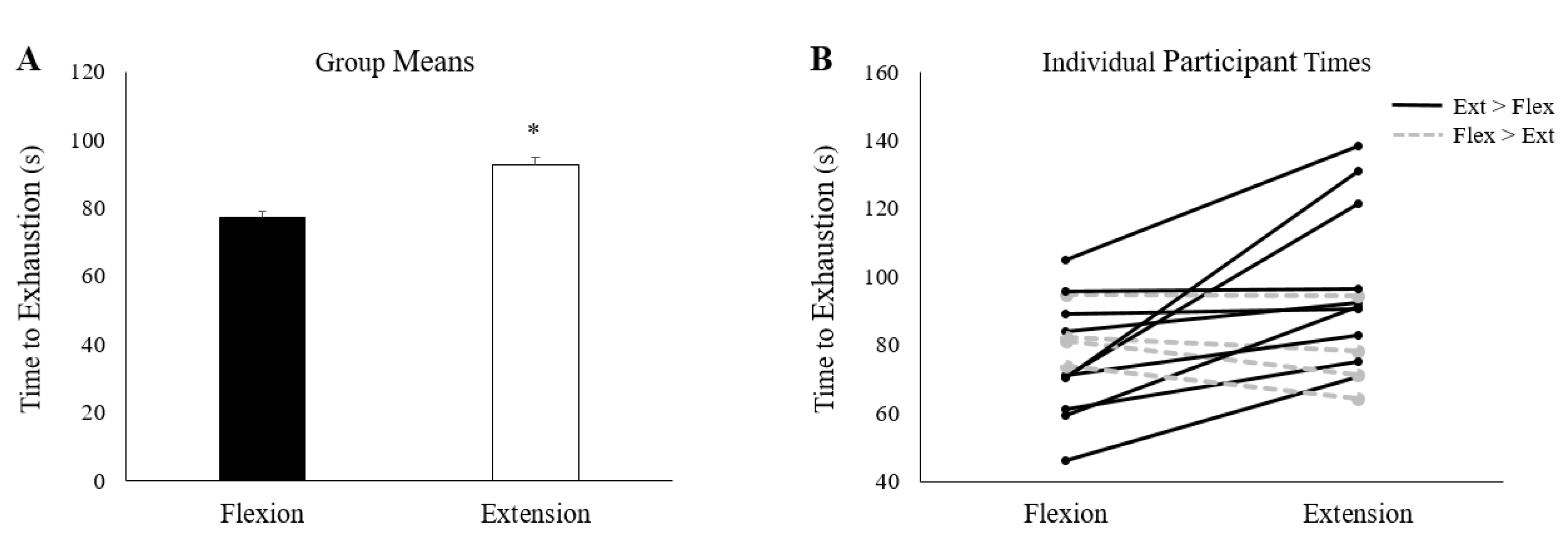
3.2. Fatigue Inducing Trial
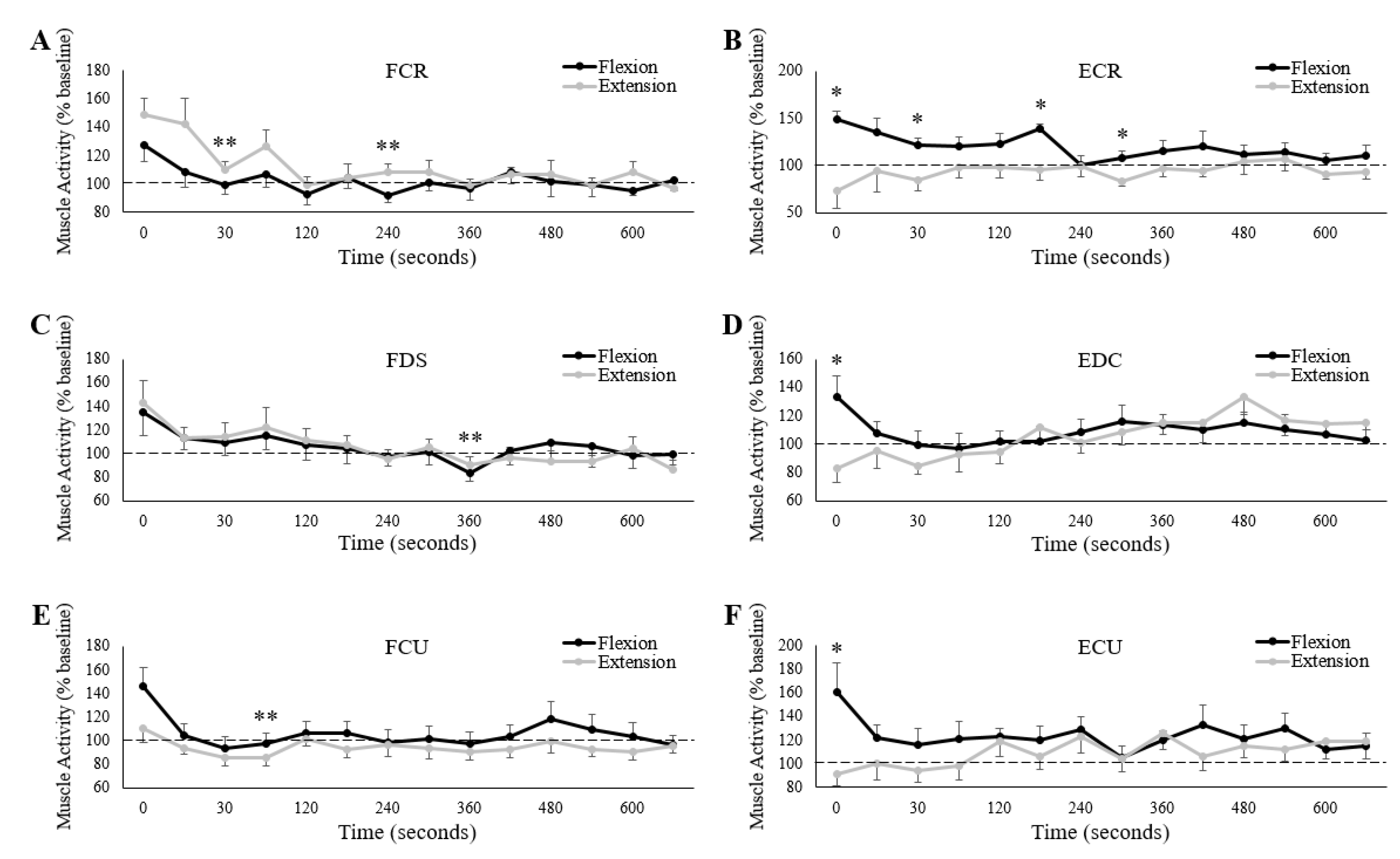
3.3. Muscle Activity
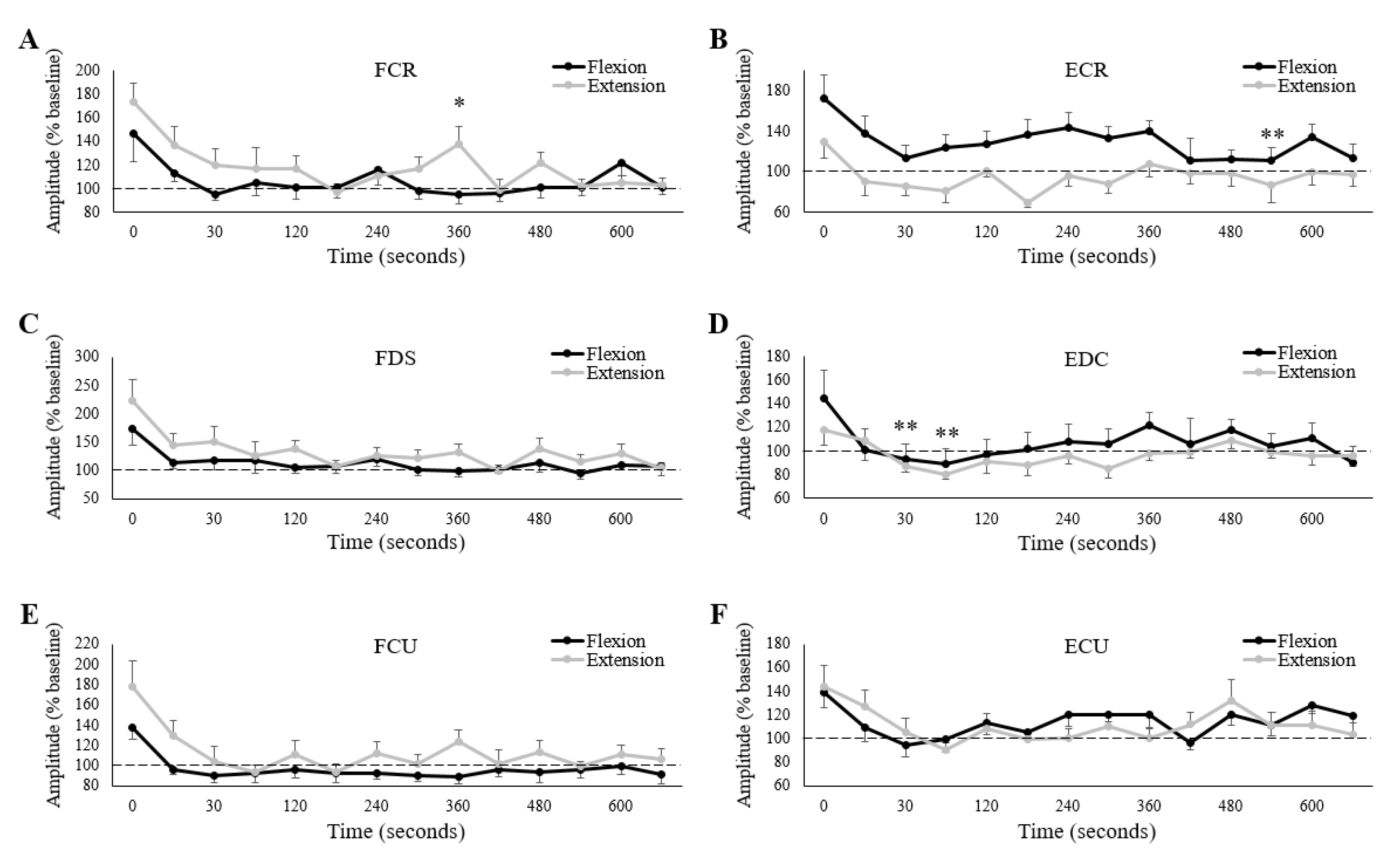
3.4. Corticospinal Excitability
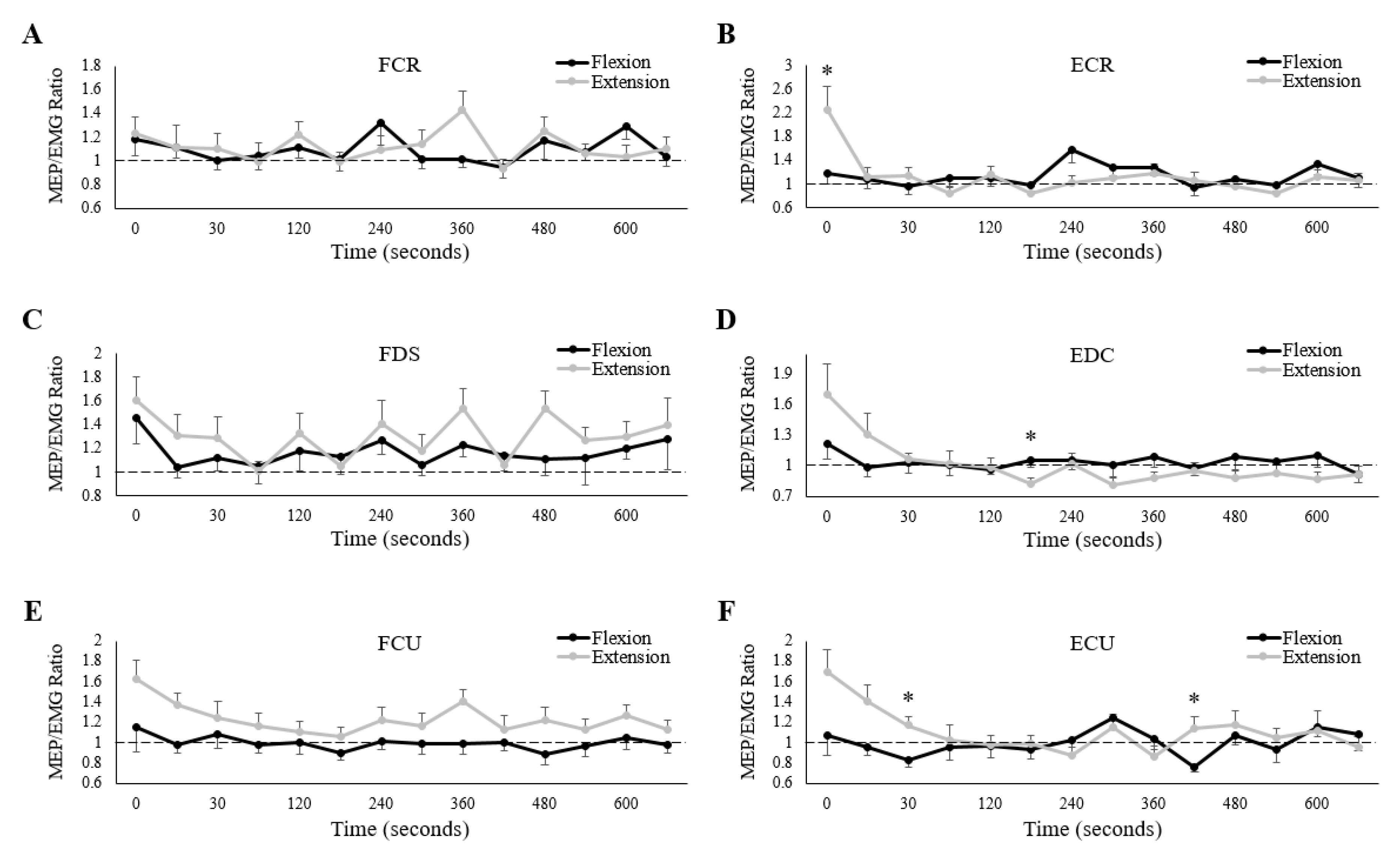
3.5. MEP/EMG Ratios
4. Discussion
4.1. Muscle Activity
- (1)
- Fatigue specificity: Fatigue was induced by sustained wrist flexion or extension MVCs, but muscle activity was assessed while participants exerted 10% of their maximal handgrip force. While research has demonstrated that muscle activity and certain neurological measures can be state, intensity, and muscle dependent following fatigue [8,42,43], it is intuitive to suggest that they might be task-dependent as well. Not just in terms of the task used to induce performance fatigability but also in terms of the task in which measurements are conducted. For instance, it is possible that handgrip force as low as 10% of maximum can be produced mostly with intrinsic finger muscles–muscles that may not have been fully recruited during maximal wrist exertions. Thus, the muscles that were active during the handgrip task may not have been effectively fatigued during isolated wrist extension. Alternatively, to compensate for this post-fatigue decrease in extensor activity, contributions from other muscles (such as the extensor pollicis longus, which lies deep to the ECR and was not assessed) may have increased. Subsequent investigations utilizing indwelling EMG would add valuable insight to this possibility.
- (2)
- Metabolic optimization: Motor outputs are optimally executed when there is an appropriate balance of joint stability (greatest contribution to joint stability produced by muscle contraction) and metabolic expenditure [44,45]. Prior to fatigue, the level of wrist extensor activity in the present study was theoretically optimal in magnitude and energy expenditure to counter the forces produced by the flexors. However, following wrist extension fatigue, not only would greater motor unit recruitment of the extensors have been needed to exert the same level of co-contraction (since motoneuron discharge rates were likely reduced), but available energy reserves would have also been reduced. Thus, exerting similar baseline forces would cost more energy in a moment of reduced availability. It is therefore possible that wrist joint stability, provided by the wrist extensors, decreased in favour of energy expenditure. While support for this possibility is scarce, some studies have shown that co-contraction [46], limb impedance [47], and joint stiffness [48] all decrease following fatigue. It should be noted that these studies were all conducted during dynamic reaching, not isometric conditions. However, antagonist muscle activity also increases less post-fatigue than agonist activity during isometric actions of the torso [49].
- (3)
- Forearm co-contraction: The suggestions raised above were likely present following the wrist flexion session as well. Thus, it is unclear why extensor muscle activity only increased following the wrist flexion session. Since the wrist flexor muscles demonstrate little activity during isolated wrist extension [11], sustained wrist extension may have only induced fatigue in the wrist extensor muscles. Thus, a feasible reduction in extensor co-contraction (for metabolic purposes) may have been compensated for by other, non-fatigued muscles. In contrast, the wrist extensors are highly active during isolated wrist flexion [11], meaning that performance fatigability was likely induced in the entire forearm following sustained wrist flexion. If so, any reduction in wrist extensor co-contraction might have adversely decreased wrist joint stability. As other forearm muscles were also likely fatigued, and unable to compensate, wrist extensor co-contraction may have increased out of necessity. Thus, muscle activity in all three extensors was higher following sustained wrist flexion than sustained wrist extension.
4.2. Corticospinal Excitability
4.3. Additional Mechanisms
4.4. Methodological Considerations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kluger, B.M.; Krupp, L.B.; Enoka, R.M. Fatigue and fatigability in neurologic illnesses: Proposal for a unified taxonomy. Neurology 2013, 80, 409–416. [Google Scholar] [CrossRef]
- Enoka, R.M.; Duchateau, J. Translating Fatigue to Human Performance. Med. Sci. Sports Exerc. 2016, 48, 2228–2238. [Google Scholar] [CrossRef] [Green Version]
- Marrelli, K.; Cheng, A.J.; Brophy, J.D.; Power, G.A. Perceived Versus Performance Fatigability in Patients with Rheumatoid Arthritis. Front. Physiol. 2018, 9, 1395. [Google Scholar] [CrossRef]
- Löscher, W.N.; Cresswell, A.G.; Thorstensson, A. Excitatory drive to the alpha-motoneuron pool during a fatiguing submaximal contraction in man. J. Physiol. 1996, 491, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Löscher, W.N.; Cresswell, A.G.; Thorstensson, A. Central fatigue during a long-lasting submaximal contraction of the triceps surae. Exp. Brain Res. 1996, 108, 305–314. [Google Scholar] [CrossRef]
- Zijdewind, I.; Zwarts, M.J.; Kernell, D. Influence of a voluntary fatigue test on the contralateral homologous muscle in humans? Neurosci. Lett. 1998, 253, 41–44. [Google Scholar] [CrossRef]
- De Haan, A.; Jones, D.A.; Sargeant, A.J. Changes in velocity of shortening, power output and relaxation rate during fatigue of rat medial gastrocnemius muscle. Pflug. Arch. 1989, 413, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Gandevia, S.C. Spinal and Supraspinal Factors in Human Muscle Fatigue. Physiol. Rev. 2001, 81, 1725–1789. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.A.; Forman, G.N.; Mugnosso, M.; Zenzeri, J.; Murphy, B.; Holmes, M.W.R. Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions Similarly Impair Hand-Tracking Accuracy in Young Adults Using a Wrist Robot. Front. Sports Act. Living 2020, 2, 53. [Google Scholar] [CrossRef]
- Holmes, M.W.R.; Tat, J.; Keir, P.J. Neuromechanical control of the forearm muscles during gripping with sudden flexion and extension wrist perturbations. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 1826–1834. [Google Scholar] [CrossRef]
- Forman, D.A.; Forman, G.N.; Robathan, J.; Holmes, M.W.R. The influence of simultaneous handgrip and wrist force on forearm muscle activity. J. Electromyogr. Kinesiol. 2019, 45, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Lieber, R.L.; Fazeli, B.M.; Botte, M.J. Architecture of selected wrist flexor and extensor muscles. J. Hand Surg. Am. 1990, 15, 244–250. [Google Scholar] [CrossRef]
- Lieber, R.L.; Jacobson, M.D.; Fazeli, B.M.; Abrams, R.A.; Botte, M.J. Architecture of selected muscles of the arm and forearm: Anatomy and implications for tendon transfer. J. Hand Surg. Am. 1992, 17, 787–798. [Google Scholar] [CrossRef]
- Jacobson, M.D.; Raab, R.; Fazeli, B.M.; Abrams, R.A.; Botte, M.J.; Lieber, R.L. Architectural design of the human intrinsic hand muscles. J. Hand Surg. Am. 1992, 17, 804–809. [Google Scholar] [CrossRef]
- Gonzalez, R.V.; Buchanan, T.S.; Delp, S.L. How muscle architecture and moment arms affect wrist flexion-extension moments. J. Biomech. 1997, 30, 705–712. [Google Scholar] [CrossRef]
- Bawa, P.; Chalmers, G.R.; Jones, K.E.; Søgaard, K.; Walsh, M.L. Control of the wrist joint in humans. Eur. J. Appl. Physiol. 2000, 83, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Mogk, J.; Keir, P. The effects of posture on forearm muscle loading during gripping. Ergonomics 2003, 46, 956–975. [Google Scholar] [CrossRef]
- Hägg, G.; Milerad, E. Forearm extensor and flexor muscle exertion during simulated gripping work—An electromyographic study. Clin. Biomech. 1997, 12, 39–43. [Google Scholar] [CrossRef]
- Marsden, C.D.; Meadows, J.C.; Merton, P.A. Isolated single motor units in human muscle and their rate of discharge during maximal voluntary effort. J. Physiol. 1971, 217, 12P–13P. [Google Scholar]
- Bellemare, F.; Woods, J.J.; Johansson, R.; Bigland-Ritchie, B. Motor-unit discharge rates in maximal voluntary contractions of three human muscles. J. Neurophysiol. 1983, 50, 1380–1392. [Google Scholar] [CrossRef]
- Person, R.S.; Kudina, L.P. Discharge frequency and discharge pattern of human motor units during voluntary contraction of muscle. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 471–483. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Cafarelli, E.; Vøllestad, N.K. Fatigue of submaximal static contractions. Acta Physiol. Scand. 1986, 556, 137–148. [Google Scholar]
- Edwards, R.G.; Lippold, O.C. The relation between force and integrated electrical activity in fatigued muscle. J. Physiol. 1956, 132, 677–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigland-Ritchie, B.; Donovan, E.F.; Roussos, C.S. Conduction velocity and EMG power spectrum changes in fatigue of sustained maximal efforts. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 51, 1300–1305. [Google Scholar] [CrossRef]
- Zanette, G.; Bonato, C.; Polo, A.; Tinazzi, M.; Manganotti, P.; Fiaschi, A. Long-lasting depression of motor-evoked potentials to transcranial magnetic stimulation following exercise. Exp. Brain Res. 1995, 107. [Google Scholar] [CrossRef]
- Samii, A.; Wassermann, E.M.; Ikoma, K.; Mercuri, B.; Hallett, M. Characterization of postexercise facilitation and depression of motor evoked potentials to transcranial magnetic stimulation. Neurology 1996, 46, 1376–1382. [Google Scholar] [CrossRef]
- Taylor, J.L.; Butler, J.E.; Allen, G.M.; Gandevia, S.C. Changes in motor cortical excitability during human muscle fatigue. J. Physiol. 1996, 490, 519–528. [Google Scholar] [CrossRef]
- Maruyama, A.; Matsunaga, K.; Tanaka, N.; Rothwell, J.C. Muscle fatigue decreases short-interval intracortical inhibition after exhaustive intermittent tasks. Clin. Neurophysiol. 2006, 117, 864–870. [Google Scholar] [CrossRef]
- Hunter, S.K.; McNeil, C.J.; Butler, J.E.; Gandevia, S.C.; Taylor, J.L. Short-interval cortical inhibition and intracortical facilitation during submaximal voluntary contractions changes with fatigue. Exp. Brain Res. 2016, 234, 2541–2551. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Forman, D.A.; Baarbé, J.; Daligadu, J.; Murphy, B.; Holmes, M.W.R. The effects of upper limb posture and a sub-maximal gripping task on corticospinal excitability to muscles of the forearm. J. Electromyogr. Kinesiol. 2016, 27, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Perotto, A.; Delagi, E.F. Anatomical Guide for the Electromyographer: The Limbs and Trunk, 5th ed.; Charles C. Thomas, Publisher, Ltd.: Springfield, IL, USA, 2011; ISBN 978-0-398-08648-0. [Google Scholar]
- Power, K.E.; Copithorne, D.B. Increased corticospinal excitability prior to arm cycling is due to enhanced supraspinal but not spinal motoneurone excitability. Appl. Physiol. Nutr. Metab. 2013, 38, 1154–1161. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.A.; Monks, M.; Power, K.E. Corticospinal excitability, assessed through stimulus response curves, is phase-, task-, and muscle-dependent during arm cycling. Neurosci. Lett. 2019, 692, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Vaseghi, B.; Zoghi, M.; Jaberzadeh, S. Inter-pulse Interval Affects the Size of Single-pulse TMS-induced Motor Evoked Potentials: A Reliability Study. Basic Clin. Neurosci. 2015, 6, 44–51. [Google Scholar]
- Aboodarda, S.J.; Copithorne, D.B.; Pearcey, G.E.P.; Button, D.C.; Power, K.E. Changes in supraspinal and spinal excitability of the biceps brachii following brief, non-fatiguing submaximal contractions of the elbow flexors in resistance-trained males. Neurosci. Lett. 2015, 607, 66–71. [Google Scholar] [CrossRef]
- Aboodarda, S.J.; Šambaher, N.; Millet, G.Y.; Behm, D.G. Knee extensors neuromuscular fatigue changes the corticospinal pathway excitability in biceps brachii muscle. Neuroscience 2017, 340, 477–486. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Johansson, R.; Lippold, O.C.; Smith, S.; Woods, J.J. Changes in motoneurone firing rates during sustained maximal voluntary contractions. J. Physiol. 1983, 340, 335–346. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Furbush, F.; Woods, J.J. Fatigue of intermittent submaximal voluntary contractions: Central and peripheral factors. J. Appl. Physiol. 1986, 61, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Enoka, R.M. Morphological features and activation patterns of motor units. J. Clin. Neurophysiol. 1995, 12, 538–559. [Google Scholar] [CrossRef]
- Sidhu, S.K.; Cresswell, A.G.; Carroll, T.J. Corticospinal Responses to Sustained Locomotor Exercises: Moving Beyond Single-Joint Studies of Central Fatigue. Sports Med. 2013, 43, 437–449. [Google Scholar] [CrossRef]
- Søgaard, K.; Gandevia, S.C.; Todd, G.; Petersen, N.T.; Taylor, J.L. The effect of sustained low-intensity contractions on supraspinal fatigue in human elbow flexor muscles. J. Physiol. 2006, 573, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; Gandevia, S.C. A comparison of central aspects of fatigue in submaximal and maximal voluntary contractions. J. Appl. Physiol. 2008, 104, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Hogan, N. Adaptive control of mechanical impedance by coactivation of antagonist muscles. IEEE Trans. Autom. Contr. 1984, 29, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Burdet, E.; Osu, R.; Franklin, D.W.; Milner, T.E.; Kawato, M. The central nervous system stabilizes unstable dynamics by learning optimal impedance. Nature 2001, 414, 446–449. [Google Scholar] [CrossRef]
- Missenard, O.; Mottet, D.; Perrey, S. The role of cocontraction in the impairment of movement accuracy with fatigue. Exp. Brain Res. 2008, 185, 151–156. [Google Scholar] [CrossRef]
- Selen, L.P.J.; Beek, P.J.; van Dieën, J.H. Fatigue-induced changes of impedance and performance in target tracking. Exp. Brain Res. 2007, 181, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Dutto, D.J.; Smith, G.A. Changes in spring-mass characteristics during treadmill running to exhaustion. Med. Sci. Sports Exerc. 2002, 34, 1324–1331. [Google Scholar] [CrossRef] [Green Version]
- Potvin, J.R.; O’Brien, P.R. Trunk muscle co-contraction increases during fatiguing, isometric, lateral bend exertions. Possible implications for spine stability. Spine 1998, 23, 774–780. [Google Scholar] [CrossRef]
- Liepert, J.; Kotterba, S.; Tegenthoff, M.; Malin, J.P. Central fatigue assessed by transcranial magnetic stimulation. Muscle Nerve 1996, 19, 1429–1434. [Google Scholar] [CrossRef]
- Kernell, D.; Monster, A.W. Motoneurone properties and motor fatigue. An intracellular study of gastrocnemius motoneurones of the cat. Exp. Brain Res. 1982, 46, 197–204. [Google Scholar] [CrossRef]
- Sawczuk, A.; Powers, R.K.; Binder, M.D. Intrinsic properties of motoneurons. Implications for muscle fatigue. Adv. Exp. Med. Biol. 1995, 384, 123–134. [Google Scholar] [PubMed]
- Ugawa, Y.; Terao, Y.; Hanajima, R.; Sakai, K.; Kanazawa, I. Facilitatory effect of tonic voluntary contraction on responses to motor cortex stimulation. Electroencephalogr. Clin. Neurophysiol. 1995, 97, 451–454. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Restuccia, D.; Oliviero, A.; Profice, P.; Ferrara, L.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.C. Effects of voluntary contraction on descending volleys evoked by transcranial stimulation in conscious humans. J. Physiol. 1998, 508, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Taylor, J.L.; Gandevia, S.C. Responses of Human Motoneurons to Corticospinal Stimulation during Maximal Voluntary Contractions and Ischemia. J. Neurosci. 2003, 23, 10224–10230. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.G.; Smith, J.L.; Butler, J.E.; Gandevia, S.C.; Taylor, J.L. Fatigue-sensitive afferents inhibit extensor but not flexor motoneurons in humans. J. Neurosci. 2006, 26, 4796–4802. [Google Scholar] [CrossRef] [Green Version]
- McNeil, C.J.; Martin, P.G.; Gandevia, S.C.; Taylor, J.L. The response to paired motor cortical stimuli is abolished at a spinal level during human muscle fatigue: Paired cortical stimuli in fatigue. J. Physiol. 2009, 587, 5601–5612. [Google Scholar] [CrossRef]
- McNeil, C.J.; Giesebrecht, S.; Gandevia, S.C.; Taylor, J.L. Behaviour of the motoneurone pool in a fatiguing submaximal contraction. J. Physiol. 2011, 589, 3533–3544. [Google Scholar] [CrossRef]
- Inghilleri, M.; Berardelli, A.; Cruccu, G.; Manfredi, M. Silent period evoked by transcranial stimulation of the human cortex and cervicomedullary junction. J. Physiol. 1993, 466, 521–534. [Google Scholar]
- Siebner, H.R.; Dressnandt, J.; Auer, C.; Conrad, B. Continuous intrathecal baclofen infusions induced a marked increase of the transcranially evoked silent period in a patient with generalized dystonia. Muscle Nerve 1998, 21, 1209–1212. [Google Scholar] [CrossRef]
- Hunter, S.K.; Todd, G.; Butler, J.E.; Gandevia, S.C.; Taylor, J.L. Recovery from supraspinal fatigue is slowed in old adults after fatiguing maximal isometric contractions. J. Appl. Physiol. 2008, 105, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Seifert, T.; Petersen, N.C. Changes in presumed motor cortical activity during fatiguing muscle contraction in humans. Acta Physiol. 2010, 199, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Tergau, F.; Geese, R.; Bauer, A.; Baur, S.; Paulus, W.; Reimers, C.D. Motor cortex fatigue in sports measured by transcranial magnetic double stimulation. Med. Sci. Sports Exerc. 2000, 32, 1942–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lazzaro, V.; Oliviero, A.; Tonali, P.A.; Mazzone, P.; Insola, A.; Pilato, F.; Saturno, E.; Dileone, M.; Rothwell, J.C. Direct demonstration of reduction of the output of the human motor cortex induced by a fatiguing muscle contraction. Exp. Brain Res. 2003, 149, 535–538. [Google Scholar] [CrossRef]
- Vucic, S.; Cheah, B.C.; Kiernan, M.C. Dissecting the mechanisms underlying short-interval intracortical inhibition using exercise. Cereb. Cortex 2011, 21, 1639–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattey, J.; Martin, P.G.; Kay, D.; Cannon, J.; Marino, F.E. Contralateral muscle fatigue in human quadriceps muscle: Evidence for a centrally mediated fatigue response and cross-over effect. Pflug. Arch. 2006, 452, 199–207. [Google Scholar] [CrossRef]
- Halperin, I.; Copithorne, D.; Behm, D.G. Unilateral isometric muscle fatigue decreases force production and activation of contralateral knee extensors but not elbow flexors. Appl. Physiol. Nutr. Metab. 2014, 39, 1338–1344. [Google Scholar] [CrossRef]
- Aboodarda, S.J.; Šambaher, N.; Behm, D.G. Unilateral elbow flexion fatigue modulates corticospinal responsiveness in non-fatigued contralateral biceps brachii: Contralateral corticospinal responsiveness. Scand. J. Med. Sci. Sports 2016, 26, 1301–1312. [Google Scholar] [CrossRef]
- Aboodarda, S.J.; Copithorne, D.B.; Power, K.E.; Drinkwater, E.; Behm, D.G. Elbow flexor fatigue modulates central excitability of the knee extensors. Appl. Physiol. Nutr. Metab. 2015, 40, 924–930. [Google Scholar] [CrossRef]
- Behm, D.G. Bilateral Knee Extensor Fatigue Modulates Force and Responsiveness of the Corticospinal Pathway in the Non-fatigued, Dominant Elbow Flexors. Front. Hum. Neurosci. 2016, 10, 11. [Google Scholar]
- Amann, M.; Venturelli, M.; Ives, S.J.; McDaniel, J.; Layec, G.; Rossman, M.J.; Richardson, R.S. Peripheral fatigue limits endurance exercise via a sensory feedback-mediated reduction in spinal motoneuronal output. J. Appl. Physiol. 2013, 115, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Vallbo, A.B. Afferent discharge from human muscle spindles in non-contracting muscles. Steady state impulse frequency as a function of joint angle. Acta Physiol. Scand. 1974, 90, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Macefield, G.; Hagbarth, K.E.; Gorman, R.; Gandevia, S.C.; Burke, D. Decline in spindle support to alpha-motoneurones during sustained voluntary contractions. J. Physiol. 1991, 440, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Hutton, R.S.; Eldred, E. Postcontraction changes in sensitivity of muscle afferents to static and dynamic stretch. Brain Res. 1974, 78, 193–202. [Google Scholar] [CrossRef]
- Hutton, R.S.; Nelson, D.L. Stretch sensitivity of Golgi tendon organs in fatigued gastrocnemius muscle. Med. Sci. Sports Exerc. 1986, 18, 69–74. [Google Scholar] [CrossRef]
- Kernell, D.; Monster, A.W. Time course and properties of late adaptation in spinal motoneurones of the cat. Exp. Brain Res. 1982, 46, 191–196. [Google Scholar] [CrossRef]
- Brownstone, R.M.; Jordan, L.M.; Kriellaars, D.J.; Noga, B.R.; Shefchyk, S.J. On the regulation of repetitive firing in lumbar motoneurones during fictive locomotion in the cat. Exp. Brain Res. 1992, 90, 441–455. [Google Scholar] [CrossRef]
- Sawczuk, A.; Powers, R.K.; Binder, M.D. Spike frequency adaptation studied in hypoglossal motoneurons of the rat. J. Neurophysiol. 1995, 73, 1799–1810. [Google Scholar] [CrossRef]
- Garland, S.J.; McComas, A.J. Reflex inhibition of human soleus muscle during fatigue. J. Physiol. 1990, 429, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Pierrot-Deseilligny, E. Propriospinal transmission of part of the corticospinal excitation in humans. Muscle Nerve 2002, 26, 155–172. [Google Scholar] [CrossRef]
- Burke, D.; Gracies, J.M.; Mazevet, D.; Meunier, S.; Pierrot-Deseilligny, E. Non-monosynaptic transmission of the cortical command for voluntary movement in man. J. Physiol. 1994, 480, 191–202. [Google Scholar] [CrossRef]
- Martin, P.G.; Gandevia, S.C.; Taylor, J.L. Muscle fatigue changes cutaneous suppression of propriospinal drive to human upper limb muscles: Propriospinal drive with fatigue. J. Physiol. 2007, 580, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.A.; Rothwell, J.C. Distinct Influence of Hand Posture on Cortical Activity during Human Grasping. J. Neurosci. 2015, 35, 4882–4889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuzzo, J.L.; Trajano, G.S.; Barry, B.K.; Gandevia, S.C.; Taylor, J.L. Arm posture-dependent changes in corticospinal excitability are largely spinal in origin. J. Neurophysiol. 2016, 115, 2076–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, A.J.; Alcock, L.; Lockyer, E.; Button, D.; Power, K. Phase- and Workload-Dependent Changes in Corticospinal Excitability to the Biceps and Triceps Brachii during Arm Cycling. Brain Sci. 2016, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Forman, D.A.; Abdel-Malek, D.; Bunce, C.M.F.; Holmes, M.W.R. Muscle length and joint angle influence spinal but not corticospinal excitability to the biceps brachii across forearm postures. J. Neurophysiol. 2019, 122, 413–423. [Google Scholar] [CrossRef]
- Smith, J.L.; Martin, P.G.; Gandevia, S.C.; Taylor, J.L. Sustained contraction at very low forces produces prominent supraspinal fatigue in human elbow flexor muscles. J. Appl. Physiol. 2007, 103, 560–568. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FCR | FDS | FCU | ECR | EDC | ECU | ||
|---|---|---|---|---|---|---|---|
| Flexion | p-value | 0.54 | 0.60 | 0.14 | 0.02 * | 0.01 * | 0.07 |
| F-Statistic | F(14,182) = 0.92 | F(14,182) = 0.86 | F(14,182) = 1.43 | F(14,182) = 2.03 | F(14,182) = 2.12 | F(14,182) = 1.69 | |
| Effect Size | 0.07 | 0.06 | 0.12 | 0.16 | 0.15 | 0.16 | |
| Extension | p-value | <0.001 * | 0.02 * | 0.32 | 0.29 | <0.001 * | 0.19 |
| F-Statistic | F(14,182) = 3.84 | F(14,182) = 1.98 | F(14,182) = 1.14 | F(14,182) = 1.19 | F(14,182) = 3.94 | F(14,182) = 1.35 | |
| Effect Size | 0.23 | 0.13 | 0.09 | 0.10 | 0.25 | 0.13 |
| FCR | FDS | FCU | ECR | EDC | ECU | ||
|---|---|---|---|---|---|---|---|
| Session | p-value | 0.11 | 0.97 | 0.24 | 0.004 * | 0.78 | 0.10 |
| F-Statistic | F(1,13) = 3.03 | F(1,13) = 0.002 | F(1,11) = 1.52 | F(1,11) = 13.0 | F(1,12) = 0.08 | F(1,9) = 3.34 | |
| Effect Size | 0.19 | <0.001 | 0.12 | 0.54 | 0.01 | 0.27 | |
| Time | p-value | <0.001 * | <0.001 * | <0.001 * | 0.75 | 0.001 * | 0.40 |
| F-Statistic | F(13,169) = 4.91 | F(13,169) = 4.04 | F(13,143) = 3.22 | F(13,143) = 0.71 | F(13,156) = 2.76 | F(13,117) = 1.06 | |
| Effect Size | 0.27 | 0.24 | 0.23 | 0.06 | 0.19 | 0.11 | |
| Interaction | p-value | 0.51 | 0.94 | 0.68 | 0.014 * | 0.003 * | 0.02 * |
| F-Statistic | F(13,169) = 0.95 | F(13,169) = 0.47 | F(13,143) = 0.78 | F(13,143) = 2.20 | F(13,156) = 2.55 | F(13,117) = 2.11 | |
| Effect Size | 0.07 | 0.04 | 0.07 | 0.16 | 0.18 | 0.19 |
| FCR | FDS | FCU | ECR | EDC | ECU | ||
|---|---|---|---|---|---|---|---|
| Flexion | p-value | 0.12 | 0.06 | <0.001 * | 0.20 | <0.001 * | 0.12 |
| F-Statistic | F(14,182) = 1.49 | F(14,182) = 1.68 | F(14,154) = 3.30 | F(14,154) = 1.33 | F(14,168) = 3.93 | F(14,140) = 1.49 | |
| Effect Size | 0.10 | 0.11 | 0.23 | 0.11 | 0.25 | 0.13 | |
| Extension | p-value | <0.001 * | <0.001 * | 0.01 * | 0.08 | 0.10 | 0.14 |
| F-Statistic | F(14,182) = 3.87 | F(14,182) = 3.45 | F(14,154) = 2.15 | F(14,154) = 1.61 | F(14,168) = 1.54 | F(14,140) = 1.44 | |
| Effect Size | 0.23 | 0.21 | 0.16 | 0.13 | 0.11 | 0.13 |
| FCR | FDS | FCU | ECR | EDC | ECU | ||
|---|---|---|---|---|---|---|---|
| Session | p-value | 0.25 | 0.21 | 0.12 | 0.006 * | 0.22 | 0.68 |
| F-Statistic | F(1,13) = 1.44 | F(1,13) = 1.73 | F(1,11) = 2.91 | F(1,11) = 11.46 | F(1,12) = 1.70 | F(1,10) = 0.19 | |
| Effect Size | 0.10 | 0.12 | 0.21 | 0.51 | 0.12 | 0.02 | |
| Time | p-value | <0.001 * | <0.001 * | <0.001 * | 0.001 * | <0.001 * | 0.002 * |
| F-Statistic | F(13,169) = 5.16 | F(13,169) = 5.38 | F(13,143) = 5.52 | F(13,143) = 2.95 | F(13,156) = 4.82 | F(13,130) = 2.67 | |
| Effect Size | 0.28 | 0.29 | 0.33 | 0.21 | 0.29 | 0.21 | |
| Interaction | p-value | 0.04 * | 0.41 | 0.19 | 0.43 | 0.58 | 0.33 |
| F-Statistic | F(13,169) = 1.84 | F(13,169) = 1.05 | F(13,143) = 1.35 | F(13,143) = 1.03 | F(13,156) = 0.88 | F(13,130) = 1.14 | |
| Effect Size | 0.12 | 0.07 | 0.11 | 0.09 | 0.07 | 0.10 |
| FCR | FDS | FCU | ECR | EDC | ECU | ||
|---|---|---|---|---|---|---|---|
| Session | p-value | 0.81 | 0.26 | 0.02 * | 0.90 | 0.75 | 0.16 |
| F-Statistic | F(1,13) = 0.06 | F(1,13) = 1.41 | F(1,11) = 6.85 | F(1,11) = 0.02 | F(1,12) = 0.11 | F(1,10) = 2.38 | |
| Effect Size | 0.01 | 0.10 | 0.38 | 0.002 | 0.01 | 0.21 | |
| Time | p-value | 0.18 | 0.08 | 0.18 | <0.001 * | <0.001 * | 0.002 * |
| F-Statistic | F(13,169) = 1.55 | F(13,169) = 1.95 | F(13,143) = 1.70 | F(13,143) = 4.12 | F(13,156) = 3.43 | F(13,130) = 2.71 | |
| Effect Size | 0.11 | 0.13 | 0.13 | 0.27 | 0.22 | 0.23 | |
| Interaction | p-value | 0.21 | 0.94 | 0.80 | <0.001 * | 0.012 * | 0.004 * |
| F-Statistic | F(13,169) = 1.47 | F(13,169) = 0.46 | F(13,143) = 0.66 | F(13,143) = 3.95 | F(13,156) = 2.20 | F(13,130) = 2.56 | |
| Effect Size | 0.10 | 0.03 | 0.06 | 0.26 | 0.16 | 0.22 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forman, D.A.; Forman, G.N.; Murphy, B.A.; Holmes, M.W.R. Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions on Corticospinal Excitability to Forearm Muscles during Low-Intensity Hand-Gripping. Brain Sci. 2020, 10, 445. https://doi.org/10.3390/brainsci10070445
Forman DA, Forman GN, Murphy BA, Holmes MWR. Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions on Corticospinal Excitability to Forearm Muscles during Low-Intensity Hand-Gripping. Brain Sciences. 2020; 10(7):445. https://doi.org/10.3390/brainsci10070445
Chicago/Turabian StyleForman, Davis A., Garrick N. Forman, Bernadette A. Murphy, and Michael W. R. Holmes. 2020. "Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions on Corticospinal Excitability to Forearm Muscles during Low-Intensity Hand-Gripping" Brain Sciences 10, no. 7: 445. https://doi.org/10.3390/brainsci10070445
APA StyleForman, D. A., Forman, G. N., Murphy, B. A., & Holmes, M. W. R. (2020). Sustained Isometric Wrist Flexion and Extension Maximal Voluntary Contractions on Corticospinal Excitability to Forearm Muscles during Low-Intensity Hand-Gripping. Brain Sciences, 10(7), 445. https://doi.org/10.3390/brainsci10070445