Consecutive Treatment with Brain-Derived Neurotrophic Factor and Electrical Stimulation Has a Protective Effect on Primary Auditory Neurons

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Subjects
2.2. Electrode Drug Delivery Device
2.3. Acoustically Evoked Auditory Brainstem Response (AABR)
2.4. Deafening Procedure
2.5. Surgical Implantation Procedure of the Electrode and the Pump
2.6. Electrically Evoked Auditory Brainstem Response (EABR)
2.7. Impedance Measurement
2.8. Chronic Electrical Stimulation
2.9. Histological Procedures
2.10. Data Analysis
3. Results
3.1. AABR and Deafening
3.2. Functional Results Based on EABR Measurements
3.3. Impedance Measurements
3.4. Histological Results
3.4.1. Spiral Ganglion Cell Survival within Each Group
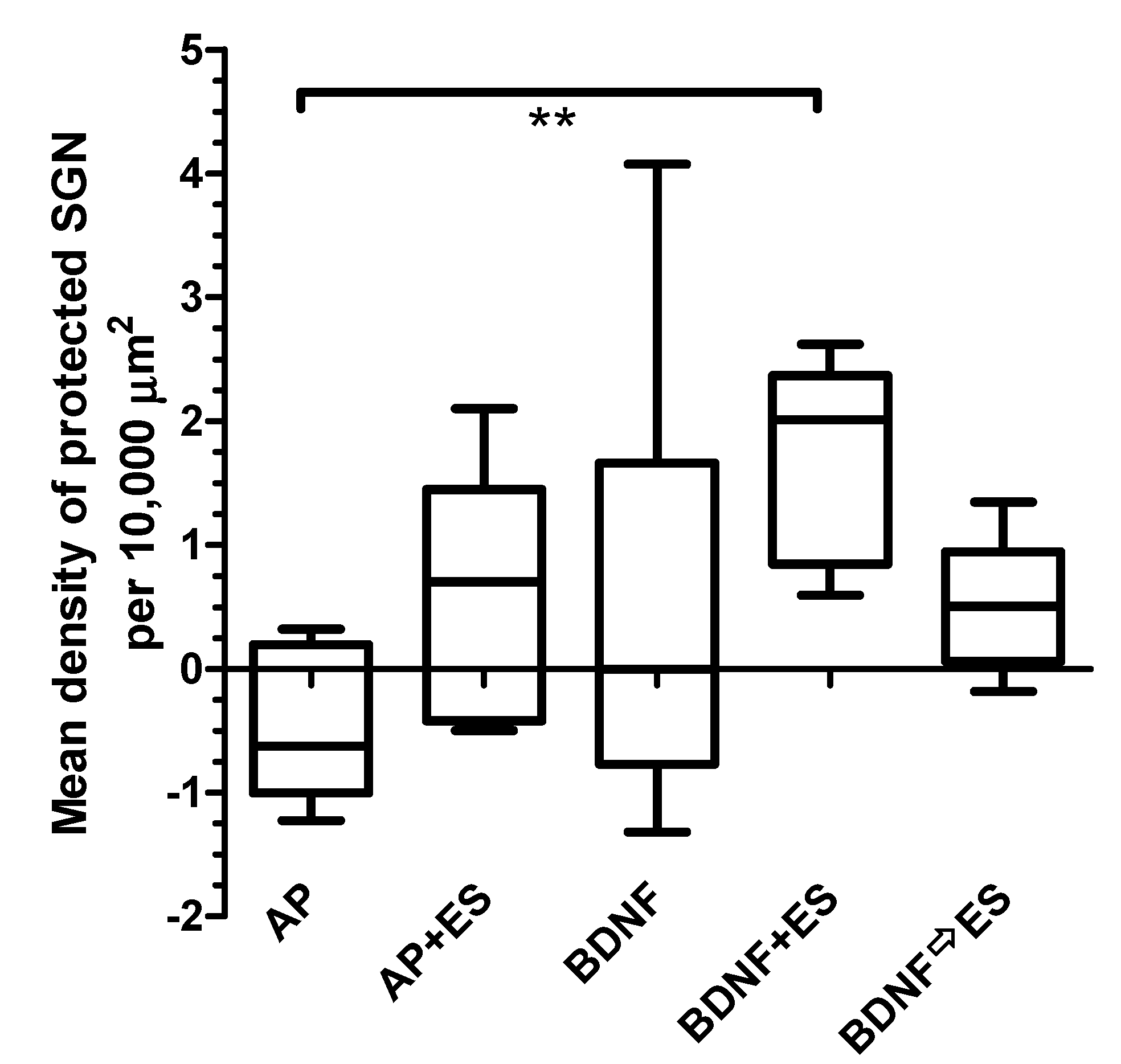
3.4.2. Protected Spiral Ganglion Cells
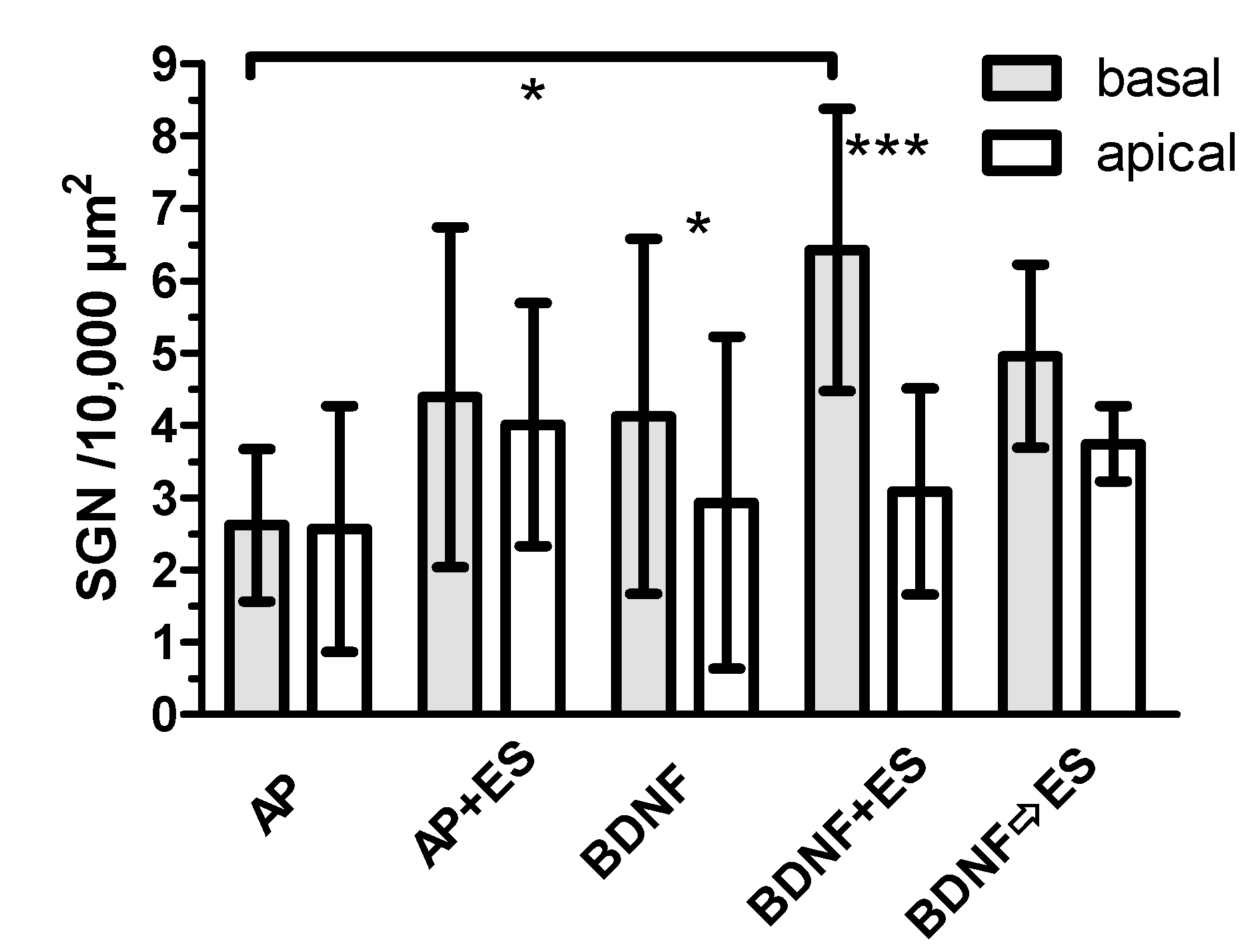
3.4.3. Comparison between Basal and Apical Turns
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Webster, M.; Webster, D.B. Spiral ganglion neuron loss following organ of Corti loss: A quantitative study. Brain Res. 1981, 212, 17–30. [Google Scholar] [CrossRef]
- Spoendlin, H. Degeneration behaviour of the cochlear nerve. Arch. Klin. Exp. Ohren Nasen Kehlkopfheilkd 1971, 200, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Ylikoski, J. Correlative studies on the cochlear pathology and hearing loss in guinea-pigs after intoxication with ototoxic antibiotics. Acta Otolaryngol. Suppl. 1974, 326, 1–62. [Google Scholar] [PubMed]
- Dodson, H.C.; Mohuiddin, A. Response of spiral ganglion neurones to cochlear hair cell destruction in the guinea pig. J. Neurocytol. 2000, 29, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Miller, J.M.; Finger, P.A.; Heller, J.W.; Raphael, Y.; Altschuler, R.A. Effects of chronic high-rate electrical stimulation on the cochlea and eighth nerve in the deafened guinea pig. Hear. Res. 1997, 105, 30–43. [Google Scholar] [CrossRef]
- Miller, J.M.; Altschuler, R.A. Effectiveness of different electrical stimulation conditions in preservation of spiral ganglion cells following deafness. Ann. Otol. Rhinol. Laryngol. Suppl. 1995, 166, 57–60. [Google Scholar]
- Hartshorn, D.O.; Miller, J.M.; Altschuler, R.A. Protective effect of electrical stimulation in the deafened guinea pig cochlea. Otolaryngol. Head Neck Surg. 1991, 104, 311–319. [Google Scholar] [CrossRef]
- Li, L.; Parkins, C.W.; Webster, D.B. Does electrical stimulation of deaf cochleae prevent spiral ganglion degeneration? Hear. Res. 1999, 133, 27–39. [Google Scholar] [CrossRef]
- Agterberg, M.J.; Versnel, H.; de Groot, J.C.; van den Broek, M.; Klis, S.F. Chronic electrical stimulation does not prevent spiral ganglion cell degeneration in deafened guinea pigs. Hear. Res. 2010, 269, 169–179. [Google Scholar] [CrossRef]
- Chen, I.; Limb, C.J.; Ryugo, D.K. The effect of cochlear-implant-mediated electrical stimulation on spiral ganglion cells in congenitally deaf white cats. J. Assoc. Res. Otolaryngol. 2010, 11, 587–603. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, L.N.; Shepherd, R.K. Clinical application of neurotrophic factors: The potential for primary auditory neuron protection. Eur. J. Neurosci. 2005, 22, 2123–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwieger, J.; Hamm, A.; Gepp, M.M.; Schulz, A.; Hoffmann, A.; Lenarz, T.; Scheper, V. Alginate-encapsulated brain-derived neurotrophic factor–overexpressing mesenchymal stem cells are a promising drug delivery system for protection of auditory neurons. J. Tissue Eng. 2020, 11, 2041731420911313. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, A.; Scheper, V.; Buhr, I.; Wenzel, G.I.; Wissel, K.; Paasche, G.; Berkingali, N.; Jorgensen, J.R.; Lenarz, T.; Stover, T. Artemin improves survival of spiral ganglion neurons in vivo and in vitro. Neuroreport 2010, 21, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Chikar, J.A.; Colesa, D.J.; Swiderski, D.L.; Di Polo, A.; Raphael, Y.; Pfingst, B.E. Over-expression of BDNF by adenovirus with concurrent electrical stimulation improves cochlear implant thresholds and survival of auditory neurons. Hear. Res. 2008, 245, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, R.K.; Coco, A.; Epp, S.B.; Crook, J.M. Chronic depolarization enhances the trophic effects of brain-derived neurotrophic factor in rescuing auditory neurons following a sensorineural hearing loss. J. Comp. Neurol. 2005, 486, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, S.; Stover, T.; Kawamoto, K.; Prieskorn, D.M.; Altschuler, R.A.; Miller, J.M.; Raphael, Y. Glial cell line-derived neurotrophic factor and chronic electrical stimulation prevent VIII cranial nerve degeneration following denervation. J. Comp. Neurol. 2002, 454, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Scheper, V.; Paasche, G.; Miller, J.M.; Warnecke, A.; Berkingali, N.; Lenarz, T.; Stover, T. Effects of delayed treatment with combined GDNF and continuous electrical stimulation on spiral ganglion cell survival in deafened guinea pigs. J. Neurosci. Res. 2009, 87, 1389–1399. [Google Scholar] [CrossRef] [Green Version]
- Jyung, R.W.; Miller, J.M.; Cannon, S.C. Evaluation of eighth nerve integrity by the electrically evoked middle latency response. Otolaryngol. Head Neck Surg. 1989, 101, 670–682. [Google Scholar] [CrossRef]
- Miller, J.M.; Le Prell, C.G.; Prieskorn, D.M.; Wys, N.L.; Altschuler, R.A. Delayed neurotrophin treatment following deafness rescues spiral ganglion cells from death and promotes regrowth of auditory nerve peripheral processes: Effects of brain-derived neurotrophic factor and fibroblast growth factor. J. Neurosci. Res. 2007, 85, 1959–1969. [Google Scholar] [CrossRef] [Green Version]
- Song, B.N.; Li, Y.X.; Han, D.M. Delayed electrical stimulation and BDNF application following induced deafness in rats. Acta Otolaryngol. 2008, 129, 1–13. [Google Scholar] [CrossRef]
- Shepherd, R.K.; Coco, A.; Epp, S.B. Neurotrophins and electrical stimulation for protection and repair of spiral ganglion neurons following sensorineural hearing loss. Hear. Res. 2008, 242, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Leake, P.A.; Stakhovskaya, O.; Hetherington, A.; Rebscher, S.J.; Bonham, B. Effects of brain-derived neurotrophic factor (BDNF) and electrical stimulation on survival and function of cochlear spiral ganglion neurons in deafened, developing cats. J. Assoc. Res. Otolaryngol. 2013, 14, 187–211. [Google Scholar] [CrossRef] [Green Version]
- West, B.A.; Brummett, R.E.; Himes, D.L. Interaction of kanamycin and ethacrynic acid. Severe cochlear damage in guinea pigs. Arch. Otolaryngol. 1973, 98, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Meyer, H.; Stover, T.; Fouchet, F.; Bastiat, G.; Saulnier, P.; Baumer, W.; Lenarz, T.; Scheper, V. Lipidic nanocapsule drug delivery: Neuronal protection for cochlear implant optimization. Int. J. Nanomed. 2012, 7, 2449–2464. [Google Scholar]
- Staecker, H.; Kopke, R.; Malgrange, B.; Lefebvre, P.; Van de Water, T.R. NT-3 and/or BDNF therapy prevents loss of auditory neurons following loss of hair cells. Neuroreport 1996, 7, 889–894. [Google Scholar] [CrossRef]
- Gillespie, L.N.; Clark, G.M.; Bartlett, P.F.; Marzella, P.L. BDNF-induced survival of auditory neurons in vivo: Cessation of treatment leads to accelerated loss of survival effects. J. Neurosci. Res. 2003, 71, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Nakaizumi, T.; Kawamoto, K.; Minoda, R.; Raphael, Y. Adenovirus-mediated expression of brain-derived neurotrophic factor protects spiral ganglion neurons from ototoxic damage. Audiol. Neurootol. 2004, 9, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, T.; Bredberg, G.; Ulfendahl, M.; Pyykko, I.; Olivius, N.P.; Kaksonen, R.; Lindstrom, B.; Altschuler, R.; Miller, J.M. Neurotrophic factor intervention restores auditory function in deafened animals. Proc. Natl. Acad. Sci. USA 2002, 99, 1657–1660. [Google Scholar] [CrossRef] [Green Version]
- Wefstaedt, P.; Scheper, V.; Lenarz, T.; Stover, T. Brain-derived neurotrophic factor/glial cell line-derived neurotrophic factor survival effects on auditory neurons are not limited by dexamethasone. Neuroreport 2005, 16, 2011–2014. [Google Scholar] [CrossRef]
- Miller, J.M.; Chi, D.H.; O’Keeffe, L.J.; Kruszka, P.; Raphael, Y.; Altschuler, R.A. Neurotrophins can enhance spiral ganglion cell survival after inner hair cell loss. Int. J. Dev. Neurosci. 1997, 15, 631–643. [Google Scholar] [CrossRef]
- Leake, P.A.; Hradek, G.T.; Snyder, R.L. Chronic electrical stimulation by a cochlear implant promotes survival of spiral ganglion neurons after neonatal deafness. J. Comp. Neurol. 1999, 412, 543–562. [Google Scholar] [CrossRef]
- Miller, A.L.; Prieskorn, D.M.; Altschuler, R.A.; Miller, J.M. Mechanism of electrical stimulation-induced neuroprotection: Effects of verapamil on protection of primary auditory afferents. Brain Res. 2003, 966, 218–230. [Google Scholar] [CrossRef]
- Araki, S.; Kawano, A.; Seldon, L.; Shepherd, R.K.; Funasaka, S.; Clark, G.M. Effects of chronic electrical stimulation on spiral ganglion neuron survival and size in deafened kittens. Laryngoscope 1998, 108, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.K.; Matsushima, J.; Martin, R.L.; Clark, G.M. Cochlear pathology following chronic electrical stimulation of the auditory nerve: II. Deafened kittens. Hear. Res. 1994, 81, 150–166. [Google Scholar] [CrossRef]
- Gallo, V.; Kingsbury, A.; Balazs, R.; Jorgensen, O.S. The role of depolarization in the survival and differentiation of cerebellar granule cells in culture. J. Neurosci. 1987, 7, 2203–2213. [Google Scholar] [CrossRef]
- Koike, T.; Martin, D.P.; Johnson, E.M., Jr. Role of Ca2+ channels in the ability of membrane depolarization to prevent neuronal death induced by trophic-factor deprivation: Evidence that levels of internal Ca2+ determine nerve growth factor dependence of sympathetic ganglion cells. Proc. Natl. Acad. Sci. USA 1989, 86, 6421–6425. [Google Scholar] [CrossRef] [Green Version]
- Collins, F.; Schmidt, M.F.; Guthrie, P.B.; Kater, S.B. Sustained increase in intracellular calcium promotes neuronal survival. J. Neurosci. 1991, 11, 2582–2587. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.W. Calcium-mediated neurotoxicity: Relationship to specific channel types and role in ischemic damage. Trends Neurosci. 1988, 11, 465–469. [Google Scholar] [CrossRef]
- Kristian, T.; Ouyang, Y.; Siesjo, B.K. Calcium-induced neuronal cell death in vivo and in vitro: Are the pathophysiologic mechanisms different? Adv. Neurol. 1996, 71, 107–113, discussion 113–108. [Google Scholar]
- Koike, T.; Tanaka, S. Evidence that nerve growth factor dependence of sympathetic neurons for survival in vitro may be determined by levels of cytoplasmic free Ca2+. Proc. Natl. Acad. Sci. USA 1991, 88, 3892–3896. [Google Scholar] [CrossRef] [Green Version]
- Hegarty, J.L.; Kay, A.R.; Green, S.H. Trophic support of cultured spiral ganglion neurons by depolarization exceeds and is additive with that by neurotrophins or cAMP and requires elevation of [Ca2+] i within a set range. J. Neurosci. 1997, 17, 1959–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwolan, T.A.; Collins, L.M.; Wakefield, G.H. Electrode discrimination and speech recognition in postlingually deafened adult cochlear implant subjects. J. Acoust. Soc. Am. 1997, 102, 3673–3685. [Google Scholar] [CrossRef] [PubMed]
- Rocamora, N.; Welker, E.; Pascual, M.; Soriano, E. Upregulation of BDNF mRNA expression in the barrel cortex of adult mice after sensory stimulation. J. Neurosci. 1996, 16, 4411–4419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, S.A.; Mack, K.J. Seizures and sensory stimulation result in different patterns of brain derived neurotrophic factor protein expression in the barrel cortex and hippocampus. Brain Res. Mol. Brain Res. 2000, 78, 1–14. [Google Scholar] [CrossRef]
- Balkowiec, A.; Katz, D.M. Activity-dependent release of endogenous Brain-Derived Neurotrophic Factor from primary sensory neurons detected by ELISA in situ. J. Neurosci. 2000, 20, 7417–7423. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Feng, L.; Yang, F.; Lu, B. Activity- and Ca(2+)-dependent modulation of surface expression of brain-derived neurotrophic factor receptors in hippocampal neurons. J. Cell Biol. 2000, 150, 1423–1434. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheper, V.; Seidel-Effenberg, I.; Lenarz, T.; Stöver, T.; Paasche, G. Consecutive Treatment with Brain-Derived Neurotrophic Factor and Electrical Stimulation Has a Protective Effect on Primary Auditory Neurons. Brain Sci. 2020, 10, 559. https://doi.org/10.3390/brainsci10080559
Scheper V, Seidel-Effenberg I, Lenarz T, Stöver T, Paasche G. Consecutive Treatment with Brain-Derived Neurotrophic Factor and Electrical Stimulation Has a Protective Effect on Primary Auditory Neurons. Brain Sciences. 2020; 10(8):559. https://doi.org/10.3390/brainsci10080559
Chicago/Turabian StyleScheper, Verena, Ira Seidel-Effenberg, Thomas Lenarz, Timo Stöver, and Gerrit Paasche. 2020. "Consecutive Treatment with Brain-Derived Neurotrophic Factor and Electrical Stimulation Has a Protective Effect on Primary Auditory Neurons" Brain Sciences 10, no. 8: 559. https://doi.org/10.3390/brainsci10080559
APA StyleScheper, V., Seidel-Effenberg, I., Lenarz, T., Stöver, T., & Paasche, G. (2020). Consecutive Treatment with Brain-Derived Neurotrophic Factor and Electrical Stimulation Has a Protective Effect on Primary Auditory Neurons. Brain Sciences, 10(8), 559. https://doi.org/10.3390/brainsci10080559