Expression of Dopamine-Related Genes in Four Human Brain Regions



Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Single Tissue Genotype x Expression Analyses
3.2. Single Tissue Expression Analyses by Demographic and Phenotypic Characteristics
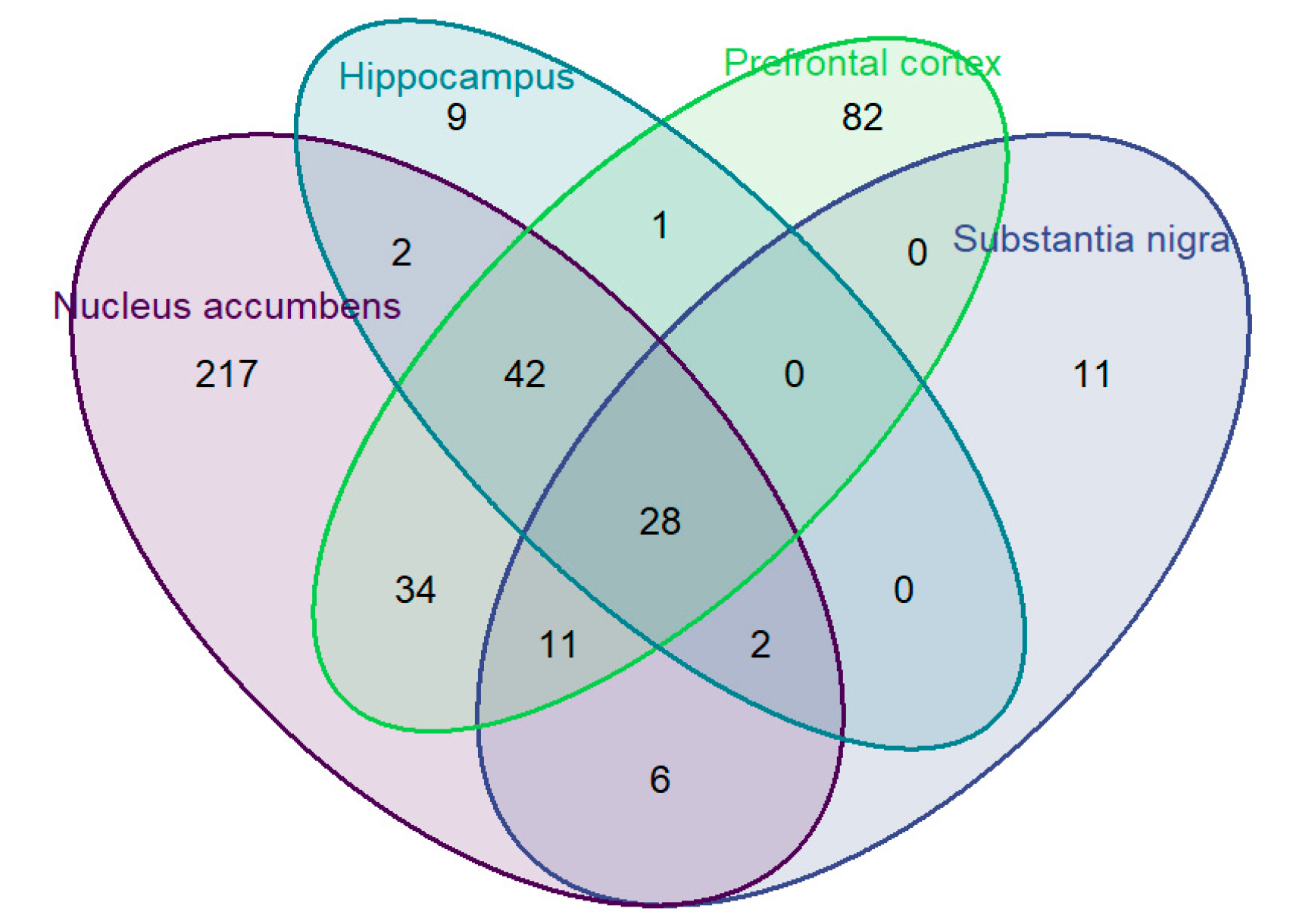
3.3. Four Tissue Genotype x Expression Analyses
4. Discussion
4.1. Age
4.2. Sex
4.3. BMI
4.4. Genotype x Expression Associations
4.5. Haplotype Block 2: rs10891549 and rs6277
4.6. Haplotype Block 3: 25 Variants in DRD2
4.7. Limitations
4.8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bühler, K.-M.; Giné, E.; Alzate, V.E.; Calleja-Conde, J.; De Fonseca, F.R.; López-Moreno, J. Common single nucleotide variants underlying drug addiction: More than a decade of research. Addict. Biol. 2015, 20, 845–871. [Google Scholar] [CrossRef]
- Clarke, T.-K.; Adams, M.J.; Davies, G.; Howard, D.M.; Hall, L.S.; Padmanabhan, S.; Murray, A.D.; Smith, B.H.; Campbell, A.; Hayward, C.; et al. Genome-wide association study of alcohol consumption and genetic overlap with other health-related traits in UK Biobank (N=112 117). Mol. Psychiatry 2017, 22, 1376–1384. [Google Scholar] [CrossRef] [Green Version]
- Howes, O.D.; McCutcheon, R.A.; Owen, M.J.; Murray, R.M. The Role of Genes, Stress, and Dopamine in the Development of Schizophrenia. Biol. Psychiatry 2017, 81, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Nakata, Y.; Kanahara, N.; Iyo, M. Dopamine supersensitivity psychosis in schizophrenia: Concepts and implications in clinical practice. J. Psychopharmacol. 2017, 31, 1511–1518. [Google Scholar] [CrossRef]
- Tang, S.; Yao, B.; Li, N.; Lin, S.; Huang, Z. Association of Dopamine Beta-Hydroxylase Polymorphisms with Alzheimer’s Disease, Parkinson’s Disease and Schizophrenia: Evidence Based on Currently Available Loci. Cell. Physiol. Biochem. 2018, 51, 411–428. [Google Scholar] [CrossRef]
- Kang, S.; Bi, M.; Du, X.; Jiao, Q.; Jiang, H. Association of the rs1611115 polymorphism in DBH gene with Parkinson’s disease: A meta-analysis. Neurol. Sci. 2018, 39, 2085–2089. [Google Scholar] [CrossRef]
- Correa, D.D.; Satagopan, J.; Martin, A.; Braun, E.; Kryza-Lacombe, M.; Cheung, K.; Sharma, A.; Dimitriadoy, S.; O’Connell, K.; Leong, S.; et al. Genetic variants and cognitive functions in patients with brain tumors. Neuro-Oncology 2019, 21, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Al-Hawi, Z.; Cummins, T.D.R.; Tong, J.; Johnson, B.P.; Lau, R.; Samarrai, W.; Bellgrove, M.A. The molecular genetic architecture of attention deficit hyperactivity disorder. Mol. Psychiatry 2015, 20, 289–297. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Azadmarzabadi, E.; Haghighatfard, A.; Mohammadi, A. Low resilience to stress is associated with candidate gene expression alterations in the dopaminergic signalling pathway. Psychogeriatrics 2018, 18, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.L.; A Kassir, S.; Underwood, M.D.; Bakalian, M.J.; Mann, J.J.; Arango, V. Dysregulation of Striatal Dopamine Receptor Binding in Suicide. Neuropsychopharmacology 2016, 42, 974–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitra, S.; Sarkar, K.; Sinha, S.; Mukhopadhyay, K. The Dopamine Receptor D5 May Influence Age of Onset. J. Child Neurol. 2016, 31, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Rothmond, D.A.; Weickert, C.S.; Webster, M.J. Developmental changes in human dopamine neurotransmission: Cortical receptors and terminators. BMC Neurosci. 2012, 13, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkinson, G.M.; Dayas, C.V.; Smith, D.W. Age-related gene expression changes in substantia nigra dopamine neurons of the rat. Mech. Ageing Dev. 2015, 149, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.; Kanungo, M. Expression of D2 dopamine receptor in the mouse brain. Biochem. Biophys. Res. Commun. 2006, 344, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Green, A.L.; Eid, A.; Zhan, L.; Zarbl, H.; Guo, G.L.; Richardson, J.R. Epigenetic Regulation of the Ontogenic Expression of the Dopamine Transporter. Front. Genet. 2019, 10, 1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinares-Garcia, P.; Stratikopoulos, M.; Zagato, A.; Loke, H.; Lee, J. Sex: A Significant Risk Factor for Neurodevelopmental and Neurodegenerative Disorders. Brain Sci. 2018, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendrek, A.; Mancini-Marïe, A. Sex/gender differences in the brain and cognition in schizophrenia. Neurosci. Biobehav. Rev. 2016, 67, 57–78. [Google Scholar] [CrossRef]
- Loke, H.; Harley, V.; Lee, J. Biological factors underlying sex differences in neurological disorders. Int. J. Biochem. Cell Biol. 2015, 65, 139–150. [Google Scholar] [CrossRef]
- Sun, X.; Luquet, S.H.; Small, D.M. DRD2: Bridging the Genome and Ingestive Behavior. Trends Cogn. Sci. 2017, 21, 372–384. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Liu, Y.; Shriner, R.; Gold, M.S. Reward circuitry dopaminergic activation regulates food and drug craving behavior. Curr. Pharm. Des. 2011, 17, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Stanfill, A.; Hathaway, D.; Cashion, A.; Homayouni, R.; Cowan, P.; Thompson, C.; Madahian, B.; Conley, Y. A Pilot Study of Demographic and Dopaminergic Genetic Contributions to Weight Change in Kidney Transplant Recipients. PLoS ONE 2015, 10, e0138885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaiser, E.C.; Gallezot, J.-D.; Worhunsky, P.D.; Jastreboff, A.M.; Pittman, B.; Kantrovitz, L.; Angarita, G.A.; Cosgrove, K.P.; Potenza, M.N.; Malison, R.T.; et al. Elevated Dopamine D2/3 Receptor Availability in Obese Individuals: A PET Imaging Study with [11C](+)PHNO. Neuropsychopharmacology 2016, 41, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Vaanholt, L.M.; Mitchell, S.E.; Sinclair, R.E.; Speakman, J.R. Mice that are resistant to diet-induced weight loss have greater food anticipatory activity and altered melanocortin-3 receptor (MC3R) and dopamine receptor 2 (D2) gene expression. Horm. Behav. 2015, 73, 83–93. [Google Scholar] [CrossRef]
- Gao, X.; Wang, Y.; Lang, M.; Yuan, L.; Reece, A.S.; Wang, W. Contribution of Genetic Polymorphisms and Haplotypes in DRD2, BDNF, and Opioid Receptors to Heroin Dependence and Endophenotypes Among the Han Chinese. OMICS: A J. Integr. Biol. 2017, 21, 404–412. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, P.; Li, Y.; Cai, X.; Yang, Z.; Miao, X.; Chen, B.; Li, S.-B.; Dang, W.; Jia, W.; et al. A 35.8 kilobases haplotype spanning ANKK1 and DRD2 is associated with heroin dependence in Han Chinese males. Brain Res. 2018, 1688, 54–64. [Google Scholar] [CrossRef]
- Meyers, J.L.; Nyman, E.; Loukola, A.; Rose, R.J.; Kaprio, J.; Dick, D.M. The association between DRD2/ANKK1 and genetically informed measures of alcohol use and problems. Addict Biol. 2013, 18, 523–536. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Wainwright, M.S.; Comeron, J.M.; Saitou, N.; Sanders, A.R.; Gelernter, J.; Gejman, P.V. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA stability and synthesis of the receptor. Hum. Mol. Genet. 2003, 12, 205–216. [Google Scholar] [CrossRef]
- Markett, S.; De Reus, M.A.; Reuter, M.; Montag, C.; Weber, B.; Schoene-Bake, J.-C.; Heuvel, M.P.V.D. Variation on the dopamine D2 receptor gene (DRD2) is associated with basal ganglia-to-frontal structural connectivity. NeuroImage 2017, 155, 473–479. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, X.; Xiang, Q.; Zhou, S.; Zhao, N.; Xie, Q.; Zhao, X.; Zhou, Y.; Cui, Y. Association between dopamine receptor gene polymorphisms and effects of risperidone treatment: A systematic review and meta-analysis. Basic Clin. Pharmacol. Toxicol. 2018, 124, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Taurisano, P.; Romano, R.; Mancini, M.; Di Giorgio, A.; Antonucci, L.A.; Fazio, L.; Rampino, A.; Quarto, T.; Gelao, B.; Porcelli, A.; et al. Prefronto-striatal physiology is associated with schizotypy and is modulated by a functional variant of DRD2. Front. Behav. Neurosci. 2014, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Luykx, J.J.; Broersen, J.L.; De Leeuw, M. The DRD2 rs1076560 polymorphism and schizophrenia-related intermediate phenotypes: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2017, 74, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Stolf, A.R.; Cupertino, R.B.; Müller, D.; Sanvicente-Vieira, B.; Roman, T.; Vitola, E.S.; Grevet, E.H.; Von Diemen, L.; Kessler, F.; Grassi-Oliveira, R.; et al. Effects of DRD2 splicing-regulatory polymorphism and DRD4 48 bp VNTR on crack cocaine addiction. J. Neural Transm. 2018, 126, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Nyman, E.S.; Loukola, A.; Varilo, T.; Taanila, A.; Hurtig, T.; Moilanen, I.; Loo, S.; McGough, J.J.; Järvelin, M.-R.; Smalley, S.L.; et al. Sex-specific influence of DRD2 on ADHD-type temperament in a large population-based birth cohort. Psychiatr. Genet. 2012, 22, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Blasi, G.; Selvaggi, P.; Fazio, L.; Antonucci, L.A.; Taurisano, P.; Masellis, R.; Romano, R.; Mancini, M.; Zhang, F.; Caforio, G.; et al. Variation in Dopamine D2 and Serotonin 5-HT2A Receptor Genes is Associated with Working Memory Processing and Response to Treatment with Antipsychotics. Neuropsychopharmacology 2015, 40, 1600–1608. [Google Scholar] [CrossRef]
- Miller, N.S.; Chou, K.L.; I Bohnen, N.; Müller, M.L.T.M.; Seidler, R.D. Dopaminergic polymorphisms associated with medication responsiveness of gait in Parkinson’s disease. Park. Relat. Disord. 2018, 48, 54–60. [Google Scholar] [CrossRef]
- Suchanecka, A.; Chmielowiec, J.; Chmielowiec, K.; Masiak, J.; Sipak-Szmigiel, O.; Sznabowicz, M.; Czarny, W.; Michałowska-Sawczyn, M.; Trybek, G.; Grzywacz, A. Dopamine Receptor DRD2 Gene rs1076560, Personality Traits and Anxiety in the Polysubstance Use Disorder. Brain Sci. 2020, 10, 262. [Google Scholar] [CrossRef]
- Alblooshi, H.; Hulse, G.K.; Osman, W.; Elkashef, A.M.; Shawky, M.; Al Ghaferi, H.; Alsafar, H.; Tay, G.K. The frequency of DRD2 rs1076560 and OPRM1 rs1799971 in substance use disorder patients from the United Arab Emirates. Ann. Gen. Psychiatry 2018, 17, 22. [Google Scholar] [CrossRef] [Green Version]
- Sznabowicz, M.; Jasiewicz, A.; Iskra-Trifunović, J.; Małecka, I.; Karakiewicz, B.; Kotwas, A.; Samochowiec, J.; Grzywacz, A. Case-control study analysis of DRD2 gene polymorphisms in drug addicted patients. Psychiatr. Polska 2018, 52, 1013–1022. [Google Scholar] [CrossRef]
- Valli, M.; Cho, S.S.; Masellis, M.; Chen, R.; Rusjan, P.; Kim, J.; Koshimori, Y.; Mihaescu, A.; Strafella, A.P. DRD2 Genotype-Based Variants Modulates D2 Receptor Distribution in Ventral Striatum. Mol. Neurobiol. 2019, 56, 6512–6520. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Total Number of Genotype Variants | Number of Variants in Genotype x Expression Analysis | Number of Significant Genotype x Expression Variants in Nucleus Accumbens | Number of Significant Genotype x Expression Variants in Substantia Nigra | Number of Significant Genotype x Expression Variants in Hippocampus | Number of Significant Genotype x Expression Variants in Prefrontal Cortex |
|---|---|---|---|---|---|---|
| ANKK1 | 512 | 119 | 37 | 1 | 1 | 27 |
| DBH | 1011 | 232 | 25 | 14 | 65 | |
| DRD1 | 404 | 99 | 14 | 11 | 16 | |
| DRD2 | 1368 | 270 | 113 | 32 | 65 | 69 |
| DRD3 | 1335 | 299 | 163 | 2 | 2 | 7 |
| DRD5 | 478 | 68 | 2 | 1 | ||
| SLC6A3 | 1494 | 280 | 7 | 9 | 5 | 13 |
| Total | 6602 | 1367 | 361 1 | 58 | 84 | 198 |
| Nucleus Accumbens | Substantia Nigra | Hippocampus | Prefrontal Cortex | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rs ID | Ref. Allele | Alt. Allele | Alt. Allele Frequency | r | p | r | p | r | p | r | p |
| rs7103679 | T | C | 0.849 | −0.293 | 7.42 × 10−5 | −0.295 | 0.0056 | −0.339 | 0.0002 | −0.252 | 0.0048 |
| rs10891549 | T | C | 0.493 | −0.321 | 1.29 × 10−5 | −0.334 | 0.0016 | −0.305 | 0.0007 | −0.186 | 0.0390 |
| rs10891549 * | T | C | 0.493 | −0.182 | 2.47 × 10−2 | −0.334 | 0.0016 | −0.305 | 0.0007 | −0.186 | 0.0390 |
| rs6277 | G | A | 0.490 | −0.316 | 1.81 × 10−5 | −0.334 | 0.0016 | −0.293 | 0.0012 | −0.190 | 0.0349 |
| rs1079596 | C | T | 0.158 | 0.300 | 5.02 × 10−5 | 0.306 | 0.0039 | 0.333 | 0.0002 | 0.286 | 0.0013 |
| rs1125394 | T | C | 0.157 | 0.296 | 6.51 × 10−5 | 0.325 | 0.0023 | 0.333 | 0.0002 | 0.286 | 0.0013 |
| rs2471857 | C | T | 0.153 | 0.295 | 6.94 × 10−5 | 0.317 | 0.0028 | 0.322 | 0.0004 | 0.301 | 0.0007 |
| rs2471854 | G | C | 0.152 | 0.287 | 1.14 × 10−4 | 0.317 | 0.0028 | 0.309 | 0.0007 | 0.288 | 0.0012 |
| rs11214599 | C | T | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs2511520 | C | T | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs2242591 | C | T | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs6278 | C | A | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs1124491 | G | A | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs1079595 | A | C | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs1079594 | A | C | 0.151 | 0.285 | 1.22 × 10−4 | 0.290 | 0.0064 | 0.319 | 0.0004 | 0.261 | 0.0035 |
| rs1076560 | C | A | 0.149 | 0.288 | 1.02 × 10−4 | 0.290 | 0.0064 | 0.325 | 0.0003 | 0.263 | 0.0032 |
| rs2471855 | C | G | 0.149 | 0.291 | 8.48 × 10−5 | 0.278 | 0.0093 | 0.316 | 0.0005 | 0.305 | 0.0006 |
| rs2471851 | A | C | 0.149 | 0.271 | 2.68 × 10−4 | 0.276 | 0.0097 | 0.316 | 0.0005 | 0.279 | 0.0017 |
| rs55900980 | TG | T | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs2283265 | C | A | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs2075654 | C | T | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs1079727 | T | C | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs2734836 | C | T | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs1962262 | G | A | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs1079598 | A | G | 0.146 | 0.296 | 6.18 × 10−5 | 0.278 | 0.0093 | 0.328 | 0.0003 | 0.298 | 0.0008 |
| rs11214601 | C | T | 0.153 | 0.304 | 3.98 × 10−5 | 0.290 | 0.0064 | 0.337 | 0.0002 | 0.281 | 0.0016 |
| rs2242593 | T | C | 0.153 | 0.304 | 3.98 × 10−5 | 0.290 | 0.0064 | 0.337 | 0.0002 | 0.281 | 0.0016 |
| rs1125393 | C | T | 0.147 | 0.292 | 8.69 × 10−5 | 0.278 | 0.0093 | 0.303 | 0.0009 | 0.292 | 0.0010 |
| rs7350522 | G | T | 0.146 | 0.271 | 2.66 × 10−4 | 0.276 | 0.0097 | 0.328 | 0.0003 | 0.243 | 0.0065 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanfill, A.G.; Cao, X. Expression of Dopamine-Related Genes in Four Human Brain Regions. Brain Sci. 2020, 10, 567. https://doi.org/10.3390/brainsci10080567
Stanfill AG, Cao X. Expression of Dopamine-Related Genes in Four Human Brain Regions. Brain Sciences. 2020; 10(8):567. https://doi.org/10.3390/brainsci10080567
Chicago/Turabian StyleStanfill, Ansley Grimes, and Xueyuan Cao. 2020. "Expression of Dopamine-Related Genes in Four Human Brain Regions" Brain Sciences 10, no. 8: 567. https://doi.org/10.3390/brainsci10080567
APA StyleStanfill, A. G., & Cao, X. (2020). Expression of Dopamine-Related Genes in Four Human Brain Regions. Brain Sciences, 10(8), 567. https://doi.org/10.3390/brainsci10080567