
Functional Connectivity as an Index of Brain Changes Following a Unicycle Intervention: A Graph-Theoretical Network Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design and Procedure
2.3. MRI Data Acquisition
2.4. Preprocessing
2.4.1. Anatomical Data Preprocessing
2.4.2. Functional Data Preprocessing
2.5. Graph Theoretical Analysis
2.5.1. Node Definition
2.5.2. Edges
2.5.3. Thresholds
2.5.4. Graph Theoretical Parameters
Hubness
2.6. Statistical Analysis
2.7. Stability of Findings
3. Results
3.1. Descriptive Statistics
3.2. Intervention Effects
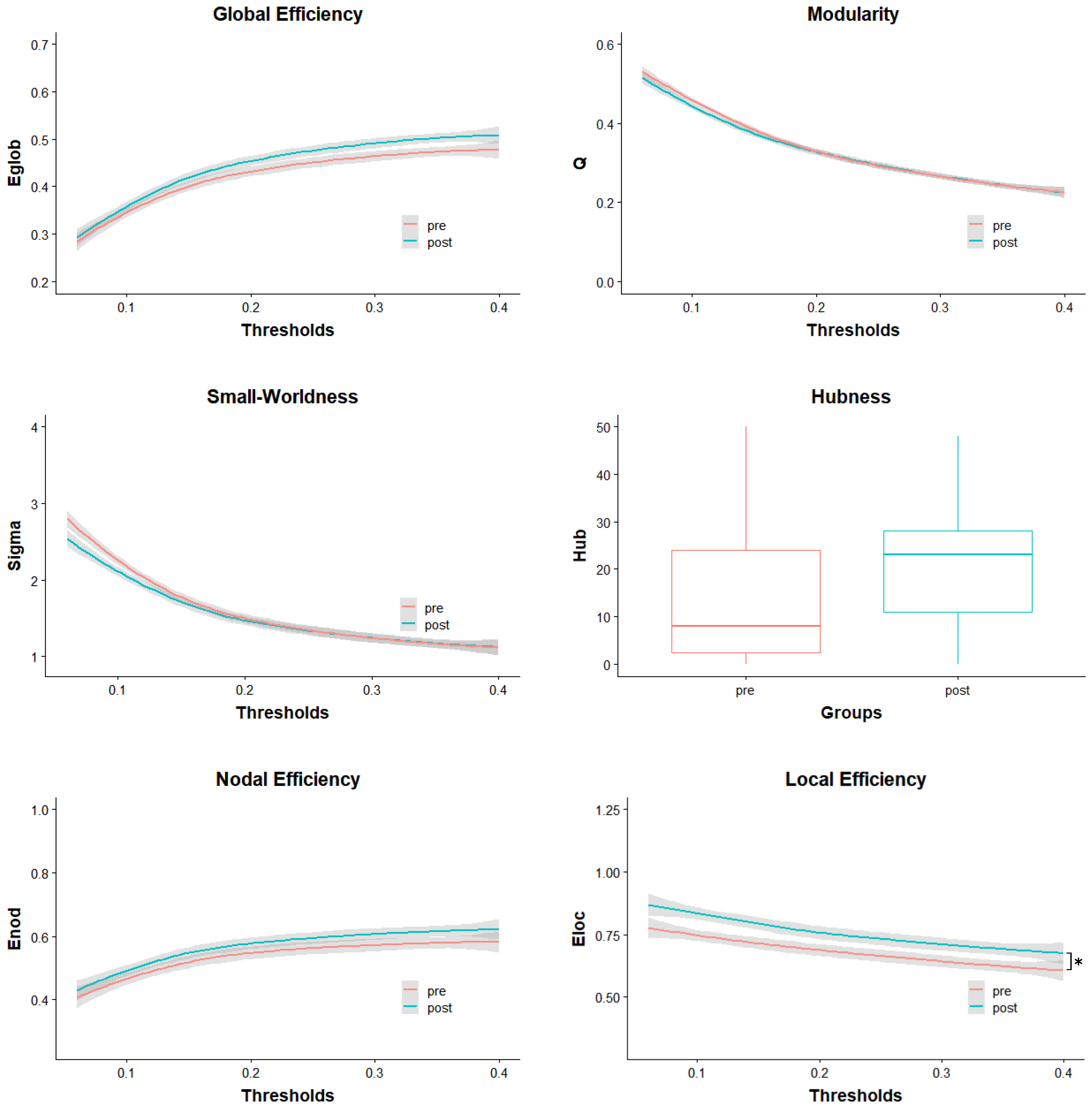
3.2.1. Local Effects
3.2.2. Global Effects
3.3. Stability of Findings
4. Discussion
4.1. Implications
4.2. Stability of Findings
4.3. Methodological Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | area under the curve |
| CSF | cerebrospinal fluid |
| GM | grey matter |
| WM | white matter |
| FD | framewise displacement |
| rSTG | right Superior Temporal Gyrus |
References
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.; Lipps, D.B. Motor control and aging: Links to age-related brain structural, functional, and biochemical effects. Neurosci. Biobehav. Rev. 2010, 34, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Baudry, S. Aging Changes the Contribution of Spinal and Corticospinal Pathways to Control Balance. Exerc. Sport Sci. Rev. 2016, 44, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Boisgontier, M.P.; Beets, I.A.; Duysens, J.; Nieuwboer, A.; Krampe, R.T.; Swinnen, S.P. Age-related differences in attentional cost associated with postural dual tasks: Increased recruitment of generic cognitive resources in older adults. Neurosci. Biobehav. Rev. 2013, 37, 1824–1837. [Google Scholar] [CrossRef] [PubMed]
- Papegaaij, S.; Taube, W.; Baudry, S.; Otten, E.; Hortobágyi, T. Aging causes a reorganization of cortical and spinal control of posture. Front. Aging Neurosci. 2014, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Allali, G.; van der Meulen, M.; Beauchet, O.; Rieger, S.W.; Vuilleumier, P.; Assal, F. The Neural Basis of Age-Related Changes in Motor Imagery of Gait: An fMRI Study. J. Gerontol. Ser. A 2013, 69, 1389–1398. [Google Scholar] [CrossRef]
- Ruffieux, J.; Mouthon, A.; Keller, M.; Mouthon, M.; Annoni, J.M.; Taube, W. Balance Training Reduces Brain Activity during Motor Simulation of a Challenging Balance Task in Older Adults: An fMRI Study. Front. Behav. Neurosci. 2018, 12, 10. [Google Scholar] [CrossRef]
- De Rond, V.; Orcioli-Silva, D.; Dijkstra, B.W.; de Xivry, J.J.O.; Pantall, A.; Nieuwboer, A. Compromised Brain Activity With Age During a Game-Like Dynamic Balance Task: Single- vs. Dual-Task Performance. Front. Aging Neurosci. 2021, 13, 657308. [Google Scholar] [CrossRef]
- Sehm, B.; Taubert, M.; Conde, V.; Weise, D.; Classen, J.; Dukart, J.; Draganski, B.; Villringer, A.; Ragert, P. Structural brain plasticity in Parkinson’s disease induced by balance training. Neurobiol. Aging 2014, 35, 232–239. [Google Scholar] [CrossRef]
- Rogge, A.K.; Röder, B.; Zech, A.; Nagel, V.; Hollander, K.; Braumann, K.M.; Hötting, K. Balance training improves memory and spatial cognition in healthy adults. Sci. Rep. 2017, 7, 5661. [Google Scholar] [CrossRef]
- Weber, B.; Koschutnig, K.; Schwerdtfeger, A.; Rominger, C.; Papousek, I.; Weiss, E.M.; Tilp, M.; Fink, A. Learning Unicycling Evokes Manifold Changes in Gray and White Matter Networks Related to Motor and Cognitive Functions. Sci. Rep. 2019, 9, 4324. [Google Scholar] [CrossRef]
- Baier, B.; Suchan, J.; Karnath, H.O.; Dieterich, M. Neural correlates of disturbed perception of verticality. Neurology 2012, 78, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A. An exploration of the role of the superior temporal gyrus in visual search and spatial perception using TMS. Brain 2004, 127, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Karnath, H.O.; Ferber, S.; Himmelbach, M. Spatial awareness is a function of the temporal not the posterior parietal lobe. Nature 2001, 411, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Karnath, H.O. New insights into the functions of the superior temporal cortex. Nat. Rev. Neurosci. 2001, 2, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Aminoff, E.M.; Kveraga, K.; Bar, M. The role of the parahippocampal cortex in cognition. Trends Cogn. Sci. 2013, 17, 379–390. [Google Scholar] [CrossRef]
- Zatorre, R.J.; Fields, R.D.; Johansen-Berg, H. Plasticity in gray and white: Neuroimaging changes in brain structure during learning. Nat. Neurosci. 2012, 15, 528–536. [Google Scholar] [CrossRef]
- Glasser, M.F.; Sotiropoulos, S.N.; Wilson, J.A.; Coalson, T.S.; Fischl, B.; Andersson, J.L.; Xu, J.; Jbabdi, S.; Webster, M.; Polimeni, J.R.; et al. The minimal preprocessing pipelines for the Human Connectome Project. NeuroImage 2013, 80, 105–124. [Google Scholar] [CrossRef]
- Esteban, O.; Markiewicz, C.J.; Goncalves, M.; DuPre, E.; Kent, J.D.; Salo, T.; Ciric, R.; Pinsard, B.; Blair, R.W.; Poldrack, R.A.; et al. fMRIPrep: A Robust Preprocessing Pipeline for Functional MRI. 2018. Available online: https://doi.org/10.5281/ZENODO.852659 (accessed on 2 June 2020).
- Esteban, O.; Markiewicz, C.J.; Blair, R.W.; Moodie, C.A.; Isik, A.I.; Erramuzpe, A.; Kent, J.D.; Goncalves, M.; DuPre, E.; Snyder, M.; et al. fMRIPrep: A robust preprocessing pipeline for functional MRI. Nat. Methods 2018, 16, 111–116. [Google Scholar] [CrossRef]
- Gorgolewski, K.; Burns, C.D.; Madison, C.; Clark, D.; Halchenko, Y.O.; Waskom, M.L.; Ghosh, S.S. Nipype: A Flexible, Lightweight and Extensible Neuroimaging Data Processing Framework in Python. Front. Neuroinformatics 2011, 5, 13. [Google Scholar] [CrossRef]
- Gorgolewski, K.J.; Esteban, O.; Markiewicz, C.J.; Burns, C.; Goncalves, M.; Jarecka, D.; Ziegler, E.; Berleant, S.; Ellis, D.G.; Pinsard, B.; et al. nipy/nipype: 1.5.1. 2018. Available online: https://doi.org/10.5281/ZENODO.596855 (accessed on 2 June 2020).
- Abraham, A.; Pedregosa, F.; Eickenberg, M.; Gervais, P.; Mueller, A.; Kossaifi, J.; Gramfort, A.; Thirion, B.; Varoquaux, G. Machine learning for neuroimaging with scikit-learn. Front. Neuroinform. 2014, 8, 14. [Google Scholar] [CrossRef]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 Bias Correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Avants, B.; Epstein, C.; Grossman, M.; Gee, J. Symmetric diffeomorphic image registration with cross-correlation: Evaluating automated labeling of elderly and neurodegenerative brain. Med. Image Anal. 2008, 12, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med. Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved Optimization for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.; Hyde, J. Software tools for analysis and visualization of fMRI data. NMR Biomed. 1997, 10, 171–178. [Google Scholar] [CrossRef]
- Greve, D.N.; Fischl, B. Accurate and robust brain image alignment using boundary-based registration. NeuroImage 2009, 48, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Mitra, A.; Laumann, T.O.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Methods to detect, characterize, and remove motion artifact in resting state fMRI. NeuroImage 2014, 84, 320–341. [Google Scholar] [CrossRef]
- Wang, J.; Wange, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A graph theoretical network analysis toolbox for imaging connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Breakspear, M. Graph analysis of the human connectome: Promise, progress, and pitfalls. NeuroImage 2013, 80, 426–444. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Bullmore, E.T. Network-based statistic: Identifying differences in brain networks. NeuroImage 2010, 53, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Fornito, A.; Zalesky, A.; Bullmore, E.T. Fundamentals of Brain Network Analysis; Elsevier: Amsterdam, The Netherlands, 2016; p. xvii, 476. [Google Scholar]
- van den Heuvel, M.P.; Stam, C.; Boersma, M.; Pol, H.H. Small-world and scale-free organization of voxel-based resting-state functional connectivity in the human brain. NeuroImage 2008, 43, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests, R Package Version 0.7.0. 2021. Available online: https://rpkgs.datanovia.com/rstatix/(accessed on 7 July 2022).
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for Descriptive Statistics, R Package Version 0.99.36. 2022. Available online: https://cran.r-project.org/web/packages/DescTools/index.html(accessed on 7 July 2022).
- Dosenbach, N.U.F.; Nardos, B.; Cohen, A.L.; Fair, D.A.; Power, J.D.; Church, J.A.; Nelson, S.M.; Wig, G.S.; Vogel, A.C.; Lessov-Schlaggar, C.N.; et al. Prediction of Individual Brain Maturity Using fMRI. Science 2010, 329, 1358–1361. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef]
- Rey, D.; Neuhäuser, M. Wilcoxon-Signed-Rank Test. In International Encyclopedia of Statistical Science; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1658–1659. [Google Scholar]
- Achard, S.; Bullmore, E. Efficiency and Cost of Economical Brain Functional Networks. PLoS Comput. Biol. 2007, 3, 1–10. [Google Scholar] [CrossRef]
- Latora, V.; Marchiori, M. Economic small-world behavior in weighted networks. Eur. Phys. J. B—Condens. Matter Complex Systems 2003, 32, 249–263. [Google Scholar] [CrossRef]
- Burdette, J.H.; Laurienti, P.J.; Espeland, M.A.; Morgan, A.; Telesford, Q.; Vechlekar, C.D.; Hayasaka, S.; Jennings, J.M.; Katula, J.A.; Kraft, R.A.; et al. Using network science to evaluate exercise-associated brain changes in older adults. Front. Aging Neurosci. 2010, 2, 23. [Google Scholar] [CrossRef]
- Passingham, R.E.; Rowe, J.B. Functional specialization. In A Short Guide to Brain Imaging; Oxford University Press: Oxford, UK, 2015; pp. 71–90. [Google Scholar] [CrossRef]
- Kitano, H. Biological robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef]
- Noppeney, U.; Friston, K.J.; Price, C.J. Degenerate neuronal systems sustaining cognitive functions. J. Anat. 2004, 205, 433–442. [Google Scholar] [CrossRef]
- Gallen, C.L.; D’Esposito, M. Brain Modularity: A Biomarker of Intervention-related Plasticity. Trends Cogn. Sci. 2019, 23, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Aurich, N.K.; Alves Filho, J.O.; Marques da Silva, A.M.; Franco, A.R. Evaluating the reliability of different preprocessing steps to estimate graph theoretical measures in resting state fMRI data. Front. Neurosci. 2015, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Mårtensson, G.; Pereira, J.B.; Mecocci, P.; Vellas, B.; Tsolaki, M.; Kłoszewska, I.; Soininen, H.; Lovestone, S.; Simmons, A.; Volpe, G.; et al. Stability of graph theoretical measures in structural brain networks in Alzheimer’s disease. Sci. Rep. 2018, 8, 11592. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, H.; Duan, Y.; Huang, J.; Ren, Z.; Ye, J.; Dong, H.; Shi, F.; Li, K.; Wang, J. Functional Brain Network Alterations in Clinically Isolated Syndrome and Multiple Sclerosis: A Graph-based Connectome Study. Radiology 2017, 282, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Honey, C.J.; Kötter, R. Identification and Classification of Hubs in Brain Networks. PLoS ONE 2007, 2, e1049. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, O.; Saarimäki, H.; Glerean, E.; Sams, M.; Saramäki, J. Consistency of Regions of Interest as nodes of fMRI functional brain networks. Netw. Neurosci. 2017, 1, 254–274. [Google Scholar] [CrossRef]
- Stanley, M.L.; Moussa, M.N.; Paolini, B.M.; Lyday, R.G.; Burdette, J.H.; Laurienti, P.J. Defining nodes in complex brain networks. Front. Comput. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Salehi, M.; Greene, A.S.; Karbasi, A.; Shen, X.; Scheinost, D.; Constable, R.T. There is no single functional atlas even for a single individual: Functional parcel definitions change with task. NeuroImage 2020, 208, 116366. [Google Scholar] [CrossRef]
- Van den Heuvel, M.P.; de Lange, S.C.; Zalesky, A.; Seguin, C.; Yeo, B.T.; Schmidt, R. Proportional thresholding in resting-state fMRI functional connectivity networks and consequences for patient-control connectome studies: Issues and recommendations. NeuroImage 2017, 152, 437–449. [Google Scholar] [CrossRef]


{kind=link}
| Total | Low | High | ||||
|---|---|---|---|---|---|---|
| t | p | t | p | t | p | |
| AAL-116 | ||||||
| Eloc | 4.25 | <0.001 | 4.38 | <0.001 | 3.96 | <0.001 |
| Enod | 1.73 | 0.097 | 1.73 | 0.099 | 1.72 | 0.099 |
| Eglob | 2.21 | 0.038 | 1.93 | 0.067 | 2.28 | 0.033 |
| Mod | −0.59 | 0.564 | −1.09 | 0.286 | 0.06 | 0.956 |
| SW | −0.84 | 0.409 | −1.06 | 0.302 | 0.07 | 0.943 |
| Hub | 1.90 | 0.070 | 2.02 | 0.056 | 0.53 | 0.599 |
| Dosenbach-160 | ||||||
| Eloc | 0.69 | 0.500 | 0.29 | 0.777 | 1.10 | 0.283 |
| Enod | 0.44 | 0.668 | 0.50 | 0.622 | 0.38 | 0.712 |
| Eglob | 1.83 | 0.080 | 1.79 | 0.088 | 1.51 | 0.146 |
| Mod | 0.03 | 0.979 | 0.20 | 0.847 | −0.18 | 0.856 |
| SW | 0.01 | 0.991 | 0.01 | 0.992 | 0.03 | 0.978 |
| Hub | 0.39 | 0.702 | 0.28 | 0.783 | 0.44 | 0.666 |
| Power-264 | ||||||
| Eloc | 0.04 | 0.971 | 0.44 | 0.665 | *136* | *0.964* |
| Enod | 0.38 | 0.707 | 0.32 | 0.749 | 0.45 | 0.657 |
| Eglob | 1.64 | 0.115 | 1.70 | 0.104 | 1.46 | 0.159 |
| Mod | −0.10 | 0.920 | <0.01 | 0.997 | −0.24 | 0.813 |
| SW | −0.41 | 0.690 | −0.40 | 0.690 | −0.38 | 0.710 |
| Hub | 0.77 | 0.449 | 0.74 | 0.465 | *108.5* | *0.378* |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riedmann, U.; Fink, A.; Weber, B.; Koschutnig, K. Functional Connectivity as an Index of Brain Changes Following a Unicycle Intervention: A Graph-Theoretical Network Analysis. Brain Sci. 2022, 12, 1092. https://doi.org/10.3390/brainsci12081092
Riedmann U, Fink A, Weber B, Koschutnig K. Functional Connectivity as an Index of Brain Changes Following a Unicycle Intervention: A Graph-Theoretical Network Analysis. Brain Sciences. 2022; 12(8):1092. https://doi.org/10.3390/brainsci12081092
Chicago/Turabian StyleRiedmann, Uwe, Andreas Fink, Bernhard Weber, and Karl Koschutnig. 2022. "Functional Connectivity as an Index of Brain Changes Following a Unicycle Intervention: A Graph-Theoretical Network Analysis" Brain Sciences 12, no. 8: 1092. https://doi.org/10.3390/brainsci12081092
APA StyleRiedmann, U., Fink, A., Weber, B., & Koschutnig, K. (2022). Functional Connectivity as an Index of Brain Changes Following a Unicycle Intervention: A Graph-Theoretical Network Analysis. Brain Sciences, 12(8), 1092. https://doi.org/10.3390/brainsci12081092