
Neural Research on Depth Perception and Stereoscopic Visual Fatigue in Virtual Reality


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Environment
2.3. Stimuli and Procedure
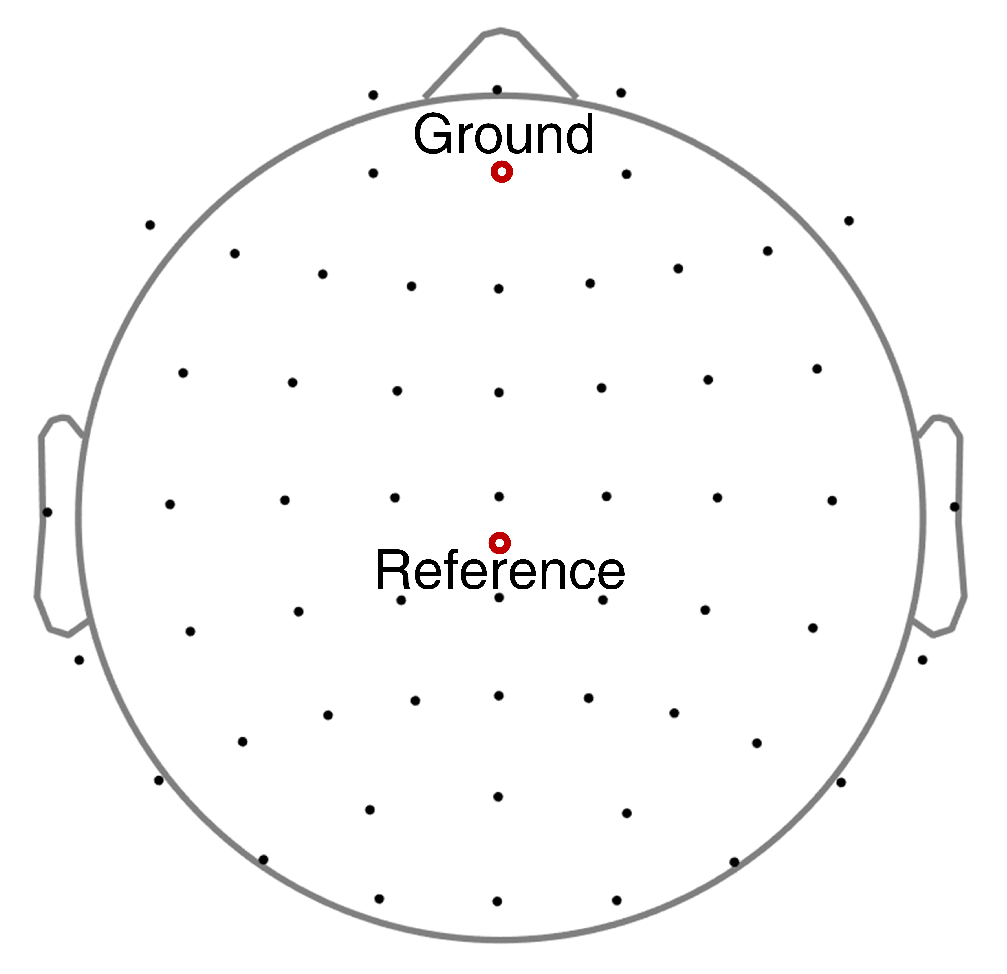
2.4. EEG Acquisition
2.5. EEG Data Analysis
2.5.1. EEG Data Pre-Processing
2.5.2. Time-Domain Analysis of VEPs
Disparity-Related Analysis
Block-Related Analysis
2.5.3. EEG cortical Source Localization
3. Results
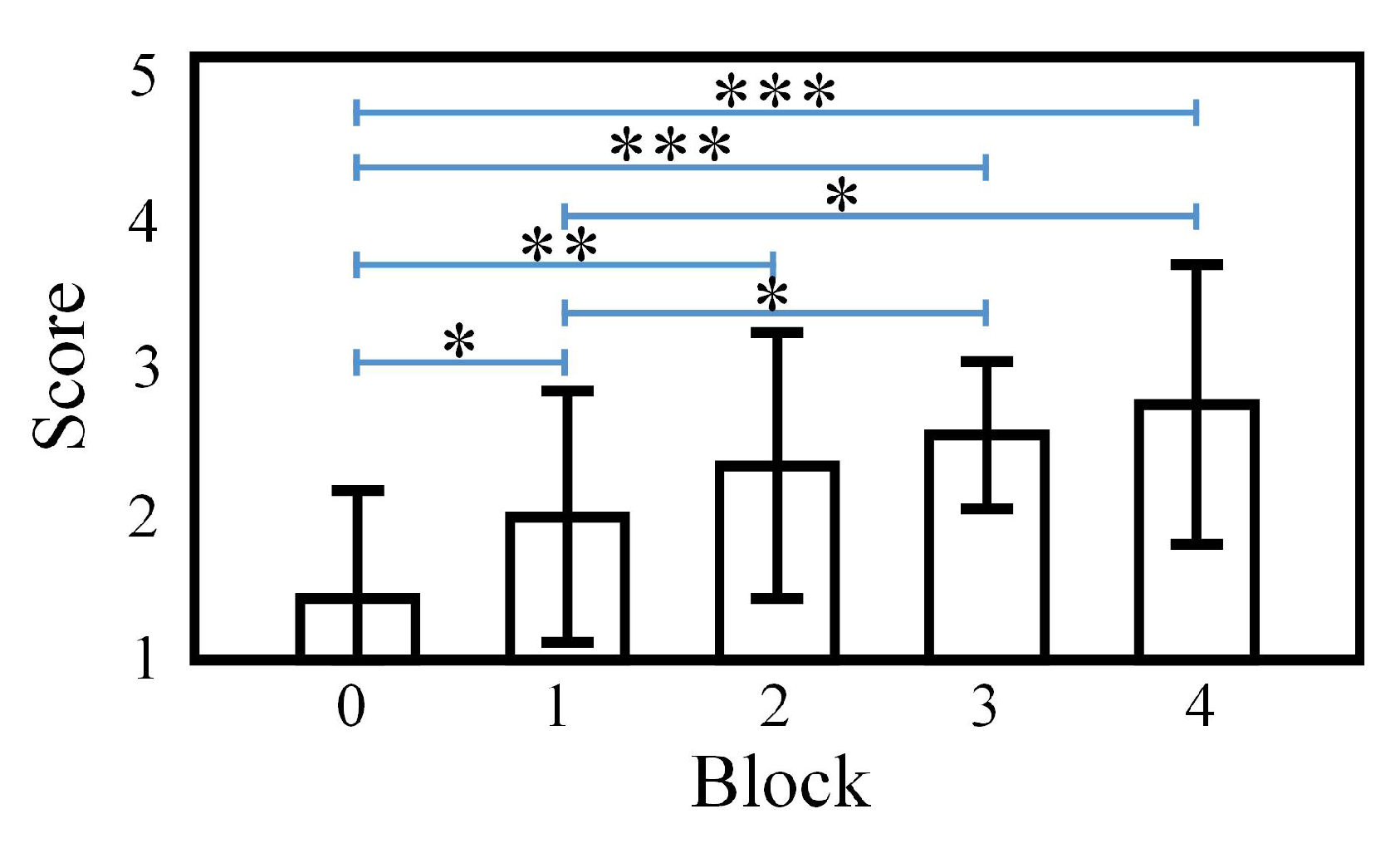
3.1. Subjective SVF Evaluation
3.2. Results of the Behavioral Task
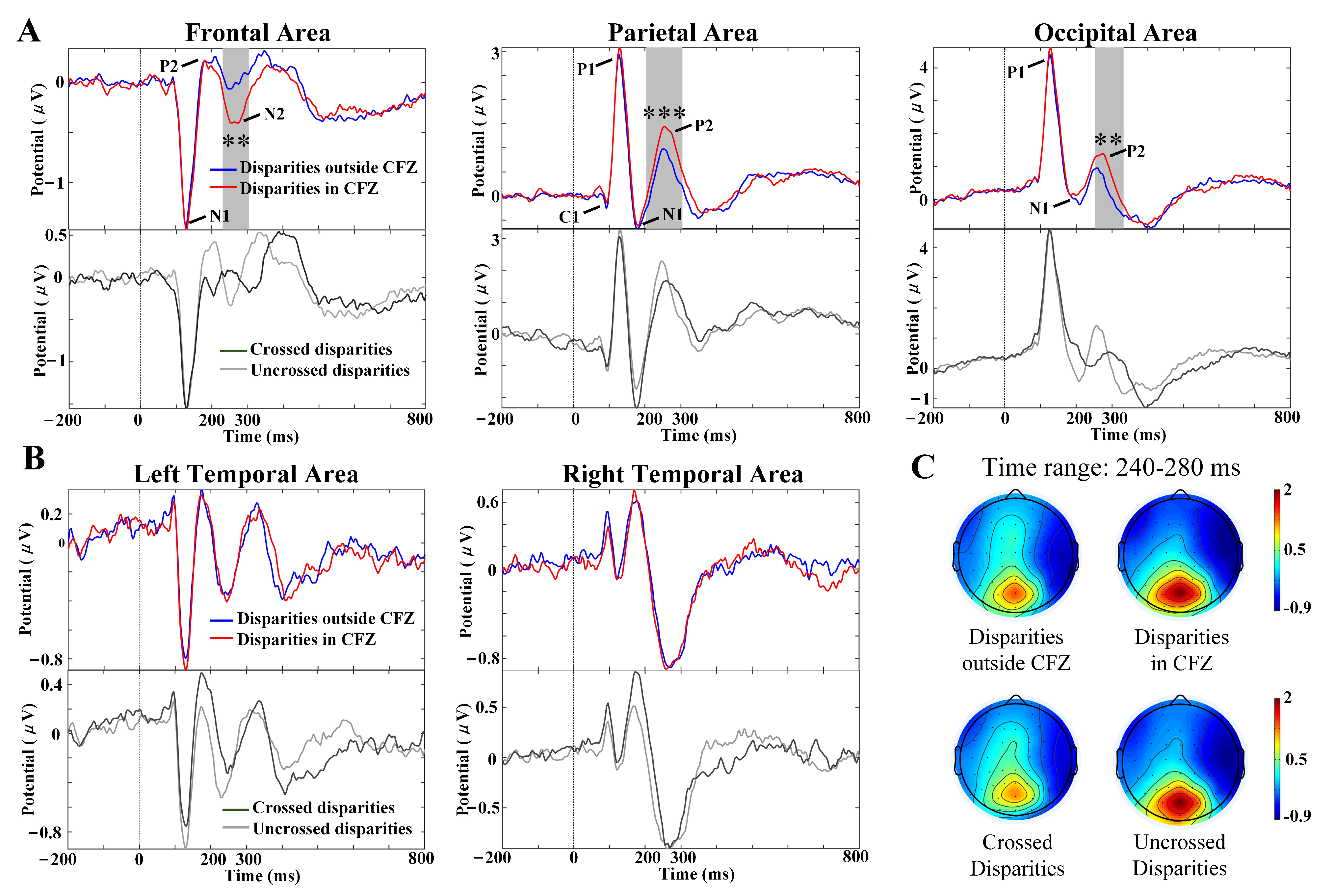
3.3. VEP Characteristics with Different Disparities
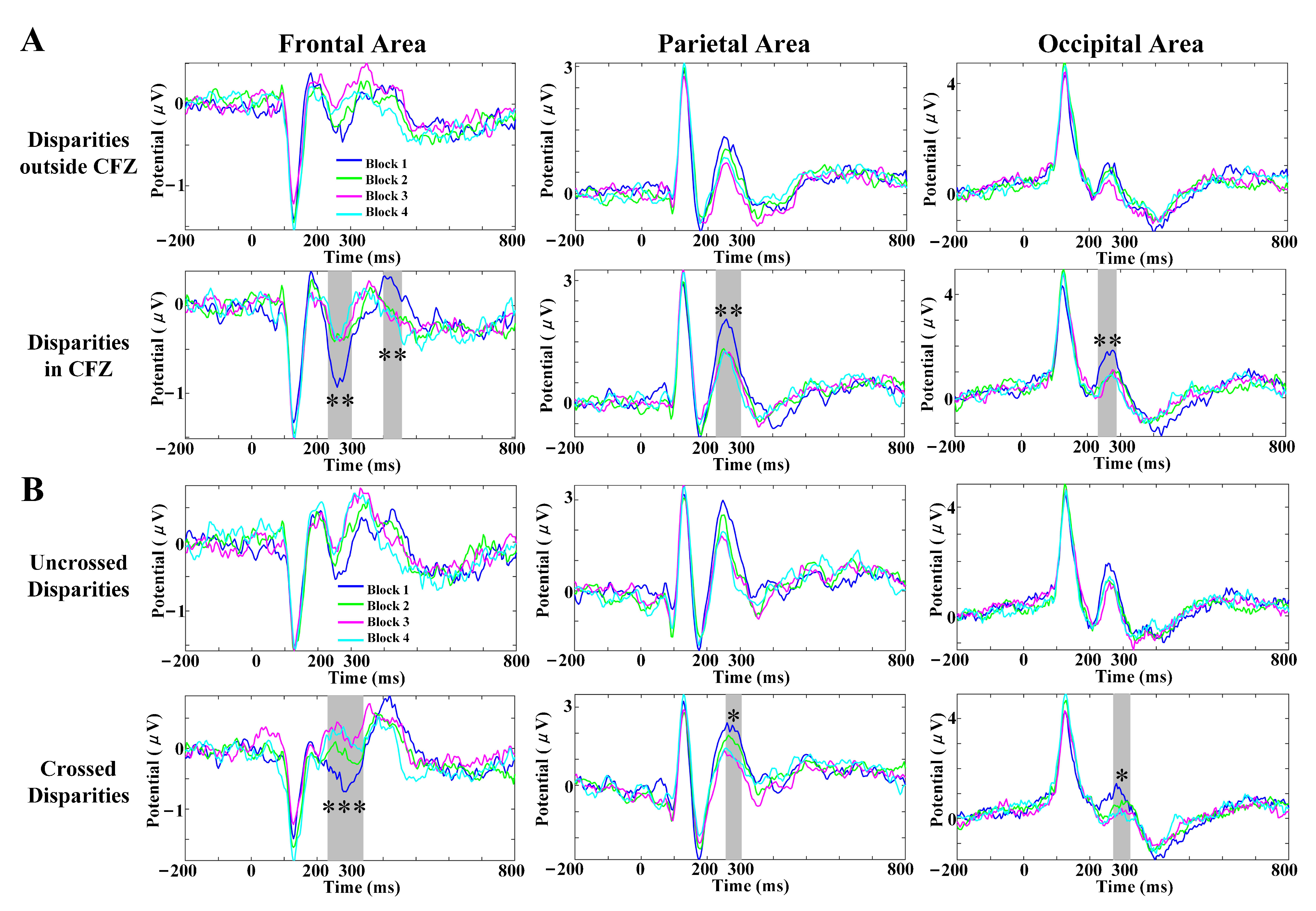
3.4. SVF Effects on Scalp VEP Components
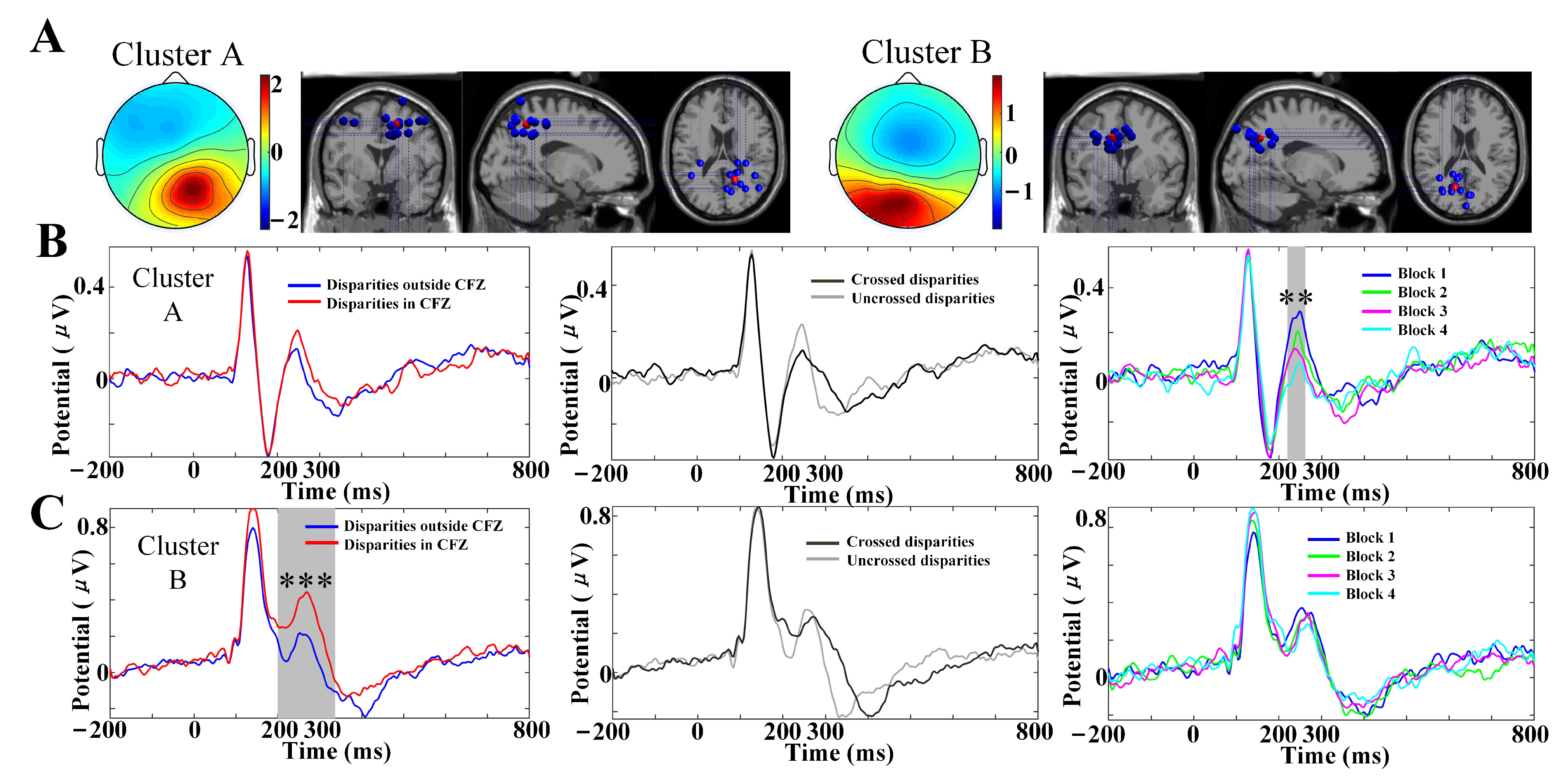
3.5. IC Clusters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheedy, J.E.; Hayes, J.; Engle, J. Is all Asthenopia the Same? Optom. Vis. Sci. 2003, 80, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Ukai, K.; Howarth, P.A. Visual fatigue caused by viewing stereoscopic motion images: Background, theories, and observations. Displays 2008, 29, 106–116. [Google Scholar] [CrossRef]
- Lambooij, M.; Ijsselsteijn, W.; Fortuin, M.; Heynderickx, I. Visual Discomfort and Visual Fatigue of Stereoscopic Displays: A Review. J. Imaging Sci. Technol. 2009, 53, 030201. [Google Scholar] [CrossRef]
- Zheng, F.; Hou, F.; Chen, R.; Mei, J.; Huang, P.; Chen, B.; Wang, Y. Investigation of the Relationship Between Subjective Symptoms of Visual Fatigue and Visual Functions. Front. Neurosci. 2021, 15, 1–11. [Google Scholar] [CrossRef]
- Blehm, C.; Vishnu, S.; Khattak, A.; Mitra, S.; Yee, R.W. Computer vision syndrome: A review. Surv. Ophthalmol. 2005, 50, 253–262. [Google Scholar] [CrossRef]
- Sheppard, A.L.; Wolffsohn, J.S. Digital eye strain: Prevalence, measurement and amelioration. BMJ Open Ophthalmol. 2018, 3. [Google Scholar] [CrossRef]
- Lee, C.C.; Chiang, H.S.; Hsiao, M.H. Effects of screen size and visual presentation on visual fatigue based on regional brain wave activity. J. Supercomput. 2021, 77, 4831–4851. [Google Scholar] [CrossRef]
- Hoffman, D.M.; Girshick, A.R.; Akeley, K.; Banks, M.S. Vergence-accommodation conflicts hinder visual performance and cause visual fatigue. J. Vis. 2008, 8, 1–30. [Google Scholar] [CrossRef]
- Kramida, G. Resolving the Vergence-Accommodation Conflict in Head-Mounted Displays. IEEE Trans. Vis. Comput. Graph. 2016, 22, 1912–1931. [Google Scholar] [CrossRef]
- Koulieris, G.A.; Bui, B.; Banks, M.S.; Drettakis, G. Accommodation and comfort in head-mounted displays. ACM Trans. Graph. 2017, 36, 87. [Google Scholar] [CrossRef]
- Do, H.; Kim, Y.M.; Min, S.W. Focus-free head-mounted display based on Maxwellian view using retroreflector film. Appl. Opt. 2019, 58, 2882. [Google Scholar] [CrossRef] [PubMed]
- Picton, T.W.; Bentin, S.; Berg, P.; Donchin, E.; Hillyard, S.A.; Johnson, R.; Miller, G.A.; Ritter, W.; Ruchkin, D.S.; Rugg, M.D.; et al. Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology 2000, 37, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Klistorner, A.; Graham, S.L. Role of Multifocal Visually Evoked Potential as a Biomarker of Demyelination, Spontaneous Remyelination, and Myelin Repair in Multiple Sclerosis. Front. Neurosci. 2021, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kasai, T.; Morotomi, T.; Katayama, J.; Kumada, T. Attending to a location in three-dimensional space modulates early ERPs. Cogn. Brain Res. 2003, 17, 273–285. [Google Scholar] [CrossRef]
- Duncan, C.C.; Barry, R.J.; Connolly, J.F.; Fischer, C.; Michie, P.T.; Näätänen, R.; Polich, J.; Reinvang, I.; Van Petten, C. Event-related potentials in clinical research: Guidelines for eliciting, recording, and quantifying mismatch negativity, P300, and N400. Clin. Neurophysiol. 2009, 120, 1883–1908. [Google Scholar] [CrossRef] [PubMed]
- Kremers, J.; McKeefry, D.J.; Murray, I.J.; Parry, N.R. Developments in non-invasive visual electrophysiology. Vis. Res. 2020, 174, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Şahinoǧlu, B. The effect of disparity change on binocular visual evoked potential parameters elicited by convergent dynamic random-dot stereogram stimuli in humans. Eur. J. Appl. Physiol. 2002, 88, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Şahinoǧlu, B. Depth-related visually evoked potentials by dynamic random-dot stereograms in humans: Negative correlation between the peaks elicited by convergent and divergent disparities. Eur. J. Appl. Physiol. 2004, 91, 689–697. [Google Scholar] [CrossRef]
- Séverac Cauquil, A.; Trotter, Y.; Taylor, M.J. At what stage of neural processing do perspective depth cues make a difference? Exp. Brain Res. 2006, 170, 457–463. [Google Scholar] [CrossRef]
- Omoto, S.; Kuroiwa, Y.; Otsuka, S.; Baba, Y.; Wang, C.; Li, M.; Mizuki, N.; Ueda, N.; Koyano, S.; Suzuki, Y. P1 and P2 components of human visual evoked potentials are modulated by depth perception of 3-dimensional images. Clin. Neurophysiol. 2010, 121, 386–391. [Google Scholar] [CrossRef]
- Gao, F.; Cao, B.; Cao, Y.; Li, F.; Li, H. Electrophysiological evidence of separate pathways for the perception of depth and 3D objects. Int. J. Psychophysiol. 2015, 96, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Pegna, A.J.; Darque, A.; Roberts, M.V.; Leek, E.C. Effects of stereoscopic disparity on early ERP components during classification of three-dimensional objects. Q. J. Exp. Psychol. 2018, 71, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Wang, D.; Chiu, S.C.; Liu, Y. Investigate the 3D Visual Fatigue Using Modified Depth-Related Visual Evoked Potential Paradigm. IEEE Trans. Neural Syst. Rehabil. Eng. 2020, 28, 2794–2804. [Google Scholar] [CrossRef] [PubMed]
- Regan, D.; Beverley, K.I. Electrophysiological Evidence for Existence of Neurones sensitive to Direction of Depth Movement. Nature 1973, 246, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Julesz, B.; Kropfl, W.; Petrig, B. Large evoked potentials to dynamic random-dot correlograms and stereograms permit quick determination of stereopsis. Proc. Natl. Acad. Sci. USA 1980, 77, 2348–2351. [Google Scholar] [CrossRef]
- Trommer, B.L.; Hoeppner, J.A.B.; Lorber, R.; Armstrong, K.J. The Go—No-Go paradigm in attention deficit disorder. Ann. Neurol. 1988, 24, 610–614. [Google Scholar] [CrossRef]
- Gomez, P.; Ratcliff, R.; Perea, M. A model of the go/no-go task. J. Exp. Psychol. Gen. 2007, 136, 389. [Google Scholar] [CrossRef]
- Rokszin, A.A.; Győri-Dani, D.; Nyul, L.G.; Csifcsák, G. Electrophysiological correlates of top-down effects facilitating natural image categorization are disrupted by the attenuation of low spatial frequency information. Int. J. Psychophysiol. 2016, 100, 19–27. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Mullen, T.R.; Kothe, C.A.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.P.; Cauwenberghs, G. Real-time neuroimaging and cognitive monitoring using wearable dry EEG. IEEE Trans. Biomed. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.H.; Pion-Tonachini, L.; Palmer, J.; Miyakoshi, M.; Makeig, S.; Jung, T.P. Modeling brain dynamic state changes with adaptive mixture independent component analysis. NeuroImage 2018, 183, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kim, J.; Hoffman, D.M.; Banks, M.S. The zone of comfort: Predicting visual discomfort with stereo displays. J. Vis. 2011, 11, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Palmer, J.; Onton, J.; Oostenveld, R.; Makeig, S. Independent EEG sources are dipolar. PLoS ONE 2012, 7, e30135. [Google Scholar] [CrossRef]
- Peterson, S.M.; Ferris, D.P. Group-level cortical and muscular connectivity during perturbations to walking and standing balance. NeuroImage 2019, 198, 93–103. [Google Scholar] [CrossRef]
- Kaneko, N.; Yokoyama, H.; Masugi, Y.; Watanabe, K.; Nakazawa, K. Phase dependent modulation of cortical activity during action observation and motor imagery of walking: An EEG study. NeuroImage 2021, 225, 117486. [Google Scholar] [CrossRef]
- Bruin, K.J.; Wijers, A.A. Inhibition, response mode, and stimulus probability: A comparative event-related potential study. Clin. Neurophysiol. 2002, 113, 1172–1182. [Google Scholar] [CrossRef]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2008, 45, 152–170. [Google Scholar] [CrossRef]
- Falkenstein, M.; Hoormann, J.; Hohnsbein, J. ERP components in Go/Nogo tasks and their relation to inhibition. Acta Psychol. 1999, 101, 267–291. [Google Scholar] [CrossRef]
- Dunn, B.R.; Dunn, D.A.; Languis, M.; Andrews, D. The relation of ERP components to complex memory processing. Brain Cogn. 1998, 36, 355–376. [Google Scholar] [CrossRef]
- Lefebvre, C.D.; Marchand, Y.; Eskes, G.A.; Connolly, J.F. Assessment of working memory abilities using an event-related brain potential (ERP)-compatible digit span backward task. Clin. Neurophysiol. 2005, 116, 1665–1680. [Google Scholar] [CrossRef] [PubMed]
- Vanduffel, W.; Fize, D.; Peuskens, H.; Denys, K.; Sunaert, S.; Todd, J.T.; Orban, G.A. Extracting 3D from motion: Differences in human and monkey intraparietal cortex. Science 2002, 298, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Galletti, C.; Fattori, P. Neuronal mechanisms for detection of motion in the field of view. Neuropsychologia 2003, 41, 1717–1727. [Google Scholar] [CrossRef]
- Wang, X.; Yao, L.; Zhao, Y.; Xing, L.; Qian, Z.; Li, W.; Yang, Y. Effects of disparity on visual discomfort caused by short-term stereoscopic viewing based on electroencephalograph analysis. Biomed. Eng. Online 2018, 17, 166. [Google Scholar] [CrossRef] [PubMed]
- Vaina, L.M. Selective impairment of visual motion interpretation following lesions of the right occipito-parietal area in humans. Biol. Cybern. 1989, 61, 347–359. [Google Scholar] [CrossRef]
- Cumming, B.G.; DeAngelis, G.C. The physiology of stereopsis. Annu. Rev. Neurosci. 2001, 24, 203–238. [Google Scholar] [CrossRef]
- Gonzalez, F.; Perez, R. Neural mechanisms underlying stereoscopic vision. Prog. Neurobiol. 1998, 55, 191–224. [Google Scholar] [CrossRef]
- Parker, A.J. Binocular depth perception and the cerebral cortex. Nat. Rev. Neurosci. 2007, 8, 379–391. [Google Scholar] [CrossRef]
- Orban, G.A. The extraction of 3D shape in the visual system of human and nonhuman primates. Annu. Rev. Neurosci. 2011, 34, 361–368. [Google Scholar] [CrossRef]
- Welchman, A.E. The Human Brain in Depth: How We See in 3D. Annu. Rev. Vis. Sci. 2016, 2, 345–376. [Google Scholar] [CrossRef] [Green Version]
- Norcia, A.M.; Suiter, E.E.; Tyler, C.W. Electrophysiological evidence for the existence of coarse and fine disparity mechanisms in human. Vis. Res. 1985, 25, 1603–1611. [Google Scholar] [CrossRef]
- Katsuki, F.; Constantinidis, C. Unique and shared roles of the posterior parietal and dorsolateral prefrontal cortex in cognitive functions. Front. Integr. Neurosci. 2012, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, F.; Constantinidis, C. Bottom-up and top-down attention: Different processes and overlapping neural systems. Neuroscientist 2014, 20, 509–521. [Google Scholar] [CrossRef]
- Puig, M.S.; Pallarés, J.M.; Zapata, L.P.; Puigcerver, L.; Cañete, J.; Supèr, H. Attentional selection accompanied by eye vergence as revealed by event-related brain potentials. PLoS ONE 2016, 11, e0167646. [Google Scholar] [CrossRef]
- Clower, D.M.; West, R.A.; Lynch, J.C.; Strick, P.L. The inferior parietal lobule is the target of output from the superior colliculus, hippocampus, and cerebellum. J. Neurosci. 2001, 21, 6283–6291. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Choi, S.; Sohn, K. Effect of vergence-accommodation conflict and parallax difference on binocular fusion for random dot stereogram. IEEE Trans. Circuits Syst. Video Technol. 2012, 22, 811–816. [Google Scholar] [CrossRef]
- Templin, K.; Didyk, P.; Myszkowski, K.; Hefeeda, M.M.; Seidel, H.P.; Matusik, W. Modeling and optimizing eye vergence response to stereoscopic cuts. ACM Trans. Graph. 2014, 33, 1–8. [Google Scholar] [CrossRef]
- Tanaka, M.; Watanabe, Y. Supraspinal regulation of physical fatigue. Neurosci. Biobehav. Rev. 2012, 36, 727–734. [Google Scholar] [CrossRef]
- Anderson, A.J.; Ren, P.; Baran, T.M.; Zhang, Z.; Lin, F. Insula and putamen centered functional connectivity networks reflect healthy agers’ subjective experience of cognitive fatigue in multiple tasks. Cortex 2019, 119, 428–440. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [Green Version]
- Margulies, D.S.; Vincent, J.L.; Kelly, C.; Lohmann, G.; Uddin, L.Q.; Biswal, B.B.; Villringer, A.; Castellanos, F.X.; Milham, M.P.; Petrides, M. Precuneus shares intrinsic functional architecture in humans and monkeys. Proc. Natl. Acad. Sci. USA 2009, 106, 20069–20074. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, D.J.; Saleem, K.S.; Baker, C.I.; Mishkin, M. A new neural framework for visuospatial processing. Nat. Rev. Neurosci. 2011, 12, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, M.J.; Coyle, J.T.; Rowland, L.P. Neurobiology of Brain Disorders: Biological Basis of Neurological and Psychiatric Disorders; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Channel | Disparity | Peak Amplitude (V) | Latency (ms) | Repeated-Measures ANOVA Analysis (Peak Amplitude) | Repeated-Measures ANOVA Analysis (Latency) |
|---|---|---|---|---|---|
| Pz | Crossed | 1.848 (1.744) | 275.89 (28.92) | F(1,69) = 3.531; | F(1,69) = 7.944; |
| Uncrossed | 2.064 (1.743) | 266.83 (22.98) | p = 0.0645 | p = 0.006 | |
| POz | Crossed | 2.567 (1.909) | 278.91 (30.52) | F(1,69) = 8.317; | F(1,69) = 19.802; |
| Uncrossed | 3.113 (2.119) | 263.43 (19.51) | p = 0.005 | p < 0.0001 | |
| Oz | Crossed | 1.989 (1.870) | 290.80 (32.46) | F(1,69) = 12.312; | F(1,69) = 39.133; |
| Uncrossed | 2.533 (1.709) | 266.06 (23.32) | p = 0.0008 | p < 0.0001 |
| Channel | Disparity | Peak Amplitude (V) | Latency (ms) | Repeated-Measures ANOVA Analysis (Peak Amplitude) | Repeated-Measures ANOVA Analysis (Latency) | Multiple Comparisons (Peak Amplitude) | Multiple Comparisons (Latency) |
|---|---|---|---|---|---|---|---|
| Pz | Uncrossed outside CFZ | 2.029 (1.913) | 273.29 (30.66) | ||||
| Uncrossed in CFZ | 2.108 (1.704) | 266.71 (21.17) | F(3,81) = 1.287; | F(3,81) = 1.272; | |||
| Crossed in CFZ | 1.978 (1.775) | 270.71 (27.09) | p = 0.285 | p = 0.289 | |||
| Crossed outside CFZ | 1.765 (1.874) | 276.43 (29.92) | |||||
| POz | Uncrossed outside CFZ | 3.128 (2.291) | 267.57 (25.45) | ||||
| Uncrossed in CFZ | 3.203 (1.962) | 261.00 (13.18) | F(2.127,57.441) = 3.697; | F(1.923,51.921) = 4.965; | p(B,D) = 0.038 | p(A,C) = 0.057 | |
| Crossed in CFZ | 2.838 (1.879) | 278.29 (22.90) | p = 0.029 | p = 0.012 | p(B,C) = 0.0004 | ||
| Crossed outside CFZ | 2.389 (1.940) | 277.29 (37.56) | |||||
| Oz | Uncrossed outside CFZ | 2.477 (1.536) | 270.86 (15.55) | p(A,C) = 0.002 | |||
| Uncrossed in CFZ | 2.641 (1.899) | 264.57 (31.17) | F(3,81) = 4.376; | F(2.376,64.164) = 7.336; | p(A,D) = 0.051 | p(B,C) = 0.006 | |
| Crossed in CFZ | 2.138 (1.666) | 289.57 (24.01) | p = 0.007 | p = 0.0007 | p(B,D) = 0.055 | p(B,D) = 0.020 | |
| Crossed outside CFZ | 1.817 (2.419) | 290.29 (41.80) |
| Cluster | Location of Cluster Centroid (Brodmann Area) | Cluster Centroid Coordinates (x, y, z) | Number of Participants (Ps) and Ics | VEPs Components |
|---|---|---|---|---|
| A | Precuneus (BA7) | 12, −53, 51 | 13 Ps, 14 ICs | C1, P1, N1, P2 |
| B | Precuneus (BA7) | −14, −67, 37 | 10 Ps, 14 ICs | C1, P1, N1, P2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, M.; Yue, K.; Hu, H.; Lu, K.; Han, Y.; Chen, S.; Liu, Y. Neural Research on Depth Perception and Stereoscopic Visual Fatigue in Virtual Reality. Brain Sci. 2022, 12, 1231. https://doi.org/10.3390/brainsci12091231
Guo M, Yue K, Hu H, Lu K, Han Y, Chen S, Liu Y. Neural Research on Depth Perception and Stereoscopic Visual Fatigue in Virtual Reality. Brain Sciences. 2022; 12(9):1231. https://doi.org/10.3390/brainsci12091231
Chicago/Turabian StyleGuo, Mei, Kang Yue, Haochen Hu, Kai Lu, Yu Han, Shanshan Chen, and Yue Liu. 2022. "Neural Research on Depth Perception and Stereoscopic Visual Fatigue in Virtual Reality" Brain Sciences 12, no. 9: 1231. https://doi.org/10.3390/brainsci12091231
APA StyleGuo, M., Yue, K., Hu, H., Lu, K., Han, Y., Chen, S., & Liu, Y. (2022). Neural Research on Depth Perception and Stereoscopic Visual Fatigue in Virtual Reality. Brain Sciences, 12(9), 1231. https://doi.org/10.3390/brainsci12091231