
Effective Connectivity Evaluation of Resting-State Brain Networks in Alzheimer’s Disease, Amnestic Mild Cognitive Impairment, and Normal Aging: An Exploratory Study
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Demographic and Clinical Characteristics
2.1.1. Inclusion Criteria
2.1.2. Exclusion Criteria
2.2. MRI Acquisition
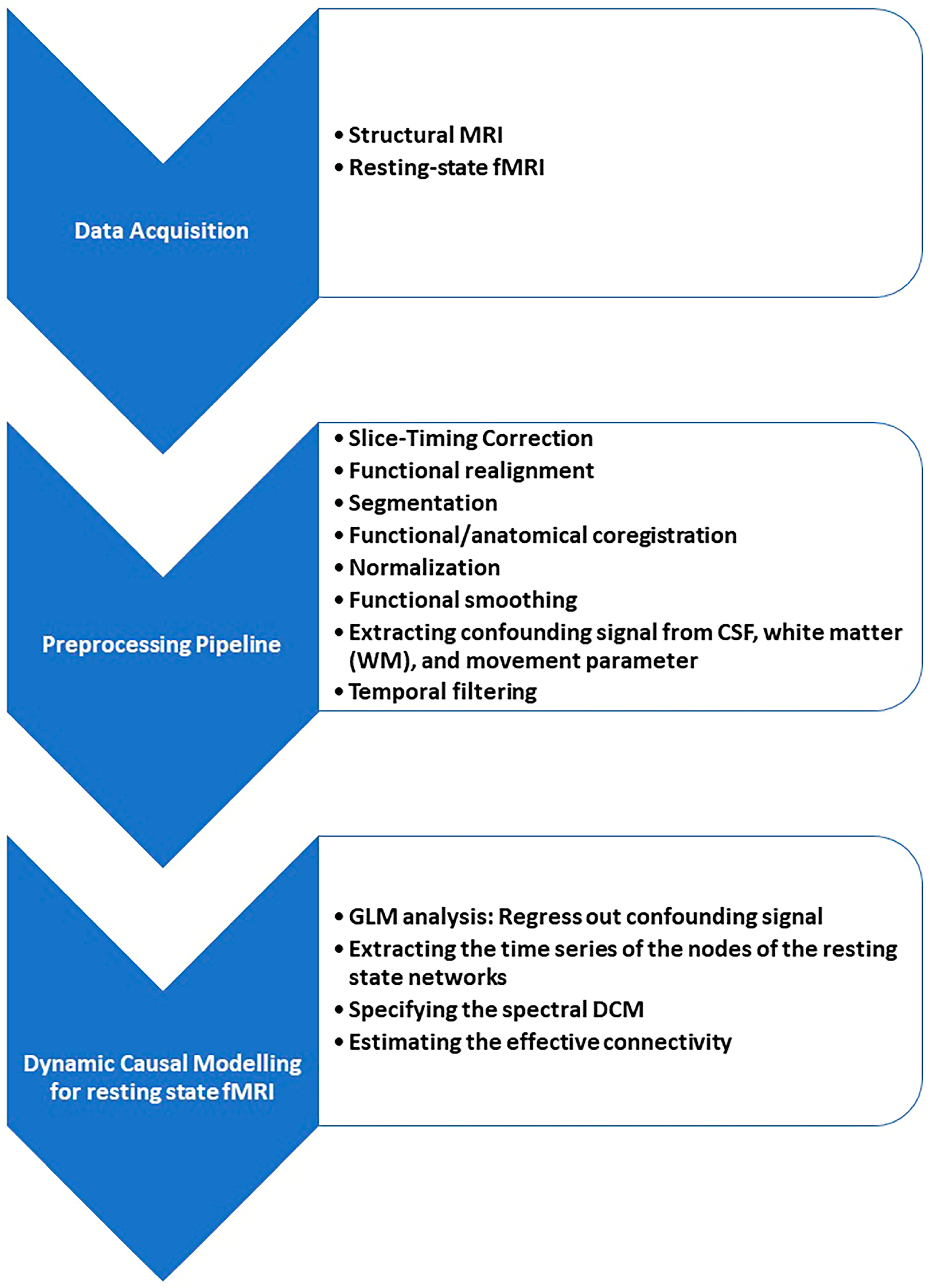
2.3. Pre-Processing
2.4. Resting-State Analysis
2.4.1. Independent Component Analyses (Group-ICA)
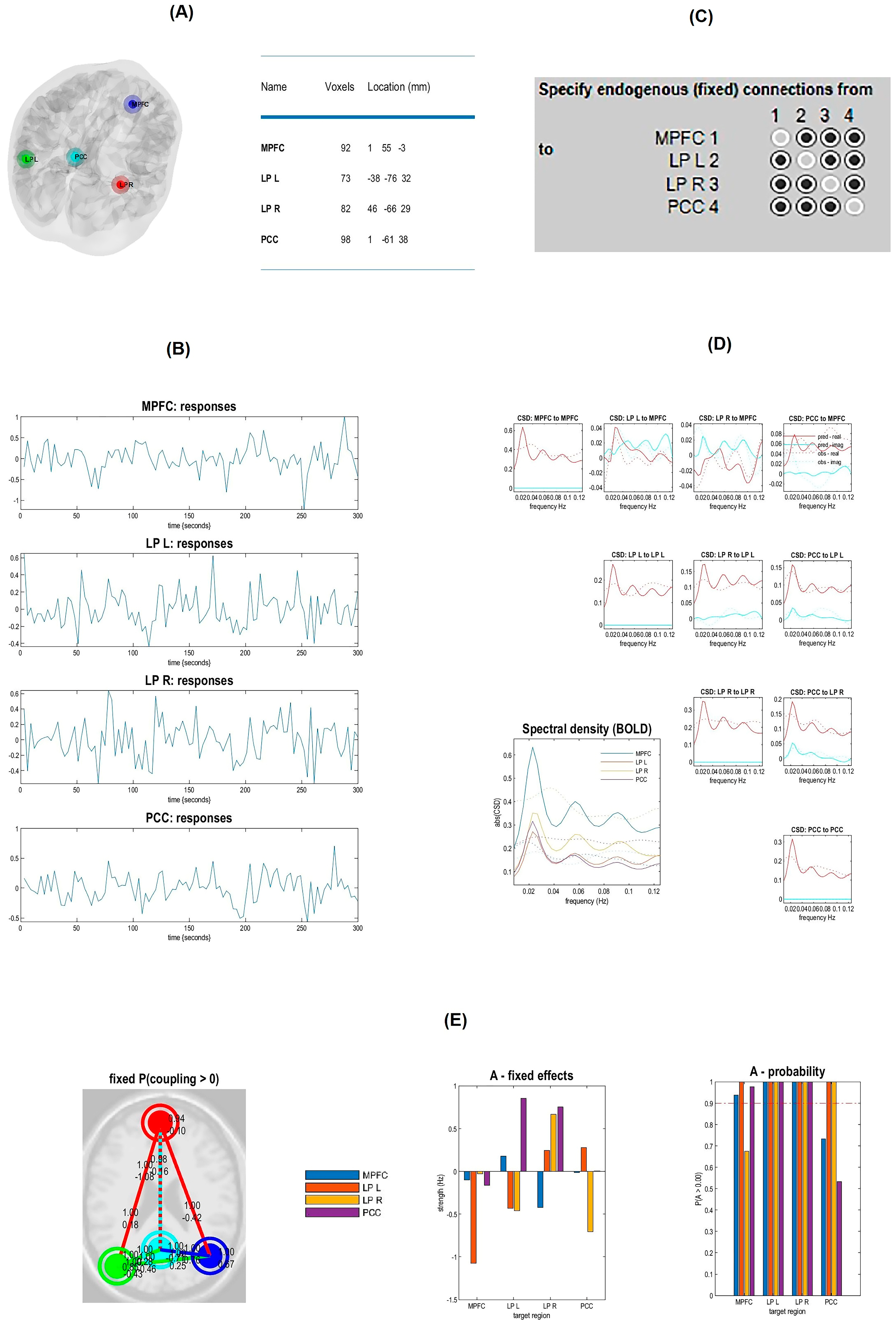
2.4.2. Spectral Dynamic Causal Modeling Analysis
2.5. Statistical Analysis
3. Results
3.1. Result of Demographic and Clinical Characteristics
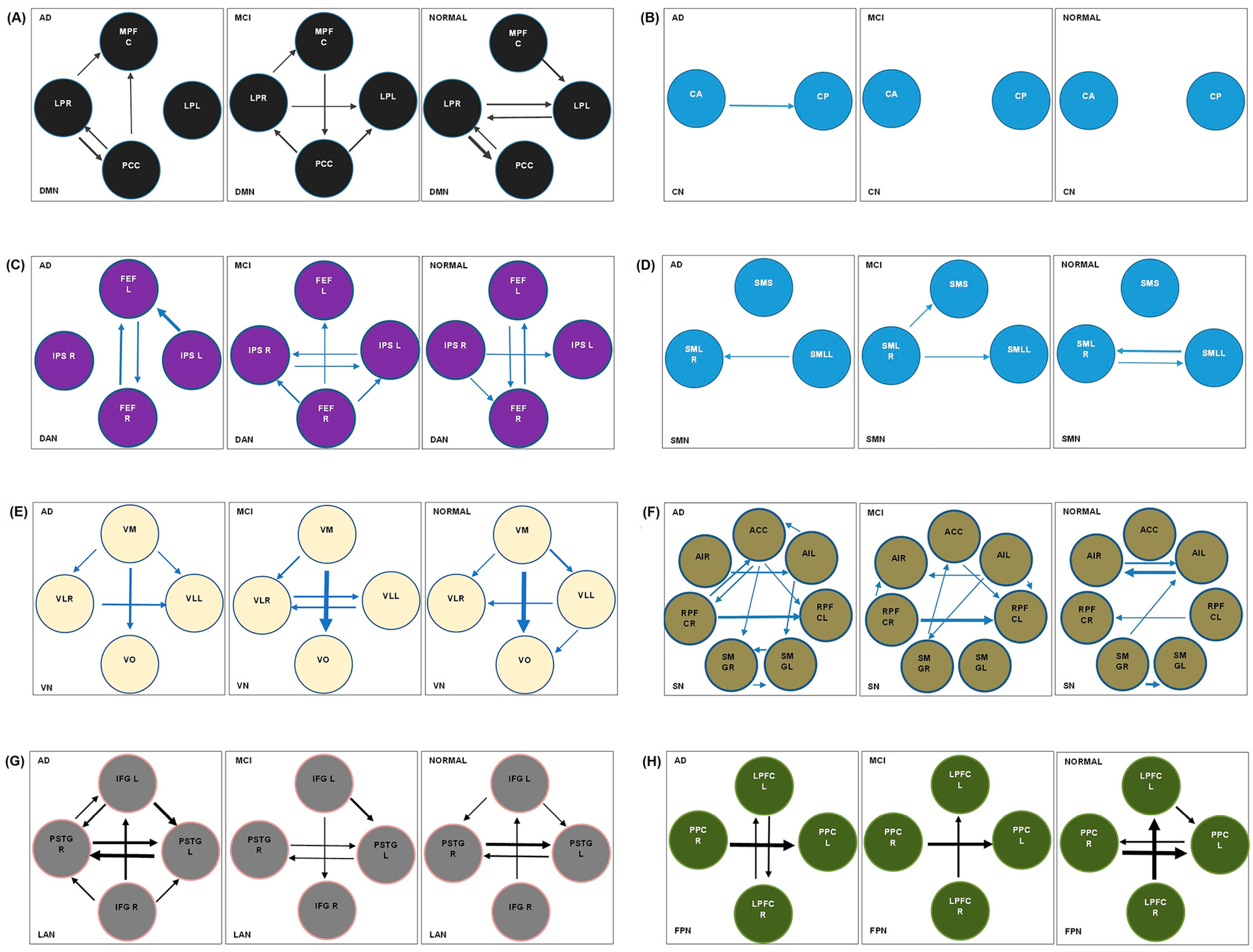
3.2. Results of spDCM Analysis
3.2.1. Default Mode Network
3.2.2. Cerebellar Network
3.2.3. Dorsal Attention Network
3.2.4. Sensorimotor Network
3.2.5. Visual Network
3.2.6. Salience Network
3.2.7. Language Network
3.2.8. Frontoparietal Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mirzaei, G.; Adeli, H. Machine learning techniques for diagnosis of alzheimer disease, mild cognitive disorder, and other types of dementia. Biomed. Signal Process. Control 2022, 72, 103293. [Google Scholar] [CrossRef]
- Soman, S.M.; Raghavan, S.; Rajesh, P.; Mohanan, N.; Thomas, B.; Kesavadas, C.; Menon, R.N. Does resting state functional connectivity differ between mild cognitive impairment and early Alzheimer’s dementia? J. Neurol. Sci. 2020, 418, 117093. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Liao, Z.; Tan, Y.; Qiu, Y.; Zhu, J.; Han, Z.; Wang, J.; Wang, X.; Wang, H.; Chen, Y. High-sensitivity neuroimaging biomarkers for the identification of amnestic mild cognitive impairment based on resting-state fMRI and a triple network model. Brain Imaging Behav. 2019, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, G.D.F. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the Global Burden of Disease Study 2019. Lancet Public Health 2022, 7, e105–e125. [Google Scholar]
- Small, G.W. Early diagnosis of Alzheimer’s disease: Update on combining genetic and brain-imaging measures. Dialogues Clin. Neurosci. 2022, 2, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhuo, Z.; Wei, L.; Ma, Z.; Li, Z.; Li, H.; Initiative, A.s.D.N. Altered temporal organization of brief spontaneous brain activities in patients with Alzheimer’s disease. Neuroscience 2020, 425, 1–11. [Google Scholar] [CrossRef]
- Liu, R.-M. Aging, cellular senescence, and Alzheimer’s disease. Int. J. Mol. Sci. 2022, 23, 1989. [Google Scholar] [CrossRef]
- Wang, J.; Khosrowabadi, R.; Ng, K.K.; Hong, Z.; Chong, J.S.X.; Wang, Y.; Chen, C.-Y.; Hilal, S.; Venketasubramanian, N.; Wong, T.Y. Alterations in brain network topology and structural-functional connectome coupling relate to cognitive impairment. Front. Aging Neurosci. 2018, 10, 404. [Google Scholar] [CrossRef]
- Scheltens, P. Imaging in Alzheimer’s disease. Dialogues Clin. Neurosci. 2022, 11, 191–199. [Google Scholar] [CrossRef]
- Kazemi-Harikandei, S.-Z.; Shobeiri, P.; Jelodar, M.-R.S.; Tavangar, S.M. Effective Connectivity in Individuals with Alzheimer’s Disease and Mild Cognitive Impairment: A Systematic Review. Neurosci. Inform. 2022, 2, 100104. [Google Scholar] [CrossRef]
- He, X.; Qin, W.; Liu, Y.; Zhang, X.; Duan, Y.; Song, J.; Li, K.; Jiang, T.; Yu, C. Abnormal salience network in normal aging and in amnestic mild cognitive impairment and Alzheimer’s disease. Hum. Brain Mapp. 2014, 35, 3446–3464. [Google Scholar] [CrossRef] [PubMed]
- Chand, G.B.; Thakuri, D.S.; Soni, B. Salience network anatomical and molecular markers are linked with cognitive dysfunction in mild cognitive impairment. J. Neuroimaging 2022, 32, 728–734. [Google Scholar] [CrossRef]
- Lim, H.K.; Nebes, R.; Snitz, B.; Cohen, A.; Mathis, C.; Price, J.; Weissfeld, L.; Klunk, W.; Aizenstein, H.J. Regional amyloid burden and intrinsic connectivity networks in cognitively normal elderly subjects. Brain 2014, 137, 3327–3338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, H.; Liang, K.; Wang, H.; Kong, S.; Hu, J.; Wu, F.; Sun, G. Functional degeneration in dorsal and ventral attention systems in amnestic mild cognitive impairment and Alzheimer’s disease: An fMRI study. Neurosci. Lett. 2015, 585, 160–165. [Google Scholar] [CrossRef] [PubMed]
- McDonough, I.M.; Wood, M.M.; Miller, W.S., Jr. Focus: Attention science: A review on the trajectory of attentional mechanisms in aging and the Alzheimer’s disease continuum through the attention network test. Yale J. Biol. Med. 2019, 92, 37. [Google Scholar]
- Mohammadian, F.; Zare Sadeghi, A.; Noroozian, M.; Malekian, V.; Abbasi Sisara, M.; Hashemi, H.; Mobarak Salari, H.; Valizadeh, G.; Samadi, F.; Sodaei, F. Quantitative Assessment of Resting-State Functional Connectivity MRI to Differentiate Amnestic Mild Cognitive Impairment, Late-Onset Alzheimer’s Disease From Normal Subjects. J. Magn. Reson. Imaging 2022. [Google Scholar] [CrossRef]
- Zhao, J.; Ding, X.; Du, Y.; Wang, X.; Men, G. Functional connectivity between white matter and gray matter based on fMRI for Alzheimer’s disease classification. Brain Behav. 2019, 9, e01407. [Google Scholar] [CrossRef]
- Gao, Z.; Feng, Y.; Ma, C.; Ma, K.; Cai, Q.; Initiative, A.s.D.N. Disrupted Time-Dependent and Functional Connectivity Brain Network in Alzheimer’s Disease: A Resting-State fMRI Study Based on Visibility Graph. Curr. Alzheimer Res. 2020, 17, 69–79. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef]
- Zhou, B.; Dou, X.; Wang, W.; Yao, H.; Feng, F.; Wang, P.; Yang, Z.; An, N.; Liu, B.; Zhang, X. Structural and functional connectivity abnormalities of the default mode network in patients with Alzheimer’s disease and mild cognitive impairment within two independent datasets. Methods 2022, 205, 29–38. [Google Scholar] [CrossRef]
- Zhao, C.; Huang, W.-J.; Feng, F.; Zhou, B.; Yao, H.-X.; Guo, Y.-E.; Wang, P.; Wang, L.-N.; Shu, N.; Zhang, X. Abnormal characterization of dynamic functional connectivity in Alzheimer’s disease. Neural Regen. Res. 2022, 17, 2014. [Google Scholar] [PubMed]
- Tang, F.; Zhu, D.; Ma, W.; Yao, Q.; Li, Q.; Shi, J. Differences changes in cerebellar functional connectivity between mild cognitive impairment and Alzheimer’s disease: A seed-based approach. Front. Neurol. 2021, 12, 987. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Guo, H.; Gao, Y.; Wang, X.; Cui, H.; Chen, Z.; Wang, B.; Xiang, J. Altered directed functional connectivity of the hippocampus in mild cognitive impairment and Alzheimer’s disease: A resting-state fMRI study. Front. Aging Neurosci. 2019, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Almgren, H.; Van de Steen, F.; Kühn, S.; Razi, A.; Friston, K.; Marinazzo, D. Variability and reliability of effective connectivity within the core default mode network: A multi-site longitudinal spectral DCM study. Neuroimage 2018, 183, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Scherr, M.; Utz, L.; Tahmasian, M.; Pasquini, L.; Grothe, M.J.; Rauschecker, J.P.; Grimmer, T.; Drzezga, A.; Sorg, C.; Riedl, V. Effective connectivity in the default mode network is distinctively disrupted in Alzheimer’s disease—A simultaneous resting-state FDG-PET/fMRI study. Hum. Brain Mapp. 2021, 42, 4134–4143. [Google Scholar] [CrossRef]
- Brettschneider, J.; Tredici, K.D.; Lee, V.M.-Y.; Trojanowski, J.Q. Spreading of pathology in neurodegenerative diseases: A focus on human studies. Nat. Rev. Neurosci. 2015, 16, 109–120. [Google Scholar] [CrossRef]
- Zhong, Y.; Huang, L.; Cai, S.; Zhang, Y.; von Deneen, K.M.; Ren, A.; Ren, J.; Initiative, A.s.D.N. Altered effective connectivity patterns of the default mode network in Alzheimer’s disease: An fMRI study. Neurosci. Lett. 2014, 578, 171–175. [Google Scholar] [CrossRef]
- Agosta, F.; Rocca, M.A.; Pagani, E.; Absinta, M.; Magnani, G.; Marcone, A.; Falautano, M.; Comi, G.; Gorno-Tempini, M.L.; Filippi, M. Sensorimotor network rewiring in mild cognitive impairment and Alzheimer’s disease. Hum. Brain Mapp. 2010, 31, 515–525. [Google Scholar] [CrossRef]
- Frässle, S.; Harrison, S.J.; Heinzle, J.; Clementz, B.A.; Tamminga, C.A.; Sweeney, J.A.; Gershon, E.S.; Keshavan, M.S.; Pearlson, G.D.; Powers, A. Regression dynamic causal modeling for resting-state fMRI. Hum. Brain Mapp. 2021, 42, 2159–2180. [Google Scholar] [CrossRef]
- Zarghami, T.S.; Friston, K.J. Dynamic effective connectivity. Neuroimage 2020, 207, 116453. [Google Scholar] [CrossRef]
- Ashburner, J.; Barnes, G.; Chen, C.-C.; Daunizeau, J.; Flandin, G.; Friston, K.; Kiebel, S.; Kilner, J.; Litvak, V.; Moran, R. SPM12 Manual; Wellcome Trust Centre for Neuroimaging: London, UK, 2014; Volume 2464, p. 4. [Google Scholar]
- Liu, Z.; Zhang, Y.; Bai, L.; Yan, H.; Dai, R.; Zhong, C.; Wang, H.; Wei, W.; Xue, T.; Feng, Y. Investigation of the effective connectivity of resting state networks in Alzheimer’s disease: A functional MRI study combining independent components analysis and multivariate Granger causality analysis. NMR Biomed. 2012, 25, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Chand, G.B.; Wu, J.; Hajjar, I.; Qiu, D. Interactions of the salience network and its subsystems with the default-mode and the central-executive networks in normal aging and mild cognitive impairment. Brain Connect. 2017, 7, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Greicius, M.D.; Gennatas, E.D.; Growdon, M.E.; Jang, J.Y.; Rabinovici, G.D.; Kramer, J.H.; Weiner, M.; Miller, B.L.; Seeley, W.W. Divergent network connectivity changes in behavioural variant frontotemporal dementia and Alzheimer’s disease. Brain 2010, 133, 1352–1367. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Spinelli, E.G.; Cividini, C.; Agosta, F. Resting state dynamic functional connectivity in neurodegenerative conditions: A review of magnetic resonance imaging findings. Front. Neurosci. 2019, 13, 657. [Google Scholar] [CrossRef]
- Badhwar, A.; Tam, A.; Dansereau, C.; Orban, P.; Hoffstaedter, F.; Bellec, P. Resting-state network dysfunction in Alzheimer’s disease: A systematic review and meta-analysis. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2017, 8, 73–85. [Google Scholar] [CrossRef]
- Li, G.; Hsu, L.-M.; Wu, Y.; Bozoki, A.C.; Shih, Y.-Y.I.; Yap, P.-T. Excitation-inhibition imbalance in Alzheimer′ s disease using multiscale neural model inversion of resting-state fMRI. medRxiv 2022. [Google Scholar] [CrossRef]
- Brier, M.R.; Thomas, J.B.; Ances, B.M. Network dysfunction in Alzheimer’s disease: Refining the disconnection hypothesis. Brain Connect. 2014, 4, 299–311. [Google Scholar] [CrossRef]
- Dennis, E.L.; Thompson, P.M. Functional brain connectivity using fMRI in aging and Alzheimer’s disease. Neuropsychol. Rev. 2014, 24, 49–62. [Google Scholar] [CrossRef]
- Luo, X.; Li, K.; Jia, Y.; Zeng, Q.; Jiaerken, Y.; Qiu, T.; Huang, P.; Xu, X.; Shen, Z.; Guan, X. Altered effective connectivity anchored in the posterior cingulate cortex and the medial prefrontal cortex in cognitively intact elderly APOE ε4 carriers: A preliminary study. Brain Imaging Behav. 2019, 13, 270–282. [Google Scholar] [CrossRef]
- Nie, Y.; Opoku, E.; Yasmin, L.; Song, Y.; Wang, J.; Wu, S.; Scarapicchia, V.; Gawryluk, J.; Wang, L.; Cao, J. Spectral Dynamic Causal Modelling of Resting-State fMRI: Relating Effective Brain Connectivity in the Default Mode Network to Genetics. arXiv 2019, arXiv:1901.09975. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Castanon, A. Handbook of Functional Connectivity Magnetic Resonance Imaging Methods in CONN; Hilbert-Press: Boston, MA, USA, 2020. [Google Scholar]
- Sharaev, M.G.; Zavyalova, V.V.; Ushakov, V.L.; Kartashov, S.I.; Velichkovsky, B.M. Effective connectivity within the default mode network: Dynamic causal modeling of resting-state fMRI data. Front. Hum. Neurosci. 2016, 10, 14. [Google Scholar] [CrossRef]
- Razi, A.; Seghier, M.L.; Zhou, Y.; McColgan, P.; Zeidman, P.; Park, H.-J.; Sporns, O.; Rees, G.; Friston, K.J. Large-scale DCMs for resting-state fMRI. Netw. Neurosci. 2017, 1, 222–241. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, X.; Ge, H.; Han, Y.; Pang, Z.; Liu, B.; Liu, S.; Friston, K. Heritability of the effective connectivity in the resting-state default mode network. Cereb. Cortex 2017, 27, 5626–5634. [Google Scholar] [CrossRef] [PubMed]
- Zeidman, P.; Jafarian, A.; Corbin, N.; Seghier, M.L.; Razi, A.; Price, C.J.; Friston, K.J. A guide to group effective connectivity analysis, part 1: First level analysis with DCM for fMRI. Neuroimage 2019, 200, 174–190. [Google Scholar] [CrossRef] [PubMed]
- Zeidman, P.; Jafarian, A.; Seghier, M.L.; Litvak, V.; Cagnan, H.; Price, C.J.; Friston, K.J. A guide to group effective connectivity analysis, part 2: Second level analysis with PEB. Neuroimage 2019, 200, 12–25. [Google Scholar] [CrossRef]
- Di, X.; Biswal, B.B. Identifying the default mode network structure using dynamic causal modeling on resting-state functional magnetic resonance imaging. Neuroimage 2014, 86, 53–59. [Google Scholar] [CrossRef]
- Yu, C.; Li, J.; Liu, Y.; Qin, W.; Li, Y.; Shu, N.; Jiang, T.; Li, K. White matter tract integrity and intelligence in patients with mental retardation and healthy adults. Neuroimage 2008, 40, 1533–1541. [Google Scholar] [CrossRef]
- Hedden, T.; Van Dijk, K.R.; Becker, J.A.; Mehta, A.; Sperling, R.A.; Johnson, K.A.; Buckner, R.L. Disruption of functional connectivity in clinically normal older adults harboring amyloid burden. J. Neurosci. 2009, 29, 12686–12694. [Google Scholar] [CrossRef]
- Hagmann, P.; Cammoun, L.; Gigandet, X.; Meuli, R.; Honey, C.J.; Wedeen, V.J.; Sporns, O. Mapping the structural core of human cerebral cortex. PLoS Biol. 2008, 6, e159. [Google Scholar] [CrossRef]
- Forouzannezhad, P.; Abbaspour, A.; Fang, C.; Cabrerizo, M.; Loewenstein, D.; Duara, R.; Adjouadi, M. A survey on applications and analysis methods of functional magnetic resonance imaging for Alzheimer’s disease. J. Neurosci. Methods 2019, 317, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Jahn, H. Memory loss in Alzheimer’s disease. Dialogues Clin. Neurosci. 2022, 15, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Liu, X.; Song, H.; Li, K.; Wang, Z. Altered functional connectivity of cognitive-related cerebellar subregions in Alzheimer’s disease. Front. Aging Neurosci. 2017, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yan, C.; Zhao, C.; Qi, Z.; Zhou, W.; Lu, J.; He, Y.; Li, K. Spatial patterns of intrinsic brain activity in mild cognitive impairment and Alzheimer’s disease: A resting-state functional MRI study. Hum. Brain Mapp. 2011, 32, 1720–1740. [Google Scholar] [CrossRef]
- Dickerson, B.C.; Sperling, R.A. Large-scale functional brain network abnormalities in Alzheimer’s disease: Insights from functional neuroimaging. Behav. Neurol. 2009, 21, 63–75. [Google Scholar] [CrossRef]
- Hampstead, B.M.; Khoshnoodi, M.; Yan, W.; Deshpande, G.; Sathian, K. Patterns of effective connectivity during memory encoding and retrieval differ between patients with mild cognitive impairment and healthy older adults. Neuroimage 2016, 124, 997–1008. [Google Scholar] [CrossRef]
- Li, R.; Wu, X.; Fleisher, A.S.; Reiman, E.M.; Chen, K.; Yao, L. Attention-related networks in Alzheimer’s disease: A resting functional MRI study. Hum. Brain Mapp. 2012, 33, 1076–1088. [Google Scholar] [CrossRef]
- Vidoni, E.D.; Thomas, G.P.; Honea, R.A.; Loskutova, N.; Burns, J.M. Evidence of altered corticomotor system connectivity in early-stage Alzheimer’s disease. J. Neurol. Phys. Ther. 2012, 36, 8. [Google Scholar] [CrossRef]
- Brier, M.R.; Thomas, J.B.; Snyder, A.Z.; Benzinger, T.L.; Zhang, D.; Raichle, M.E.; Holtzman, D.M.; Morris, J.C.; Ances, B.M. Loss of intranetwork and internetwork resting state functional connections with Alzheimer’s disease progression. J. Neurosci. 2012, 32, 8890–8899. [Google Scholar] [CrossRef]
- Rytsar, R.; Fornari, E.; Frackowiak, R.S.; Ghika, J.A.; Knyazeva, M.G. Inhibition in early Alzheimer’s disease: An fMRI-based study of effective connectivity. Neuroimage 2011, 57, 1131–1139. [Google Scholar] [CrossRef]
- Montembeault, M.; Chapleau, M.; Jarret, J.; Boukadi, M.; Laforce, R., Jr.; Wilson, M.A.; Rouleau, I.; Brambati, S.M. Differential language network functional connectivity alterations in Alzheimer’s disease and the semantic variant of primary progressive aphasia. Cortex 2019, 117, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Neufang, S.; Akhrif, A.; Riedl, V.; Förstl, H.; Kurz, A.; Zimmer, C.; Sorg, C.; Wohlschläger, A.M. Disconnection of frontal and parietal areas contributes to impaired attention in very early Alzheimer’s disease. J. Alzheimer’s Dis. 2011, 25, 309–321. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Late-Onset AD | aMCI | Normal | p Value | Post Hoc Pairwise Comparisons | Observed Power | |
|---|---|---|---|---|---|---|
| (n = 13) | (n = 16) | (n = 14) | ||||
| Age (years) | 77.77 ± 7.95 | 72.44 ± 7.11 | 71.57 ± 7.14 | 0.062 | - | 0.696 |
| Gender (female–male) | 5–8 | 6–10 | 6–8 | 0.95 | - | 0.88 |
| FAST | 4–5 | 3 | 1 | <0.001 | Normal vs. aMCI: < 0.001 | 0.73 |
| Normal vs. AD: < 0.001 | ||||||
| aMCI vs. AD: < 0.001 | ||||||
| MMSE | 18.62 ± 1.39 | 25.06 ± 2.24 | 28.07 ± 0.83 | <0.001 | Normal vs. aMCI: 0.031 | 0.99 |
| Normal vs. AD: < 0.008 | ||||||
| aMCI vs. AD: < 0.001 | ||||||
| MoCA | 16.15 ± 3.53 | 22.19 ± 3.94 | 27.29 ± 1.14 | <0.001 | Normal vs. aMCI: 0.003 | 0.98 |
| Normal vs. AD: < 0.001 | ||||||
| aMCI vs. AD: < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammadian, F.; Noroozian, M.; Sadeghi, A.Z.; Malekian, V.; Saffar, A.; Talebi, M.; Hashemi, H.; Mobarak Salari, H.; Samadi, F.; Sodaei, F.; et al. Effective Connectivity Evaluation of Resting-State Brain Networks in Alzheimer’s Disease, Amnestic Mild Cognitive Impairment, and Normal Aging: An Exploratory Study. Brain Sci. 2023, 13, 265. https://doi.org/10.3390/brainsci13020265
Mohammadian F, Noroozian M, Sadeghi AZ, Malekian V, Saffar A, Talebi M, Hashemi H, Mobarak Salari H, Samadi F, Sodaei F, et al. Effective Connectivity Evaluation of Resting-State Brain Networks in Alzheimer’s Disease, Amnestic Mild Cognitive Impairment, and Normal Aging: An Exploratory Study. Brain Sciences. 2023; 13(2):265. https://doi.org/10.3390/brainsci13020265
Chicago/Turabian StyleMohammadian, Fatemeh, Maryam Noroozian, Arash Zare Sadeghi, Vahid Malekian, Azam Saffar, Mahsa Talebi, Hasan Hashemi, Hanieh Mobarak Salari, Fardin Samadi, Forough Sodaei, and et al. 2023. "Effective Connectivity Evaluation of Resting-State Brain Networks in Alzheimer’s Disease, Amnestic Mild Cognitive Impairment, and Normal Aging: An Exploratory Study" Brain Sciences 13, no. 2: 265. https://doi.org/10.3390/brainsci13020265
APA StyleMohammadian, F., Noroozian, M., Sadeghi, A. Z., Malekian, V., Saffar, A., Talebi, M., Hashemi, H., Mobarak Salari, H., Samadi, F., Sodaei, F., & Rad, H. S. (2023). Effective Connectivity Evaluation of Resting-State Brain Networks in Alzheimer’s Disease, Amnestic Mild Cognitive Impairment, and Normal Aging: An Exploratory Study. Brain Sciences, 13(2), 265. https://doi.org/10.3390/brainsci13020265