




Efficiency of the Brain Network Is Associated with the Mental Workload with Developed Mental Schema


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
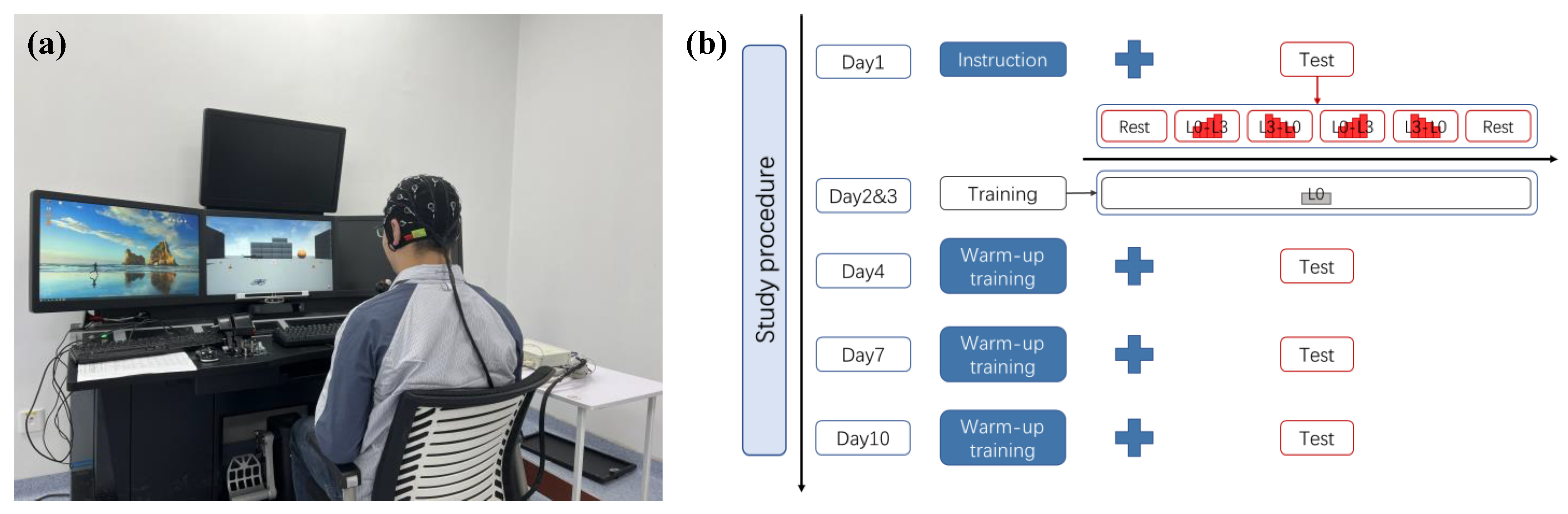
2.2. Experimental Protocol
2.3. EEG Acquisition and Preprocessing
2.4. Functional Connectivity and Network Topology Analysis
2.5. Statistical Analysis
3. Results
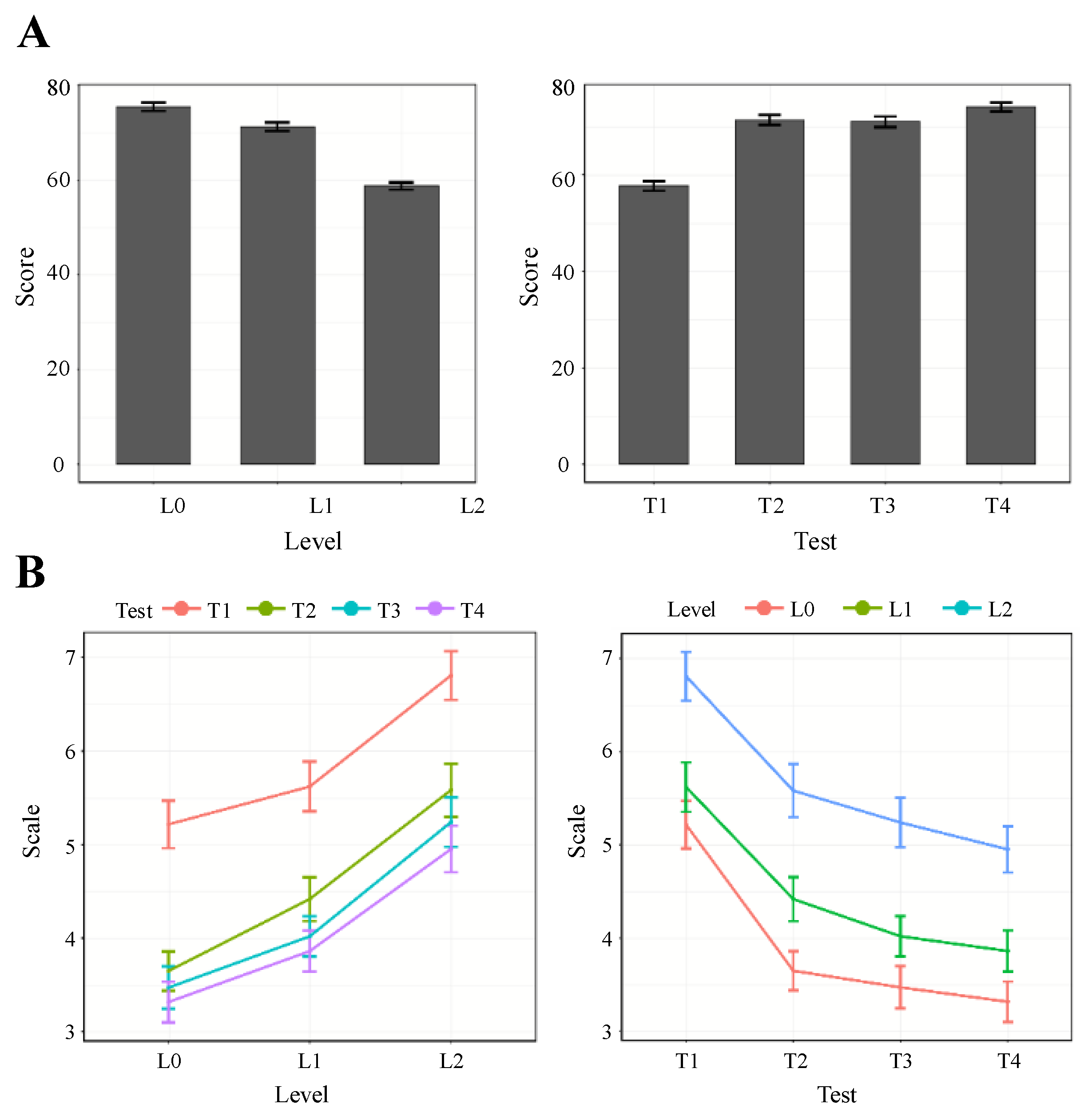
3.1. Task Performance
3.2. Perceived Workload
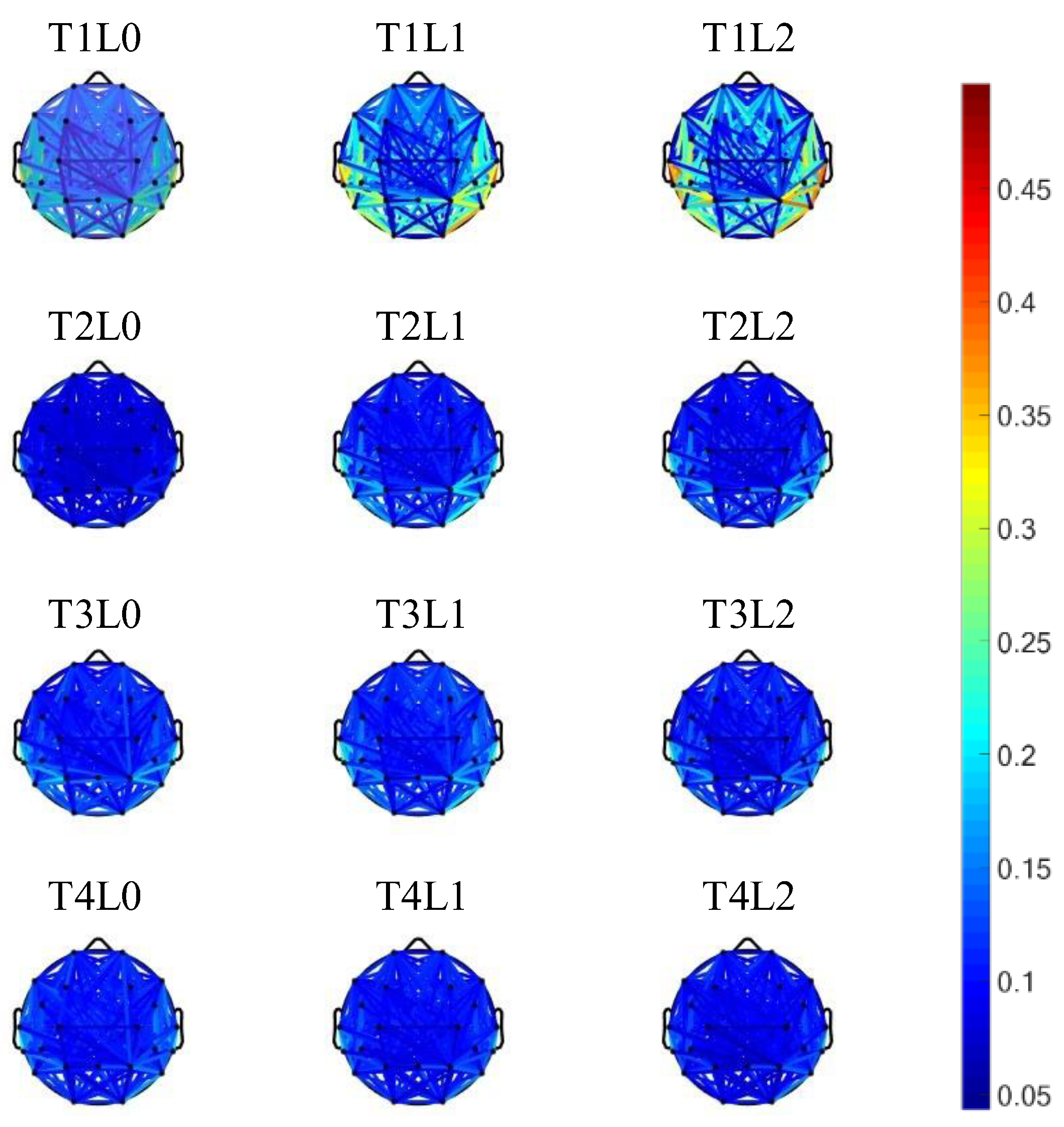
3.3. Network Analysis Results
4. Discussion
Limitations and Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Donnell, R.D.; Eggemeier, F.T. Workload assessment methodology. In Handbook of Perception and Human Performance, Volume 2: Cognitive Processes and Performance; John Wiley & Sons: Oxford, UK, 1986; pp. 1–49. [Google Scholar]
- Rojas, R.F.; Debie, E.; Fidock, J.; Barlow, M.; Kasmarik, K.; Anavatti, S.; Garratt, M.; Abbass, H. Electroencephalographic Workload Indicators During Teleoperation of an Unmanned Aerial Vehicle Shepherding a Swarm of Unmanned Ground Vehicles in Contested Environments. Front. Neurosci. 2020, 14, 40. [Google Scholar] [CrossRef]
- Tao, D.; Tan, H.; Wang, H.; Zhang, X.; Qu, X.; Zhang, T. A Systematic Review of Physiological Measures of Mental Workload. Int. J. Environ. Res. Public Health 2019, 16, 2716. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.G.; Staveland, L.E. Development of NASA-TLX (Task Load Index): Results of Empirical and Theoretical Research. In Advances in Psychology; Hancock, P.A., Meshkati, N., Eds.; North-Holland: Amsterdam, The Netherlands, 1988; Volume 52, pp. 139–183. [Google Scholar]
- Boutyline, A.; Soter, L.K. Cultural Schemas: What They Are, How to Find Them, and What to Do Once You’ve Caught One. Am. Sociol. Rev. 2021, 86, 728–758. [Google Scholar] [CrossRef]
- Iqbal, M.U.; Shahab, M.A.; Choudhary, M.; Srinivasan, B.; Srinivasan, R. Electroencephalography (EEG) based cognitive measures for evaluating the effectiveness of operator training. Process Saf. Environ. Prot. 2021, 150, 51–67. [Google Scholar] [CrossRef]
- Bernacer, J.; Murillo, J.I. The Aristotelian conception of habit and its contribution to human neuroscience. Front. Hum. Neurosci. 2014, 8, 883. [Google Scholar] [CrossRef] [Green Version]
- Taya, F.; Sun, Y.; Babiloni, F.; Thakor, N.V.; Bezerianos, A. Topological Changes in the Brain Network Induced by the Training on a Piloting Task: An EEG-Based Functional Connectome Approach. IEEE Trans. Neural Syst. Rehabil. Eng. Publ. IEEE Eng. Med. Biol. Soc. 2018, 26, 263–271. [Google Scholar] [CrossRef]
- Ghaderi, A.H.; Nazari, M.A.; Darooneh, A.H. Functional brain segregation changes during demanding mathematical task. Int. J. Neurosci. 2019, 129, 904–915. [Google Scholar] [CrossRef]
- Nguyen, P.T.M.; Li, X.Z.; Hayashi, Y.; Yano, S.; Kondo, T. Estimation of brain dynamics under visuomotor task using functional connectivity analysis based on graph theory. In Proceedings of the 19th Annual IEEE International Conference on Bioinformatics and Bioengineering (BIBE), Athens, Greece, 28–30 October 2019; pp. 577–582. [Google Scholar]
- Cattai, T.; Colonnese, S.; Corsi, M.C.; Bassett, D.S.; Scarano, G.; Fallani, F.D. Characterization of Mental States through Node Connectivity between Brain Signals. In Proceedings of the European Signal Processing Conference (EUSIPCO), Rome, Italy, 3–7 September 2018; pp. 1377–1381. [Google Scholar]
- Sporns, O. Contributions and challenges for network models in cognitive neuroscience. Nat. Neurosci. 2014, 17, 652–660. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.M.; Koenig, T. EEG microstates as a tool for studying the temporal dynamics of whole-brain neuronal networks: A review. NeuroImage 2018, 180, 577–593. [Google Scholar] [CrossRef]
- Goldenberg, D.; Galván, A. The use of functional and effective connectivity techniques to understand the developing brain. Dev. Cogn. Neurosci. 2015, 12, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Kitzbichler, M.G.; Henson, R.N.; Smith, M.L.; Nathan, P.J.; Bullmore, E.T. Cognitive effort drives workspace configuration of human brain functional networks. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 8259–8270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Ren, A.; Shang, J.; Lei, Q.; Zhang, Y.; Yin, Z.; Li, J.; von Deneen, K.M.; Huang, L. Combining Partial Directed Coherence and Graph Theory to Analyse Effective Brain Networks of Different Mental Tasks. Front. Hum. Neurosci. 2016, 10, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klados, M.A.; Kanatsouli, K.; Antoniou, I.; Babiloni, F.; Tsirka, V.; Bamidis, P.D.; Micheloyannis, S. A Graph theoretical approach to study the organization of the cortical networks during different mathematical tasks. PLoS ONE 2013, 8, e71800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakkos, I.; Dimitrakopoulos, G.N.; Gao, L.; Zhang, Y.; Qi, P.; Matsopoulos, G.K.; Thakor, N.; Bezerianos, A.; Sun, Y. Mental Workload Drives Different Reorganizations of Functional Cortical Connectivity Between 2D and 3D Simulated Flight Experiments. IEEE Trans. Neural Syst. Rehabil. Eng. Publ. IEEE Eng. Med. Biol. Soc. 2019, 27, 1704–1713. [Google Scholar] [CrossRef]
- Dimitrakopoulos, G.N.; Kakkos, I.; Dai, Z.X.; Wang, H.T.; Sgarbas, K.; Thakor, N.; Bezerianos, A.; Sun, Y. Functional Connectivity Analysis of Mental Fatigue Reveals Different Network Topological Alterations Between Driving and Vigilance Tasks. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 740–749. [Google Scholar] [CrossRef]
- Qi, P.; Gao, L.Y.; Meng, J.J.; Thakor, N.; Bezerianos, A.; Sun, Y. Effects of Rest-Break on Mental Fatigue Recovery Determined by a Novel Temporal Brain Network Analysis of Dynamic Functional Connectivity. IEEE Trans. Neural Syst. Rehabil. Eng. 2020, 28, 62–71. [Google Scholar] [CrossRef]
- Van Atteveldt, N.; Murray, M.M.; Thut, G.; Schroeder, C.E. Multisensory integration: Flexible use of general operations. Neuron 2014, 81, 1240–1253. [Google Scholar] [CrossRef] [Green Version]
- VanRullen, R. Perceptual Cycles. Trends Cogn. Sci. 2016, 20, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitriadis, S.I.; Sun, Y.; Kwok, K.; Laskaris, N.A.; Bezerianos, A. A tensorial approach to access cognitive workload related to mental arithmetic from EEG functional connectivity estimates. IEEE Eng. Med. Biol. Soc. Annu. Int. Conf. 2013, 2013, 2940–2943. [Google Scholar] [CrossRef]
- Shaw, E.P.; Rietschel, J.C.; Shuggi, I.M.; Xu, Y.; Chen, S.; Miller, M.W.; Hatfield, B.D.; Gentili, R.J. Cerebral cortical networking for mental workload assessment under various demands during dual-task walking. Exp. Brain Res. 2019, 237, 2279–2295. [Google Scholar] [CrossRef]
- Ren, S.; Taya, F.; Sun, Y.; deSouza, J.; Thakor, N.V.; Bezerianos, A. Assessing Small-worldness of Dynamic Functional Brain Connectivity During Complex Tasks. In Proceedings of the 37th Annual International Conference of the IEEE-Engineering-in-Medicine-and-Biology-Society (EMBC), Milan, Italy, 25–29 August 2015; pp. 2904–2907. [Google Scholar]
- Ren, S.; Li, J.H.; Taya, F.; Desouza, J.; Thakor, N.V.; Bezerianos, A. Dynamic Functional Segregation and Integration in Human Brain Network During Complex Tasks. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 547–556. [Google Scholar] [CrossRef]
- Porter, S.; Silverberg, N.D.; Virji-Babul, N. Cortical activity and network organization underlying physical and cognitive exertion in active young adult athletes: Implications for concussion. J. Sci. Med. Sport 2019, 22, 397–402. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; van Wingerden, M.; Battaglia, F.; Pennartz, C.M. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. NeuroImage 2011, 55, 1548–1565. [Google Scholar] [CrossRef]
- Gu, H.; Yao, Q.; Chen, H.; Ding, Z.; Zhao, X.; Liu, H.; Feng, Y.; Li, C.; Li, X. The effect of mental schema evolution on mental workload measurement: An EEG study with simulated quadrotor UAV operation. J. Neural Eng. 2022, 19, 026058. [Google Scholar] [CrossRef]
- Cooper, G.; Harper, R. The Use of Pilot Ratings in Evaluation of Aircraft Handling Qualities; NASA Ames Technical Report; National Aeronautics and Space Administration: Washington, DC, USA, 1969. Available online: https://www.researchgate.net/publication/24332309_The_use_of_pilot_ratings_in_evaluation_of_aircraft_ha (accessed on 31 January 2023).
- Iriarte, J.; Urrestarazu, E.; Valencia, M.; Alegre, M.; Malanda, A.; Viteri, C.; Artieda, J. Independent component analysis as a tool to eliminate artifacts in EEG: A quantitative study. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 2003, 20, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimesch, W.; Schimke, H.; Pfurtscheller, G. Alpha frequency, cognitive load and memory performance. Brain Topogr. 1993, 5, 241–251. [Google Scholar] [CrossRef]
- Borghini, G.; Astolfi, L.; Vecchiato, G.; Mattia, D.; Babiloni, F. Measuring neurophysiological signals in aircraft pilots and car drivers for the assessment of mental workload, fatigue and drowsiness. Neurosci. Biobehav. Rev. 2014, 44, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Sauseng, P.; Griesmayr, B.; Freunberger, R.; Klimesch, W. Control mechanisms in working memory: A possible function of EEG theta oscillations. Neurosci. Biobehav. Rev. 2010, 34, 1015–1022. [Google Scholar] [CrossRef]
- Krugel, L.K.; Biele, G.; Mohr, P.N.; Li, S.C.; Heekeren, H.R. Genetic variation in dopaminergic neuromodulation influences the ability to rapidly and flexibly adapt decisions. Proc. Natl. Acad. Sci. USA 2009, 106, 17951–17956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, S.; Bullmore, E. Efficiency and cost of economical brain functional networks. PLoS Comput. Biol. 2007, 3, e17. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.P.; Stam, C.J.; Boersma, M.; Pol, H.E.H. Small-world and scale-free organization of voxel-based resting-state functional connectivity in the human brain. NeuroImage 2008, 43, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Palva, S.; Palva, J.M. Roles of Brain Criticality and Multiscale Oscillations in Temporal Predictions for Sensorimotor Processing. Trends Neurosci. 2018, 41, 729–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, A.C.; Ambrosia, V.G.; Hinkley, E.A. Unmanned Aircraft Systems in Remote Sensing and Scientific Research: Classification and Considerations of Use. Remote Sens. 2012, 4, 1671–1692. [Google Scholar] [CrossRef] [Green Version]
- Braun, U.; Schäfer, A.; Walter, H.; Erk, S.; Romanczuk-Seiferth, N.; Haddad, L.; Schweiger, J.I.; Grimm, O.; Heinz, A.; Tost, H.; et al. Dynamic reconfiguration of frontal brain networks during executive cognition in humans. Proc. Natl. Acad. Sci. USA 2015, 112, 11678–11683. [Google Scholar] [CrossRef] [Green Version]
- Fornito, A.; Harrison, B.J.; Zalesky, A.; Simons, J.S. Competitive and cooperative dynamics of large-scale brain functional networks supporting recollection. Proc. Natl. Acad. Sci. USA 2012, 109, 12788–12793. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zou, Q.; He, Y.; Yang, Y. Topologically Reorganized Connectivity Architecture of Default-Mode, Executive-Control, and Salience Networks across Working Memory Task Loads. Cereb. Cortex 2016, 26, 1501–1511. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.W.; Reynolds, J.R.; Power, J.D.; Repovs, G.; Anticevic, A.; Braver, T.S. Multi-task connectivity reveals flexible hubs for adaptive task control. Nat. Neurosci. 2013, 16, 1348–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Vasilakos, A.V.; He, Y. Small-world human brain networks: Perspectives and challenges. Neurosci. Biobehav. Rev. 2017, 77, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Mišić, B.; Betzel, R.F.; Nematzadeh, A.; Goñi, J.; Griffa, A.; Hagmann, P.; Flammini, A.; Ahn, Y.Y.; Sporns, O. Cooperative and Competitive Spreading Dynamics on the Human Connectome. Neuron 2015, 86, 1518–1529. [Google Scholar] [CrossRef] [Green Version]
- Hansen, E.C.; Battaglia, D.; Spiegler, A.; Deco, G.; Jirsa, V.K. Functional connectivity dynamics: Modeling the switching behavior of the resting state. NeuroImage 2015, 105, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Resting brains never rest: Computational insights into potential cognitive architectures. Trends Neurosci. 2013, 36, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haimovici, A.; Tagliazucchi, E.; Balenzuela, P.; Chialvo, D.R. Brain organization into resting state networks emerges at criticality on a model of the human connectome. Phys. Rev. Lett. 2013, 110, 178101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbara, A.; Khalil, M.; O’Neill, G.; Dujardin, K.; El Traboulsi, Y.; Wendling, F.; Hassan, M. Detecting modular brain states in rest and task. Netw. Neurosci. 2019, 3, 878–901. [Google Scholar] [CrossRef]
- Nackaerts, E.; D’Cruz, N.; Dijkstra, B.W.; Gilat, M.; Kramer, T.; Nieuwboer, A. Towards understanding neural network signatures of motor skill learning in Parkinson’s disease and healthy aging. Br. J. Radiol. 2019, 92, 20190071. [Google Scholar] [CrossRef]
- Fallani, F.D.; Baluch, F.; Astolfi, L.; Subramanian, D.; Zouridakis, G.; Babiloni, F. Structural organization of functional networks from EEG signals during motor learning tasks. Int. J. Bifurc. Chaos 2010, 20, 905–912. [Google Scholar] [CrossRef]
- Ismail, L.E.; Karwowski, W. A Graph Theory-Based Modeling of Functional Brain Connectivity Based on EEG: A Systematic Review in the Context of Neuroergonomics. IEEE Access 2020, 8, 155103–155135. [Google Scholar] [CrossRef]
- McDonnell, J.; Murray, N.P.; Ahn, S.; Clemens, S.; Everhart, E.; Mizelle, J.C. Examination and Comparison of Theta Band Connectivity in Left- and Right-Hand Dominant Individuals throughout a Motor Skill Acquisition. Symmetry 2021, 13, 728. [Google Scholar] [CrossRef]
- Gu, H.; Chen, H.; Yao, Q.; Wang, S.; Ding, Z.; Yuan, Z.; Zhao, X.; Li, X. Cortical theta-gamma coupling tracks the mental workload as an indicator of mental schema development during simulated quadrotor UAV operation. J. Neural Eng. 2022, 19, 066029. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, H.; Chen, H.; Yao, Q.; He, W.; Wang, S.; Yang, C.; Li, J.; Liu, H.; Li, X.; Zhao, X.; et al. Efficiency of the Brain Network Is Associated with the Mental Workload with Developed Mental Schema. Brain Sci. 2023, 13, 373. https://doi.org/10.3390/brainsci13030373
Gu H, Chen H, Yao Q, He W, Wang S, Yang C, Li J, Liu H, Li X, Zhao X, et al. Efficiency of the Brain Network Is Associated with the Mental Workload with Developed Mental Schema. Brain Sciences. 2023; 13(3):373. https://doi.org/10.3390/brainsci13030373
Chicago/Turabian StyleGu, Heng, He Chen, Qunli Yao, Wenbo He, Shaodi Wang, Chao Yang, Jiaxi Li, Huapeng Liu, Xiaoli Li, Xiaochuan Zhao, and et al. 2023. "Efficiency of the Brain Network Is Associated with the Mental Workload with Developed Mental Schema" Brain Sciences 13, no. 3: 373. https://doi.org/10.3390/brainsci13030373
APA StyleGu, H., Chen, H., Yao, Q., He, W., Wang, S., Yang, C., Li, J., Liu, H., Li, X., Zhao, X., & Liang, G. (2023). Efficiency of the Brain Network Is Associated with the Mental Workload with Developed Mental Schema. Brain Sciences, 13(3), 373. https://doi.org/10.3390/brainsci13030373