


Exploring a Possible Interplay between Schizophrenia, Oxytocin, and Estrogens: A Narrative Review

Abstract
:1. Introduction
2. Methods
3. The Oxytocin System in Schizophrenia
3.1. The Endogenous Oxytocin System in Schizophrenia
Positive and Negative Symptoms
3.2. Exogenous Oxytocin Effects in Schizophrenia
3.2.1. Clinical Studies
3.2.2. Preclinical Studies
4. Involvement of Sex Hormones
4.1. Estrogen Hypothesis
Antipsychotics and Estrogens
5. Schizophrenia, Oxytocin, Estrogens: A Possible Interplay
6. Limitations and Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; ISBN 0-89042-554-X. [Google Scholar]
- Zhang, K.; Liao, P.; Wen, J.; Hu, Z. Synaptic Plasticity in Schizophrenia Pathophysiology. IBRO Neurosci. Rep. 2022, 13, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, S.; Chafee, M.V.; Lee, E.; Morishita, H. Psychosis Spectrum Illnesses as Disorders of Prefrontal Critical Period Plasticity. Neuropsychopharmacology 2023, 48, 168–185. [Google Scholar] [CrossRef] [PubMed]
- Keepers, G.A.; Fochtmann, L.J.; Anzia, J.M.; Benjamin, S.; Lyness, J.M.; Mojtabai, R.; Servis, M.; Walaszek, A.; Buckley, P.; Lenzenweger, M.F.; et al. The American Psychiatric Association Practice Guideline for the Treatment of Patients with Schizophrenia. AJP 2020, 177, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Bujanow, W. Hormones in the Treatment of Psychoses. Br. Med. J. 1972, 4, 298. [Google Scholar] [CrossRef] [Green Version]
- Argiolas, A.; Gessa, G.L. Central Functions of Oxytocin. Neurosci. Biobehav. Rev. 1991, 15, 217–231. [Google Scholar] [CrossRef]
- Iovino, M.; Giagulli, V.A.; Licchelli, B.; Iovino, E.; Guastamacchia, E.; Triggiani, V. Synaptic Inputs of Neural Afferent Pathways to Vasopressin- and Oxytocin-Secreting Neurons of Supraoptic and Paraventricular Hypothalamic Nuclei. Endocr. Metab. Immune Disord. Drug Targets 2016, 16, 276–287. [Google Scholar] [CrossRef]
- Gimpl, G.; Fahrenholz, F. The Oxytocin Receptor System: Structure, Function, and Regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [Green Version]
- Knobloch, H.; Grinevich, V. Evolution of Oxytocin Pathways in the Brain of Vertebrates. Front. Behav. Neurosci. 2014, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Fonagy, P.; Koos, O.; Dorsett, K.; Strathearn, L. Maternal Oxytocin Response Predicts Mother-to-Infant Gaze. Brain Res. 2014, 1580, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Carter, C.S. Sex, Love and Oxytocin: Two Metaphors and a Molecule. Neurosci. Biobehav. Rev. 2022, 143, 104948. [Google Scholar] [CrossRef]
- Barchi-Ferreira, A.M.; Osório, F.L. Associations between Oxytocin and Empathy in Humans: A Systematic Literature Review. Psychoneuroendocrinology 2021, 129, 105268. [Google Scholar] [CrossRef]
- De Dreu, C.K.W.; Kret, M.E. Oxytocin Conditions Intergroup Relations Through Upregulated In-Group Empathy, Cooperation, Conformity, and Defense. Biol. Psychiatry 2016, 79, 165–173. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Young, L.J.; Insel, T.R. The Neuroendocrine Basis of Social Recognition. Front. Neuroendocr. 2002, 23, 200–224. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.; Shergill, S.S.; Averbeck, B.B. Oxytocin Decreases Aversion to Angry Faces in an Associative Learning Task. Neuropsychopharmacology 2010, 35, 2502–2509. [Google Scholar] [CrossRef]
- Jurek, B.; Meyer, M. Anxiolytic and Anxiogenic? How the Transcription Factor MEF2 Might Explain the Manifold Behavioral Effects of Oxytocin. Front. Endocrinol. 2020, 11, 186. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.K.; Chen, C.-H.; Lane, H.-Y. Oxytocin in Schizophrenia: Pathophysiology and Implications for Future Treatment. Int. J. Mol. Sci. 2021, 22, 2146. [Google Scholar] [CrossRef]
- Davis, M.C.; Lee, J.; Horan, W.P.; Clarke, A.D.; McGee, M.R.; Green, M.F.; Marder, S.R. Effects of Single Dose Intranasal Oxytocin on Social Cognition in Schizophrenia. Schizophr. Res. 2013, 147, 393–397. [Google Scholar] [CrossRef]
- Feifel, D.; Macdonald, K.; Nguyen, A.; Cobb, P.; Warlan, H.; Galangue, B.; Minassian, A.; Becker, O.; Cooper, J.; Perry, W.; et al. Adjunctive Intranasal Oxytocin Reduces Symptoms in Schizophrenia Patients. Biol. Psychiatry 2010, 68, 678–680. [Google Scholar] [CrossRef]
- Riecher-Rössler, A.; Häfner, H. Gender Aspects in Schizophrenia: Bridging the Border between Social and Biological Psychiatry. Acta Psychiatr. Scand. 2000, 102, 58–62. [Google Scholar] [CrossRef]
- Lu, Q.; Hu, S. Sex Differences of Oxytocin and Vasopressin in Social Behaviors. Handb. Clin. Neurol. 2021, 180, 65–88. [Google Scholar] [CrossRef]
- Taylor, S.E.; Klein, L.C.; Lewis, B.P.; Gruenewald, T.L.; Gurung, R.A.; Updegraff, J.A. Biobehavioral Responses to Stress in Females: Tend-and-Befriend, Not Fight-or-Flight. Psychol. Rev. 2000, 107, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Preckel, K.; Scheele, D.; Kendrick, K.M.; Maier, W.; Hurlemann, R. Oxytocin Facilitates Social Approach Behavior in Women. Front. Behav. Neurosci. 2014, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheele, D.; Striepens, N.; Kendrick, K.M.; Schwering, C.; Noelle, J.; Wille, A.; Schläpfer, T.E.; Maier, W.; Hurlemann, R. Opposing Effects of Oxytocin on Moral Judgment in Males and Females. Hum. Brain Mapp. 2014, 35, 6067–6076. [Google Scholar] [CrossRef]
- Hoge, E.A.; Anderson, E.; Lawson, E.A.; Bui, E.; Fischer, L.E.; Khadge, S.D.; Barrett, L.F.; Simon, N.M. Gender Moderates the Effect of Oxytocin on Social Judgments. Hum. Psychopharmacol. 2014, 29, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Shofty, M.; Levkovitz, Y.; Shamay-Tsoory, S.G. Oxytocin Facilitates Accurate Perception of Competition in Men and Kinship in Women. Soc. Cogn. Affect Neurosci. 2013, 8, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Scheele, D.; Striepens, N.; Güntürkün, O.; Deutschländer, S.; Maier, W.; Kendrick, K.M.; Hurlemann, R. Oxytocin Modulates Social Distance between Males and Females. J. Neurosci. 2012, 32, 16074–16079. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, K. Sex, Receptors, and Attachment: A Review of Individual Factors Influencing Response to Oxytocin. Front. Neurosci. 2013, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Gogos, A.; Ney, L.J.; Seymour, N.; Van Rheenen, T.E.; Felmingham, K.L. Sex Differences in Schizophrenia, Bipolar Disorder, and Post-Traumatic Stress Disorder: Are Gonadal Hormones the Link? Br. J. Pharm. 2019, 176, 4119–4135. [Google Scholar] [CrossRef]
- Engel, S.; Klusmann, H.; Ditzen, B.; Knaevelsrud, C.; Schumacher, S. Menstrual Cycle-Related Fluctuations in Oxytocin Concentrations: A Systematic Review and Meta-Analysis. Front. Neuroendocrinol. 2019, 52, 144–155. [Google Scholar] [CrossRef]
- Seeman, M.V. The Role of Estrogen in Schizophrenia. J. Psychiatry Neurosci. 1996, 21, 123–127. [Google Scholar]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—A Scale for the Quality Assessment of Narrative Review Articles. Res. Integr. Peer Rev. 2019, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, H.K.; Stephens, S.L.; Young, W.S. Oxytocin as a Natural Antipsychotic: A Study Using Oxytocin Knockout Mice. Mol. Psychiatry 2009, 14, 190–196. [Google Scholar] [CrossRef]
- Ventriglio, A.; Bellomo, A.; Ricci, F.; Magnifico, G.; Rinaldi, A.; Borraccino, L.; Piccininni, C.; Cuoco, F.; Gianfelice, G.; Fornaro, M.; et al. New Pharmacological Targets for the Treatment of Schizophrenia: A Literature Review. Curr. Top. Med. Chem. 2021, 21, 1500–1516. [Google Scholar] [CrossRef]
- Walss-Bass, C.; Fernandes, J.M.; Roberts, D.L.; Service, H.; Velligan, D. Differential Correlations between Plasma Oxytocin and Social Cognitive Capacity and Bias in Schizophrenia. Schizophr. Res. 2013, 147, 387–392. [Google Scholar] [CrossRef]
- Jobst, A.; Dehning, S.; Ruf, S.; Notz, T.; Buchheim, A.; Henning-Fast, K.; Meißner, D.; Meyer, S.; Bondy, B.; Müller, N.; et al. Oxytocin and Vasopressin Levels Are Decreased in the Plasma of Male Schizophrenia Patients. Acta Neuropsychiatr. 2014, 26, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tao, H.; Yang, X.; Huang, K.; Zhang, X.; Li, C. Decreased Serum Oxytocin and Increased Homocysteine in First-Episode Schizophrenia Patients. Front. Psychiatry 2019, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.C.; de Osório, F.L. Peripheral Oxytocin Concentrations in Psychiatric Disorders—A Systematic Review and Methanalysis: Further Evidence. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 117, 110561. [Google Scholar] [CrossRef]
- Hernández-Díaz, Y.; González-Castro, T.B.; Tovilla-Zárate, C.A.; López-Narváez, M.L.; Genis-Mendoza, A.D.; Castillo-Avila, R.G.; Ramos-Méndez, M.Á.; Juárez-Rojop, I.E. Oxytocin Levels in Individuals with Schizophrenia Are High in Cerebrospinal Fluid but Low in Serum: A Systematic Review and Meta-Analysis. Metab. Brain Dis. 2021, 36, 2415–2424. [Google Scholar] [CrossRef]
- Li, Y.; Hassett, A.L.; Seng, J.S. Exploring the Mutual Regulation between Oxytocin and Cortisol as a Marker of Resilience. Arch. Psychiatr. Nurs. 2019, 33, 164–173. [Google Scholar] [CrossRef]
- Donadon, M.F.; Martin-Santos, R.; de Osório, F.L. The Associations Between Oxytocin and Trauma in Humans: A Systematic Review. Front. Pharmacol. 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Rubin, L.H.; Carter, C.S.; Drogos, L.; Pournajafi-Nazarloo, H.; Sweeney, J.A.; Maki, P.M. Peripheral Oxytocin Is Associated with Reduced Symptom Severity in Schizophrenia. Schizophr. Res. 2010, 124, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzel, D.; Yazici, A.B.; Pek, T.M.; Doganay, S.; Simsek, A.B.S.; Saglam, K.; Turan, C.; Yazici, E. Atrial Natriuretic Peptide and Posterior Pituitary Neurohormone Changes in Patients with Acute Schizophrenia. Neuropsychiatr. Dis. Treat. 2018, 14, 1855–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, L.H.; Wehring, H.J.; Demyanovich, H.; Sue Carter, C.; Pournajafi-Nazarloo, H.; Feldman, S.M.; Earl, A.K.; August, S.; Gold, J.M.; Kelly, D.L. Peripheral Oxytocin and Vasopressin Are Associated with Clinical Symptom Severity and Cognitive Functioning in Midlife Women with Chronic Schizophrenia. Schizophr. Res. 2018, 195, 409–411. [Google Scholar] [CrossRef]
- Rubin, L.H.; Carter, C.S.; Bishop, J.R.; Pournajafi-Nazarloo, H.; Drogos, L.L.; Hill, S.K.; Ruocco, A.C.; Keedy, S.K.; Reilly, J.L.; Keshavan, M.S.; et al. Reduced Levels of Vasopressin and Reduced Behavioral Modulation of Oxytocin in Psychotic Disorders. Schizophr. Bull. 2014, 40, 1374–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, K.; Keller, W.; Buchanan, R.; Gold, J.; Koenig, J.; Ossenfort, K.; Katz, A.; Strauss, G. C-14Plasma Oxytocin Levels Are Associated with Impaired Social Cognition and Neurocognition in Schizophrenia. Arch. Clin. Neuropsychol. 2014, 29, 577–578. [Google Scholar] [CrossRef] [Green Version]
- Strauss, G.P.; Keller, W.R.; Koenig, J.I.; Sullivan, S.K.; Gold, J.M.; Buchanan, R.W. Endogenous Oxytocin Levels Are Associated with the Perception of Emotion in Dynamic Body Expressions in Schizophrenia. Schizophr. Res. 2015, 162, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Strauss, G.P.; Chapman, H.C.; Keller, W.R.; Koenig, J.I.; Gold, J.M.; Carpenter, W.T.; Buchanan, R.W. Endogenous Oxytocin Levels Are Associated with Impaired Social Cognition and Neurocognition in Schizophrenia. J. Psychiatr. Res. 2019, 112, 38–43. [Google Scholar] [CrossRef]
- Goh, K.K.; Lu, M.-L. Relationship between the Domains of Theory of Mind, Social Dysfunction, and Oxytocin in Schizophrenia. J. Psychiatr. Res. 2022, 155, 420–429. [Google Scholar] [CrossRef]
- Strauss, G.P.; Keller, W.R.; Koenig, J.I.; Gold, J.M.; Ossenfort, K.L.; Buchanan, R.W. Plasma Oxytocin Levels Predict Olfactory Identification and Negative Symptoms in Individuals with Schizophrenia. Schizophr. Res. 2015, 162, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Kéri, S.; Kiss, I.; Kelemen, O. Sharing Secrets: Oxytocin and Trust in Schizophrenia. Soc. Neurosci. 2009, 4, 287–293. [Google Scholar] [CrossRef]
- Aydın, O.; Lysaker, P.H.; Balıkçı, K.; Ünal-Aydın, P.; Esen-Danacı, A. Associations of Oxytocin and Vasopressin Plasma Levels with Neurocognitive, Social Cognitive and Meta Cognitive Function in Schizophrenia. Psychiatry Res. 2018, 270, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Spilka, M.J.; Keller, W.R.; Buchanan, R.W.; Gold, J.M.; Koenig, J.I.; Strauss, G.P. Endogenous Oxytocin Levels Are Associated with Facial Emotion Recognition Accuracy but Not Gaze Behavior in Individuals with Schizophrenia. Acta Psychiatr. Scand 2022, 145, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Strauss, G.P.; Keller, W.R.; Koenig, J.I.; Gold, J.M.; Frost, K.H.; Buchanan, R.W. Plasma Oxytocin Levels Predict Social Cue Recognition in Individuals with Schizophrenia. Schizophr. Res. 2015, 162, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, L.H.; Carter, C.S.; Drogos, L.; Jamadar, R.; Pournajafi-Nazarloo, H.; Sweeney, J.A.; Maki, P.M. Sex-Specific Associations between Peripheral Oxytocin and Emotion Perception in Schizophrenia. Schizophr. Res. 2011, 130, 266–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, M.; Marlow-O’Connor, M.; Torres, I.; Carter, C.S. Diminished Plasma Oxytocin in Schizophrenic Patients with Neuroendocrine Dysfunction and Emotional Deficits. Schizophr. Res. 2008, 98, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Uhrig, S.; Hirth, N.; Broccoli, L.; von Wilmsdorff, M.; Bauer, M.; Sommer, C.; Zink, M.; Steiner, J.; Frodl, T.; Malchow, B.; et al. Reduced Oxytocin Receptor Gene Expression and Binding Sites in Different Brain Regions in Schizophrenia: A Post-Mortem Study. Schizophr. Res. 2016, 177, 59–66. [Google Scholar] [CrossRef]
- Gibson, C.M.; Penn, D.L.; Smedley, K.L.; Leserman, J.; Elliott, T.; Pedersen, C.A. A Pilot Six-Week Randomized Controlled Trial of Oxytocin on Social Cognition and Social Skills in Schizophrenia. Schizophr. Res. 2014, 156, 261–265. [Google Scholar] [CrossRef]
- Jarskog, L.F.; Pedersen, C.A.; Johnson, J.L.; Hamer, R.M.; Rau, S.W.; Elliott, T.; Penn, D.L. A 12-Week Randomized Controlled Trial of Twice-Daily Intranasal Oxytocin for Social Cognitive Deficits in People with Schizophrenia. Schizophr. Res. 2017, 185, 88–95. [Google Scholar] [CrossRef]
- Modabbernia, A.; Rezaei, F.; Salehi, B.; Jafarinia, M.; Ashrafi, M.; Tabrizi, M.; Hosseini, S.M.R.; Tajdini, M.; Ghaleiha, A.; Akhondzadeh, S. Intranasal Oxytocin as an Adjunct to Risperidone in Patients with Schizophrenia: An 8-Week, Randomized, Double-Blind, Placebo-Controlled Study. CNS Drugs 2013, 27, 57–65. [Google Scholar] [CrossRef]
- Ota, M.; Yoshida, S.; Nakata, M.; Yada, T.; Kunugi, H. The Effects of Adjunctive Intranasal Oxytocin in Patients with Schizophrenia. Postgrad. Med. 2018, 130, 122–128. [Google Scholar] [CrossRef]
- Marotta, R.F.; Buono, F.D.; Garakani, A.; Collins, E.; Cerrito, B.; David Rowe, D.R. The Effects of Augmenting Clozapine with Oxytocin in Schizophrenia: An Initial Case Series. Ann. Clin. Psychiatry 2020, 32, 90–96. [Google Scholar]
- Abram, S.V.; De Coster, L.; Roach, B.J.; Mueller, B.A.; van Erp, T.G.M.; Calhoun, V.D.; Preda, A.; Lim, K.O.; Turner, J.A.; Ford, J.M.; et al. Oxytocin Enhances an Amygdala Circuit Associated with Negative Symptoms in Schizophrenia: A Single-Dose, Placebo-Controlled, Crossover, Randomized Control Trial. Schizophr. Bull. 2020, 46, 661–669. [Google Scholar] [CrossRef]
- Korann, V.; Jacob, A.; Lu, B.; Devi, P.; Thonse, U.; Nagendra, B.; Maria Chacko, D.; Dey, A.; Padmanabha, A.; Shivakumar, V.; et al. Effect of Intranasal Oxytocin on Resting-State Effective Connectivity in Schizophrenia. Schizophr. Bull. 2022, 48, 1115–1124. [Google Scholar] [CrossRef]
- Averbeck, B.B.; Bobin, T.; Evans, S.; Shergill, S.S. Emotion Recognition and Oxytocin in Patients with Schizophrenia. Psychol. Med. 2012, 42, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Goldman, M.B.; Gomes, A.M.; Carter, C.S.; Lee, R. Divergent Effects of Two Different Doses of Intranasal Oxytocin on Facial Affect Discrimination in Schizophrenic Patients with and without Polydipsia. Psychopharmacology 2011, 216, 101–110. [Google Scholar] [CrossRef]
- Woolley, J.D.; Chuang, B.; Lam, O.; Lai, W.; O’Donovan, A.; Rankin, K.P.; Mathalon, D.H.; Vinogradov, S. Oxytocin Administration Enhances Controlled Social Cognition in Patients with Schizophrenia. Psychoneuroendocrinology 2014, 47, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Andari, E.; Massa, N.M.; Fargotstein, M.D.; Taylor, N.B.; Halverson, D.M.; Owens, A.V.; Currin, D.L.; Bhattacharya, A.; Gitman, D.; Cuthbert, B.C.; et al. Effects of Oxytocin on Emotion Recognition in Schizophrenia: A Randomized Double-Blind Pilot Study. J. Clin. Psychopharmacol. 2021, 41, 103–113. [Google Scholar] [CrossRef]
- Shin, N.Y.; Park, H.Y.; Jung, W.H.; Park, J.W.; Yun, J.-Y.; Jang, J.H.; Kim, S.N.; Han, H.J.; Kim, S.-Y.; Kang, D.-H.; et al. Effects of Oxytocin on Neural Response to Facial Expressions in Patients with Schizophrenia. Neuropsychopharmacology 2015, 40, 2286. [Google Scholar] [CrossRef] [Green Version]
- Wigton, R.; Tracy, D.K.; Verneuil, T.M.; Johns, M.; White, T.; Michalopoulou, P.G.; Averbeck, B.; Shergill, S. The Importance of Pro-Social Processing, and Ameliorating Dysfunction in Schizophrenia. An FMRI Study of Oxytocin. Schizophr. Res. Cogn. 2022, 27, 100221. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.A.; Gibson, C.M.; Rau, S.W.; Salimi, K.; Smedley, K.L.; Casey, R.L.; Leserman, J.; Jarskog, L.F.; Penn, D.L. Intranasal Oxytocin Reduces Psychotic Symptoms and Improves Theory of Mind and Social Perception in Schizophrenia. Schizophr. Res. 2011, 132, 50–53. [Google Scholar] [CrossRef]
- Feifel, D.; MacDonald, K.; Cobb, P.; Minassian, A. Adjunctive Intranasal Oxytocin Improves Verbal Memory in People with Schizophrenia. Schizophr. Res. 2012, 139, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Halverson, T.; Jarskog, L.F.; Pedersen, C.; Penn, D. Effects of Oxytocin on Empathy, Introspective Accuracy, and Social Symptoms in Schizophrenia: A 12-Week Twice-Daily Randomized Controlled Trial. Schizophr. Res. 2019, 204, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Guastella, A.J.; Ward, P.B.; Hickie, I.B.; Shahrestani, S.; Hodge, M.A.R.; Scott, E.M.; Langdon, R. A Single Dose of Oxytocin Nasal Spray Improves Higher-Order Social Cognition in Schizophrenia. Schizophr. Res. 2015, 168, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.C.; Green, M.F.; Lee, J.; Horan, W.P.; Senturk, D.; Clarke, A.D.; Marder, S.R. Oxytocin-Augmented Social Cognitive Skills Training in Schizophrenia. Neuropsychopharmacology 2014, 39, 2070–2077. [Google Scholar] [CrossRef] [Green Version]
- Abu-Akel, A.; Palgi, S.; Klein, E.; Decety, J.; Shamay-Tsoory, S. Oxytocin Increases Empathy to Pain When Adopting the Other- but Not the Self-Perspective. Soc. Neurosci. 2015, 10, 7–15. [Google Scholar] [CrossRef]
- Buchanan, R.W.; Kelly, D.L.; Weiner, E.; Gold, J.M.; Strauss, G.P.; Koola, M.M.; McMahon, R.P.; Carpenter, W.T. A Randomized Clinical Trial of Oxytocin or Galantamine for the Treatment of Negative Symptoms and Cognitive Impairments in People With Schizophrenia. J. Clin. Psychopharmacol. 2017, 37, 394–400. [Google Scholar] [CrossRef]
- Cacciotti-Saija, C.; Langdon, R.; Ward, P.B.; Hickie, I.B.; Scott, E.M.; Naismith, S.L.; Moore, L.; Alvares, G.A.; Redoblado Hodge, M.A.; Guastella, A.J. A Double-Blind Randomized Controlled Trial of Oxytocin Nasal Spray and Social Cognition Training for Young People with Early Psychosis. Schizophr. Bull. 2015, 41, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.R.; Wehring, H.J.; McMahon, R.P.; Liu, F.; Linthicum, J.; Buchanan, R.W.; Strauss, G.P.; Rubin, L.H.; Kelly, D.L. The Effect of Intranasal Oxytocin on Measures of Social Cognition in Schizophrenia: A Negative Report. J. Psychiatr. Brain Sci. 2019, 4, e190001. [Google Scholar] [CrossRef]
- Horta de Macedo, L.R.; Zuardi, A.W.; Machado-de-Sousa, J.P.; Chagas, M.H.N.; Hallak, J.E.C. Oxytocin Does Not Improve Performance of Patients with Schizophrenia and Healthy Volunteers in a Facial Emotion Matching Task. Psychiatry Res. 2014, 220, 125–128. [Google Scholar] [CrossRef]
- Bradley, E.R.; Tai, M.; Hankin, M.; Woolley, J.D. Preliminary Evidence That Oxytocin Does Not Improve Mentalizing in Women with Schizophrenia. Horm. Behav. 2021, 128, 104915. [Google Scholar] [CrossRef]
- Rosenfeld, A.J.; Lieberman, J.A.; Jarskog, L.F. Oxytocin, Dopamine, and the Amygdala: A Neurofunctional Model of Social Cognitive Deficits in Schizophrenia. Schizophr. Bull. 2011, 37, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Domes, G.; Heinrichs, M.; Michel, A.; Berger, C.; Herpertz, S.C. Oxytocin Improves “Mind-Reading” in Humans. Biol. Psychiatry 2007, 61, 731–733. [Google Scholar] [CrossRef]
- Sabe, M.; Zhao, N.; Crippa, A.; Strauss, G.P.; Kaiser, S. Intranasal Oxytocin for Negative Symptoms of Schizophrenia: Systematic Review, Meta-Analysis, and Dose-Response Meta-Analysis of Randomized Controlled Trials. Int. J. Neuropsychopharmacol. 2021, 24, 601–614. [Google Scholar] [CrossRef]
- Zheng, W.; Zhu, X.-M.; Zhang, Q.-E.; Yang, X.-H.; Cai, D.-B.; Li, L.; Li, X.-B.; Ng, C.H.; Ungvari, G.S.; Ning, Y.-P.; et al. Adjunctive Intranasal Oxytocin for Schizophrenia: A Meta-Analysis of Randomized, Double-Blind, Placebo-Controlled Trials. Schizophr. Res. 2019, 206, 13–20. [Google Scholar] [CrossRef]
- Feifel, D.; Reza, T. Oxytocin Modulates Psychotomimetic-Induced Deficits in Sensorimotor Gating. Psychopharmacology 1999, 141, 93–98. [Google Scholar] [CrossRef]
- Tapias-Espinosa, C.; Cañete, T.; Sampedro-Viana, D.; Brudek, T.; Kaihøj, A.; Oliveras, I.; Tobeña, A.; Aznar, S.; Fernández-Teruel, A. Oxytocin Attenuates Schizophrenia-like Reduced Sensorimotor Gating in Outbred and Inbred Rats in Line with Strain Differences in CD38 Gene Expression. Physiol. Behav. 2021, 240, 113547. [Google Scholar] [CrossRef]
- Bowen, M.T.; Carson, D.S.; Spiro, A.; Arnold, J.C.; McGregor, I.S. Adolescent Oxytocin Exposure Causes Persistent Reductions in Anxiety and Alcohol Consumption and Enhances Sociability in Rats. PLoS ONE 2011, 6, e27237. [Google Scholar] [CrossRef] [Green Version]
- Kohli, S.; King, M.V.; Williams, S.; Edwards, A.; Ballard, T.M.; Steward, L.J.; Alberati, D.; Fone, K.C.F. Oxytocin Attenuates Phencyclidine Hyperactivity and Increases Social Interaction and Nucleus Accumben Dopamine Release in Rats. Neuropsychopharmacology 2019, 44, 295–305. [Google Scholar] [CrossRef]
- Chang, S.W.C.; Barter, J.W.; Ebitz, R.B.; Watson, K.K.; Platt, M.L. Inhaled Oxytocin Amplifies Both Vicarious Reinforcement and Self Reinforcement in Rhesus Macaques (Macaca Mulatta). Proc. Natl. Acad. Sci. USA 2012, 109, 959–964. [Google Scholar] [CrossRef] [Green Version]
- Rault, J.-L.; Carter, C.S.; Garner, J.P.; Marchant-Forde, J.N.; Richert, B.T.; Lay, D.C. Repeated Intranasal Oxytocin Administration in Early Life Dysregulates the HPA Axis and Alters Social Behavior. Physiol. Behav. 2013, 112–113, 40–48. [Google Scholar] [CrossRef]
- Huang, H.; Michetti, C.; Busnelli, M.; Managò, F.; Sannino, S.; Scheggia, D.; Giancardo, L.; Sona, D.; Murino, V.; Chini, B.; et al. Chronic and Acute Intranasal Oxytocin Produce Divergent Social Effects in Mice. Neuropsychopharmacology 2014, 39, 1102–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bales, K.L.; Perkeybile, A.M.; Conley, O.G.; Lee, M.H.; Guoynes, C.D.; Downing, G.M.; Yun, C.R.; Solomon, M.; Jacob, S.; Mendoza, S.P. Chronic Intranasal Oxytocin Causes Long-Term Impairments in Partner Preference Formation in Male Prairie Voles. Biol. Psychiatry 2013, 74, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parr, L.A.; Modi, M.; Siebert, E.; Young, L.J. Intranasal Oxytocin Selectively Attenuates Rhesus Monkeys’ Attention to Negative Facial Expressions. Psychoneuroendocrinology 2013, 38, 1748–1756. [Google Scholar] [CrossRef] [Green Version]
- Rubin, L.H.; Li, S.; Yao, L.; Keedy, S.K.; Reilly, J.L.; Hill, S.K.; Bishop, J.R.; Sue Carter, C.; Pournajafi-Nazarloo, H.; Drogos, L.L.; et al. Peripheral Oxytocin and Vasopressin Modulates Regional Brain Activity Differently in Men and Women with Schizophrenia. Schizophr. Res. 2018, 202, 173–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gossen, A.; Hahn, A.; Westphal, L.; Prinz, S.; Schultz, R.T.; Gründer, G.; Spreckelmeyer, K.N. Oxytocin Plasma Concentrations after Single Intranasal Oxytocin Administration—A Study in Healthy Men. Neuropeptides 2012, 46, 211–215. [Google Scholar] [CrossRef]
- Hiroi, R.; Lacagnina, A.F.; Hinds, L.R.; Carbone, D.G.; Uht, R.M.; Handa, R.J. The Androgen Metabolite, 5α-Androstane-3β,17β-Diol (3β-Diol), Activates the Oxytocin Promoter through an Estrogen Receptor-β Pathway. Endocrinology 2013, 154, 1802–1812. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.M.; Young, L.J. Neuropeptidergic Regulation of Affiliative Behavior and Social Bonding in Animals. Horm. Behav. 2006, 50, 506–517. [Google Scholar] [CrossRef]
- Richard, S.; Zingg, H.H. The Human Oxytocin Gene Promoter Is Regulated by Estrogens. J. Biol. Chem. 1990, 265, 6098–6103. [Google Scholar] [CrossRef]
- Iovino, M.; Messana, T.; Tortora, A.; Giusti, C.; Lisco, G.; Giagulli, V.A.; Guastamacchia, E.; De Pergola, G.; Triggiani, V. Oxytocin Signaling Pathway: From Cell Biology to Clinical Implications. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 91–110. [Google Scholar] [CrossRef]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and Estrogen Receptor β in the Brain: An Overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.; Müller, L.E.; Ditzen, B.; Herpertz, S.C.; Bertsch, K. Oxytocin and Social Anxiety: Interactions with Sex Hormones. Psychoneuroendocrinology 2021, 128, 105224. [Google Scholar] [CrossRef]
- Frankiensztajn, L.M.; Gur-Pollack, R.; Wagner, S. A Combinatorial Modulation of Synaptic Plasticity in the Rat Medial Amygdala by Oxytocin, Urocortin3 and Estrogen. Psychoneuroendocrinology 2018, 92, 95–102. [Google Scholar] [CrossRef]
- Kolatorova, L.; Vitku, J.; Suchopar, J.; Hill, M.; Parizek, A. Progesterone: A Steroid with Wide Range of Effects in Physiology as Well as Human Medicine. Int. J. Mol. Sci. 2022, 23, 7989. [Google Scholar] [CrossRef]
- Yamamoto, K.; Nakano, Y.; Iwata, N.; Soejima, Y.; Suyama, A.; Hasegawa, T.; Otsuka, F. Oxytocin Enhances Progesterone Production with Upregulation of BMP-15 Activity by Granulosa Cells. Biochem. Biophys. Res. Commun. 2023, 646, 103–109. [Google Scholar] [CrossRef]
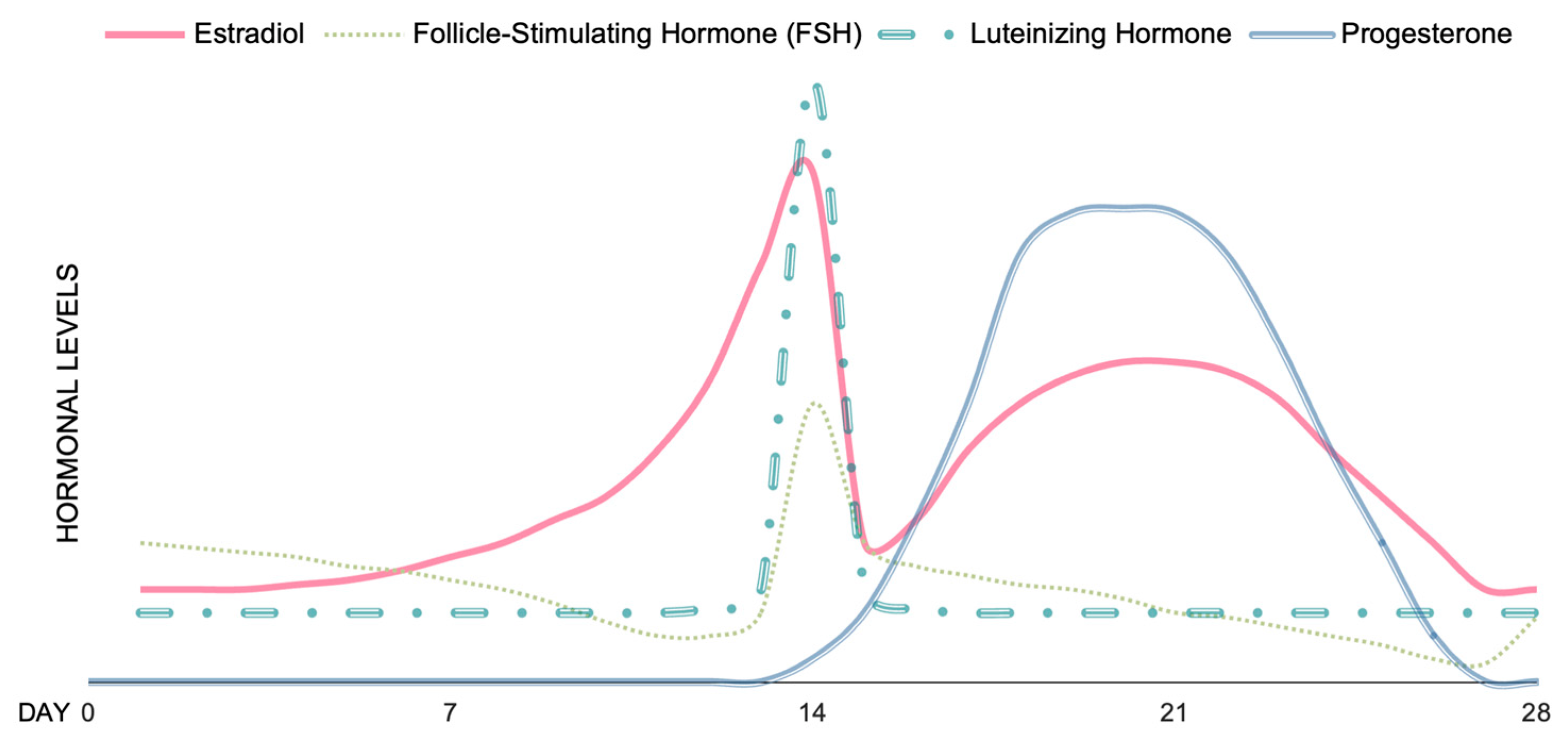
- Reed, B.G.; Carr, B.R. The Normal Menstrual Cycle and the Control of Ovulation. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Hofland, J., Dungan, K., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Draper, C.F.; Duisters, K.; Weger, B.; Chakrabarti, A.; Harms, A.C.; Brennan, L.; Hankemeier, T.; Goulet, L.; Konz, T.; Martin, F.P.; et al. Menstrual Cycle Rhythmicity: Metabolic Patterns in Healthy Women. Sci. Rep. 2018, 8, 14568. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-P.; Wang, L.; Han, L.; Wang, S.C. Nonsocial Functions of Hypothalamic Oxytocin. Int. Sch. Res. Not. 2013, 2013, e179272. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, A.; Yamamoto, Y.; Noguchi, H.; Takeda, A.; Tamura, K.; Aoki, H.; Minato, S.; Uchishiba, M.; Yamamoto, S.; Kamada, S.; et al. Changes in Serum Oxytocin Levels under Physiological and Supraphysiological Gonadal Steroid Hormone Conditions in Women of Reproductive Age: A Preliminary Study. Nutrients 2022, 14, 5350. [Google Scholar] [CrossRef]
- Riecher-Rössler, A.; Butler, S.; Kulkarni, J. Sex and Gender Differences in Schizophrenic Psychoses—A Critical Review. Arch. Womens Ment. Health 2018, 21, 627–648. [Google Scholar] [CrossRef]
- Felgel-Farnholz, V.; Hlusicka, E.B.; Edemann-Callesen, H.; Garthe, A.; Winter, C.; Hadar, R. Adolescent Raloxifene Treatment in Females Prevents Cognitive Deficits in a Neurodevelopmental Rodent Model of Schizophrenia. Behav. Brain Res. 2023, 441, 114276. [Google Scholar] [CrossRef]
- Brzezinski-Sinai, N.A.; Brzezinski, A. Schizophrenia and Sex Hormones: What Is the Link? Front. Psychiatry 2020, 11, 693. [Google Scholar] [CrossRef]
- Sezer, E.; Köşger, F.; Altınöz, A.E.; Yiğitaslan, S. Relationship Between Gonadal Hormone Levels and Symptom Severity in Female Patients with Schizophrenia. Alpha Psychiatry 2021, 22, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Kang, S.-B.; Joe, S.-H. Changes in Premenstrual Symptoms in Women with Schizophrenia: A Prospective Study. Psychosom. Med. 2001, 63, 822. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Mandal, N.; Sinha, V.K. Change of Symptoms of Schizophrenia across Phases of Menstrual Cycle. Arch. Womens Ment. Health 2020, 23, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.J.; Sagnay de la Bastida, V.C.; Joyce, D.W.; Cullen, A.E.; McGuire, P. Exacerbation of Psychosis During the Perimenstrual Phase of the Menstrual Cycle: Systematic Review and Meta-Analysis. Schizophr. Bull. 2020, 46, 78–90. [Google Scholar] [CrossRef]
- Seeman, M.V. Menstrual Exacerbation of Schizophrenia Symptoms. Acta. Psychiatr. Scand. 2012, 125, 363–371. [Google Scholar] [CrossRef]
- Begemann, M.J.H.; Dekker, C.F.; van Lunenburg, M.; Sommer, I.E. Estrogen Augmentation in Schizophrenia: A Quantitative Review of Current Evidence. Schizophr. Res. 2012, 141, 179–184. [Google Scholar] [CrossRef]
- Heringa, S.M.; Begemann, M.J.H.; Goverde, A.J.; Sommer, I.E.C. Sex Hormones and Oxytocin Augmentation Strategies in Schizophrenia: A Quantitative Review. Schizophr. Res. 2015, 168, 603–613. [Google Scholar] [CrossRef]
- Brand, B.A.; de Boer, J.N.; Sommer, I.E.C. Estrogens in Schizophrenia: Progress, Current Challenges and Opportunities. Curr. Opin. Psychiatry 2021, 34, 228–237. [Google Scholar] [CrossRef]
- de Boer, J.; Prikken, M.; Lei, W.U.; Begemann, M.; Sommer, I. The Effect of Raloxifene Augmentation in Men and Women with a Schizophrenia Spectrum Disorder: A Systematic Review and Meta-Analysis. NPJ Schizophr. 2018, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.-M.; Zheng, W.; Li, X.-H.; Cai, D.-B.; Yang, X.-H.; Ungvari, G.S.; Ng, C.H.; Wang, X.-P.; Kulkarni, J.; Grigg, J.; et al. Adjunctive Raloxifene for Postmenopausal Women with Schizophrenia: A Meta-Analysis of Randomized, Double-Blind, Placebo-Controlled Trials. Schizophr. Res. 2018, 197, 288–293. [Google Scholar] [CrossRef]
- Seeman, M.V. Men and Women Respond Differently to Antipsychotic Drugs. Neuropharmacology 2020, 163, 107631. [Google Scholar] [CrossRef]
- Vandegrift, B.J.; You, C.; Satta, R.; Brodie, M.S.; Lasek, A.W. Estradiol Increases the Sensitivity of Ventral Tegmental Area Dopamine Neurons to Dopamine and Ethanol. PLoS ONE 2017, 12, e0187698. [Google Scholar] [CrossRef] [Green Version]
- Brand, B.A.; Haveman, Y.R.A.; de Beer, F.; de Boer, J.N.; Dazzan, P.; Sommer, I.E.C. Antipsychotic Medication for Women with Schizophrenia Spectrum Disorders. Psychol. Med. 2022, 52, 649–663. [Google Scholar] [CrossRef]
- Jönsson, A.K.; Spigset, O.; Reis, M. A Compilation of Serum Concentrations of 12 Antipsychotic Drugs in a Therapeutic Drug Monitoring Setting. Drug Monit. 2019, 41, 348–356. [Google Scholar] [CrossRef]
- Edinoff, A.N.; Silverblatt, N.S.; Vervaeke, H.E.; Horton, C.C.; Girma, E.; Kaye, A.D.; Kaye, A.; Kaye, J.S.; Garcia, A.J.; Neuchat, E.E.; et al. Hyperprolactinemia, Clinical Considerations, and Infertility in Women on Antipsychotic Medications. Psychopharmacol. Bull. 2021, 51, 131–148. [Google Scholar]
- González-Rodríguez, A.; Seeman, M.V. The Association between Hormones and Antipsychotic Use: A Focus on Postpartum and Menopausal Women. Adv. Psychopharmacol. 2019, 9, 2045125319859973. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Chen, M.; Chen, C.; Lin, X.; Jiang, D.; Zhang, Y.; Wang, L.; Zhuo, C.; Tian, H.; Du, C. Efficacy and Acceptability of Three Prolactin-Sparing Antipsychotics in Patient with Schizophrenia: A Network Meta-Analysis. Psychiatry Clin. Psychopharmacol. 2019, 29, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Gogos, A.; Sbisa, A.M.; Sun, J.; Gibbons, A.; Udawela, M.; Dean, B. A Role for Estrogen in Schizophrenia: Clinical and Preclinical Findings. Int. J. Endocrinol. 2015, 2015, e615356. [Google Scholar] [CrossRef] [Green Version]
- Augustine, R.A.; Ladyman, S.R.; Bouwer, G.T.; Alyousif, Y.; Sapsford, T.J.; Scott, V.; Kokay, I.C.; Grattan, D.R.; Brown, C.H. Prolactin Regulation of Oxytocin Neurone Activity in Pregnancy and Lactation. J. Physiol. 2017, 595, 3591–3605. [Google Scholar] [CrossRef] [Green Version]
- Marsh, N.; Marsh, A.A.; Lee, M.R.; Hurlemann, R. Oxytocin and the Neurobiology of Prosocial Behavior. Neuroscientist 2021, 27, 604–619. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol. Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Shilling, P.D.; Feifel, D. Potential of Oxytocin in the Treatment of Schizophrenia. CNS Drugs 2016, 30, 193–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valstad, M.; Alvares, G.A.; Egknud, M.; Matziorinis, A.M.; Andreassen, O.A.; Westlye, L.T.; Quintana, D.S. The Correlation between Central and Peripheral Oxytocin Concentrations: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2017, 78, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, I.D.; Landgraf, R. Balance of Brain Oxytocin and Vasopressin: Implications for Anxiety, Depression, and Social Behaviors. Trends Neurosci. 2012, 35, 649–659. [Google Scholar] [CrossRef]
- Caldwell, H.K. Oxytocin and Sex Differences in Behavior. Curr. Opin. Behav. Sci. 2018, 23, 13–20. [Google Scholar] [CrossRef]
- Butlen-Ducuing, F.; Balkowiec-Iskra, E.; Dalla, C.; Slattery, D.A.; Ferretti, M.T.; Kokras, N.; Balabanov, P.; De Vries, C.; Mellino, S.; Santuccione Chadha, A. Implications of Sex-Related Differences in Central Nervous System Disorders for Drug Research and Development. Nat. Rev. Drug Discov. 2021, 20, 881–882. [Google Scholar] [CrossRef]
- Garforth, B.; Degnbol, H.; Terris, E.T.; Zak, P.J.; Winterdahl, M. Elevated Plasma Oxytocin Levels and Higher Satisfaction with Life in Young Oral Contraceptive Users. Sci. Rep. 2020, 10, 8208. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author, Year | Design | Subjects N (M/F) | Trial Duration | OXT Administration | Testing after Dose | Outcome Measures | Results (OXT) | Conclusions * |
|---|---|---|---|---|---|---|---|---|
| Davis et al., 2013 [18] | RDBPC | 23 (23M) SCHZ | 3 sessions | 40 IU or placebo; intranasal | 30 min (3rd session) | Social cognition | Improved inferential and regulatory processes. | Acute therapeutic effects on higher-order social cognition. |
| Feifel et al., 2010 [19] | RDBPC/CO | 15 (12M, 3F) SCHZ | 7 weeks | 2 × 20 IU daily for 1 week, 2 × 40 IU thereafter or placebo; intranasal | N/A | Clinical symptoms, symptom severity | Clinical symptom improvement at 3-weeks. | Antipsychotic properties, with a delayed onset of action. |
| Gibson et al., 2014 [58] | RDBPC | 14 (11M, 3F) SCHZ | 6 weeks | 2 × 24 IU or placebo; intranasal | 50 min after the AM dose at the end of week 6 | Clinical symptoms, social cognition, and skills | Improved negative symptoms, fear recognition, self-reported perspective taking. | Limited efficacy on social cognition and social skills. |
| Jarskog et al., 2017 [59] | RDBPC | 62 (47M, 15F) SCHZ or schizoaffective disorder | 12 weeks | 2 × 24 IU or placebo; intranasal | N/A | Clinical symptoms, social cognition and skills | Improved social skills and negative symptoms. | No significant effect on sociocognitive function. |
| Modabbernia et al., 2013 [60] | RDBPC | 40 (33M, 7F) SCHZ, stable on risperidone | 8 weeks | 2 × 20 IU daily for 1 week, 2 × 40 thereafter or placebo; intranasal | N/A | Clinical symptoms | Clinical symptom improvement at 8-week end point. | Therapeutic effect additive to risperidone treatment. |
| Ota et al., 2018 [61] | NROL | 16 (7M, 9F) chronic SCHZ | 12 weeks | 2 × 12 IU daily; intranasal | N/A | Clinical symptoms, cognition, social cognition, brain structure (MRI) | Clinical symptom improvement, negative correlation with gray matter of right insula and left cingulate cortex, verbal fluency improvement. | Therapeutic effect associated with gray matter volume changes. |
| Marotta et al., 2020 [62] | Retrospective Medical Chart Review | 5 (4M, 1F) treatment resistant SCHZ | 1 year | 10 IU–3 × 20 IU daily; sublingual (+ clozapine, 200–600 mg) | N/A | Clinical symptoms, social functioning | Reduced negative, maintained low positive symptoms, and improved occupational and social functioning. | Augmentation of clozapine with OXT effective in reducing negative symptoms. |
| Abram et al., 2020 [63] | RDBPC/CO | 22 (22M) SCHZ, 24 (24M) HC | 2 sessions, 2 weeks apart | 40 IU or placebo; intranasal | 90 min | Functional connectivity | Enhanced functional connectivity between amygdala and MTG/STS/AngG | OXT can normalize amygdala circuit associated with negative symptoms. |
| Korann et al., 2022 [64] | PC | 31 (31M) SCHZ, 21 (21M) HC | 2 sessions | 24 IU or placebo; intranasal | 45 min | Functional connectivity | Enhanced connectivity between left caudate, left supplementary motor area, left precentral gyrus, left frontal triangular gyrus | Enhanced connectivity in regions associated with cognitive insight and negative symptoms. |
| Averbeck et al., 2012 [65] | RDBPC/CO | 21 (21M) SCHZ | 2 sessions, approx. 7–8 days apart | 24 IU or saline placebo; intranasal | 50 min | Emotion recognition | Improved recognition of basic emotions. | Acute therapeutic effect on emotion recognition. |
| Goldman et al., 2011 [66] | RDBPC | 5 (3M, 2F) polydipsic SCHZ, 8 (4M, 4F) non-polydipsic SCHZ, 11 (4M, 7F) HC | 3 sessions, approx. 7 days apart | 10 IU or 20 IU or placebo; intranasal | 45 min | Emotion recognition | Low dose increases true and false positives, high dose reduces false positives in polydipsic patients. | Emotion recognition improvement after 20 IU. |
| Woolley et al., 2014 [67] | RDBPC/CO | 29 (29M) chronic SCHZ, 31 (31M) HC | 2 sessions, approx. 1 week apart | 40 IU or saline placebo; intranasal | 30 min | Social cognition | Improved controlled (not automatic) social cognition. | Improved comprehension of indirectly expressed emotions, thoughts, and intentions. |
| Andari et al., 2021 [68] | RDBPC | 20 (20M) SCHZ, 19 (19M) HC | 1 session | 24 IU or placebo; intranasal | 45 min | Emotion recognition | Enhanced emotion recognition during emotion-based ball-tossing game. | Acute low-dose OXT modest effect on social cue processing. |
| Shin et al., 2015 [69] | RDBPC/CO | 16 (16M) SCHZ, 16 (16M) HC | 2 sessions, 1 week apart | 40 IU or placebo; intranasal | 45 min | Amygdala reactivity | Amygdala reactivity increased for happy and decreased for fearful faces. | Attenuated amygdala reactivity in SCHZ, but increased reactivity in HC. |
| Wigton et al., 2022 [70] | DBPC/CO | 20 (20M) SCHZ or schizoaffective disorder | 2 sessions, 1 week apart | 40 IU or saline placebo; intranasal | 45 min (fMRI testing), 90 min (fMRI task) | Social cognition, neural activity (fMRI) | Attenuated bias for happy faces and attenuated neural activity in right insula, bilateral temporal gyri, and amygdala. | Prosocial properties supported. |
| Pedersen et al., 2011 [71] | RDBPC | 20 (17M, 3F) Paranoid or undifferentiated SCHZ | 2 weeks | 2 × 24 IU or placebo daily; intranasal | 50 min after the AM dose on day 14 | Clinical symptoms, social cognition | Improved identification of second-order false belief, clinical symptoms, suspiciousness, anxiety, and paranoia. | Therapeutic effect on social cognition supported. |
| Feifel et al., 2012 [72] | RDBPC/CO | 15 (12M, 3F) SCHZ | 6 weeks | 2 × 20 IU daily for 1 week, 2 × 40 IU thereafter or placebo; intranasal | N/A | Cognition | Improved verbal memory: total recall, short delayed free recall, and total recall discrimination. | Supported therapeutic effect on cognition. |
| Halverson et al., 2019 [73] | RDBPC | 68 (68M) SCHZ or schizoaffective disorder | 12 weeks | 2 × 24 IU or placebo; intranasal | N/A | Social cognition, empathy, social behavior | Improvement on Interpersonal Reactivity index, Perspective-Taking Subscale | Little evidence for therapeutic efficacy on social symptoms, empathy, and introspective accuracy. |
| Guastella et al., 2015 [74] | RDBPC/CO | 22 (22M) SCHZ or schizoaffective disorder | 2 sessions, 2 weeks apart | 24 IU or placebo; intranasal | 45 min | Social cognition, neurocognition | Improvement on the DANVA paralinguistic scale, and higher-order social cognition. | Acute therapeutic effects on higher-order social cognition. |
| Davis et al., 2014 [75] | RDBPC | 27 (27M) SCHZ | 4 sessions | 40 IU or placebo; intranasal | 30 min (2nd session) | Clinical symptoms, social cognition, neurocognition | Improved social cognition, facial emotion recognition, empathy, emotional intelligence. | Improved empathy with combination of OXT and social skills training. |
| Abu-Akel et al., 2015 [76] | RDBPC/CO | 29 (19M, 10F) Healthy Participants | 2 sessions, 7 days apart | 24 IU or placebo; intranasal | 45 min | Empathy | Enhanced empathy to pain from perspective of others. | Consideration of social salience hypothesis. |
| Buchanan et al., 2017 [77] | RDBPC | 58 (47M, 11F) SCHZ or schizoaffective disorder | 6 weeks | 2 × 24 IU daily; intranasal | N/A | Negative symptoms, cognition | No group differences for cognitive and negative symptoms. | No evidence for OXT’s therapeutic efficacy. |
| Cacciotti-Saija et al., 2015 [78] | RDBPC | 52 (36M, 16F) SCHZ | 6 weeks | 2 × 24 IU daily (+24 IU prior to each weekly session); intranasal | 15 min | Social cognition and functioning, symptom severity | Association between spray usage and change in negative symptoms. | Reduced negative symptoms over time, no improvement in social cognition, functioning, and symptom severity. |
| Lee et al., 2019 [79] | RDBPC | 28 (20M/8F) SCHZ or schizoaffective disorder | 3 weeks | 2 × 20 IU or placebo; intranasal | N/A | Social cognition, social functioning | No difference between treatment groups | No evidence for OXT’s therapeutic efficacy. |
| Horta de Macedo et al., 2014 [80] | RDBPC | 20 (20M) SCHZ, 20 (20M) HC | 2 sessions, 15 days apart | 48 IU; intranasal | 50 min | Emotion recognition | No effects on facial emotion recognition. | No evidence for therapeutic efficacy. |
| Bradley et al., 2021 [81] | RDBPC/CO | 26 (25F) SCHZ, 38 (38F) HC | 1 session | 40 IU or placebo; intranasal | 45 min | Mentalizing | No evidence of OXT’s effect on mentalizing | OXT treatment may have sex-specific effects |
| Author, Year | Animal Model | Trial Duration | OXT Administration | Outcome Measures | Results (OXT) | Conclusions * |
|---|---|---|---|---|---|---|
| Ferguson et al., 2002 [14] | Male OXT −/− and OXT +/+ (Hybrid mice constructed from 129S7/SvEvBrd-Hprt^b-m2 and C57BL/6J background strains, N = 42 per genotype) | 4 administrations, 48–72 h apart | 1 ng; intracerebroventricular | Social memory | OXT −/− mice no social memory, OXT +/+ mice intact social memory; OXT restored social memory, OXT antagonist produced social amnesia. | OXT essential in development of social memory in mice. |
| Feifel and Reza, 1999 [86] | Male Sprague-Dawley rats (N = 32); weight 225–250 g | 4 sessions, 7 days apart | 0.04–1 mg/kg; subcutaneous | sensorimotor gating (intact ppi and ppi disrupted by apomorphine, amphetamine, and dizocilpine). | OXT dose-dependently antagonizing effects of amphetamine and dizocilpine on PPI. | OXT receptor may modulate dopaminergic and glutamatergic regulation of PPI. |
| Tapias-Espinosa et al., 2021 [87] | Naïve male HS (“National Institutes of Health genetically heterogeneous” rat stock, N = 46), inbred Roman high-avoidance (N = 54) and Roman low-avoidance (N = 45) rats; 3–4 months; weight 320–390 g | HS rats: 2 sessions, 7 days apart; RHA, RLA rats: 1 session | 0.04 mg/kg or 0.2 mg/kg; subcutaneous | Sensorimotor gating (PPI), oxytocinergic mechanisms (OXTR and CD38) in medial prefrontal cortex (mPFC). | Increased PPI in HS rats, attenuated PPI deficits in RHA rats, no effect in RLA rats; increased OXTR expression in RHA and RLA rats. | Antipsychotic-like effects likely related to OXT-related gene expression influences in mPFC. |
| Bowen et al., 2011 [88] | Male Australian Albino Wistar (AAW) rats (N = 48); PND 33 at dosing; weight 127–177 g | 10 days (PND 33–42) | 1 mg/kg; intraperitoneal | Physiological and behavioral effect during a key developmental epoch. | Reduced anxiety-like behavior (PND 50); Enhanced social interaction (PND 55). | Effect on sociability and anxiety reduction supported. |
| Kohli et al., 2019 [89] | Male Lister-hooded rats (N = 56); weight 150–200 g | 4 sessions | 0.03–0.3 mg/kg; subcutaneous | Locomotor activity, core body temperature, social behavior | Enhanced social interaction between unfamiliar rats, and nucleus accumbens dopamine release; attenuated hyperactivity | Therapeutic potential of oxytocin in social behavior. |
| Chang et al., 2012 [90] | Male rhesus macaques (Macaca mulatta) (N = 2) | 12 OXT, 10 saline placebo sessions (alternating days) | 25 IU; intranasal | Social cognition | Enhanced preference and attention to reward other monkey over time, enhanced prosocial choices. | Enhanced social donation behavior, other-oriented attention, decision times. |
| Rault et al., 2013 [91] | Male (castrated) and female pigs (progenies of Yorkshite x Landrace dams bred to Duroc sires, N = 43); 1–3 days of age | 3 days | 24 IU; intranasal | Social stress | Increased aggression in social mixing, greater cortisol concentrations. | Long-term dysregulation of HPA axis, increased aggression, and decreased social contact. |
| Huang et al., 2014 [92] | Male C57BL/6J mice; 12 and 20 weeks of age | 7–21 days (chronic treatment); 1 session (acute treatment) | 2 × 0.15 IU or 0.3 IU; intranasal | Social behavior | Acute treatment increased social behavior to opposite-sex unfamiliar subjects (vs. same-sex unfamiliar subjects). | Different social behavior effect of chronic vs. acute administration. |
| Bales et al., 2013 [93] | Prairie voles (Microtus ochrogaster) (N = 89) | 21 days | 0.08–8.0 IU; intranasal | Social behavior | Acute treatment enhanced social behavior; chronic treatment reduced partner preference behavior | Difference in long-term and short-term OXT treatment. |
| Parr et al., 2013 [94] | Rhesus macaques (4M, 2F); | 1 session | 48 IU; intranasal | Social perception | Reduced attention to negative stimuli. | Monkey social perception mediated by OXT. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadea, D.; Dalla, C.; Tata, D.A. Exploring a Possible Interplay between Schizophrenia, Oxytocin, and Estrogens: A Narrative Review. Brain Sci. 2023, 13, 461. https://doi.org/10.3390/brainsci13030461
Papadea D, Dalla C, Tata DA. Exploring a Possible Interplay between Schizophrenia, Oxytocin, and Estrogens: A Narrative Review. Brain Sciences. 2023; 13(3):461. https://doi.org/10.3390/brainsci13030461
Chicago/Turabian StylePapadea, Danae, Christina Dalla, and Despina A. Tata. 2023. "Exploring a Possible Interplay between Schizophrenia, Oxytocin, and Estrogens: A Narrative Review" Brain Sciences 13, no. 3: 461. https://doi.org/10.3390/brainsci13030461
APA StylePapadea, D., Dalla, C., & Tata, D. A. (2023). Exploring a Possible Interplay between Schizophrenia, Oxytocin, and Estrogens: A Narrative Review. Brain Sciences, 13(3), 461. https://doi.org/10.3390/brainsci13030461