
Irritability Moderates the Association between Cognitive Flexibility Task Performance and Related Prefrontal Cortex Activation in Young Children

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cognitive Flexibility Task
2.3. fNIRS Data Acquisition and Preprocessing
2.4. Parent-Rated Irritability
2.5. Analysis Strategy
3. Results
3.1. Cognitive Flexibility Task Performance
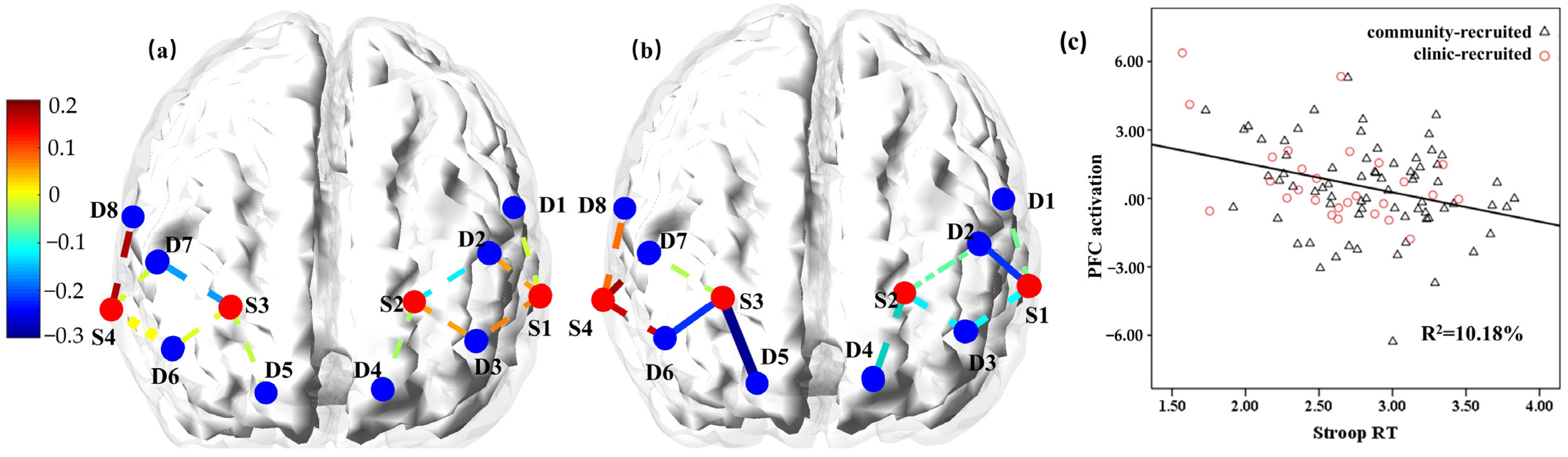
3.2. Association between the Cognitive Flexibility Task Performance and the PFC Activation
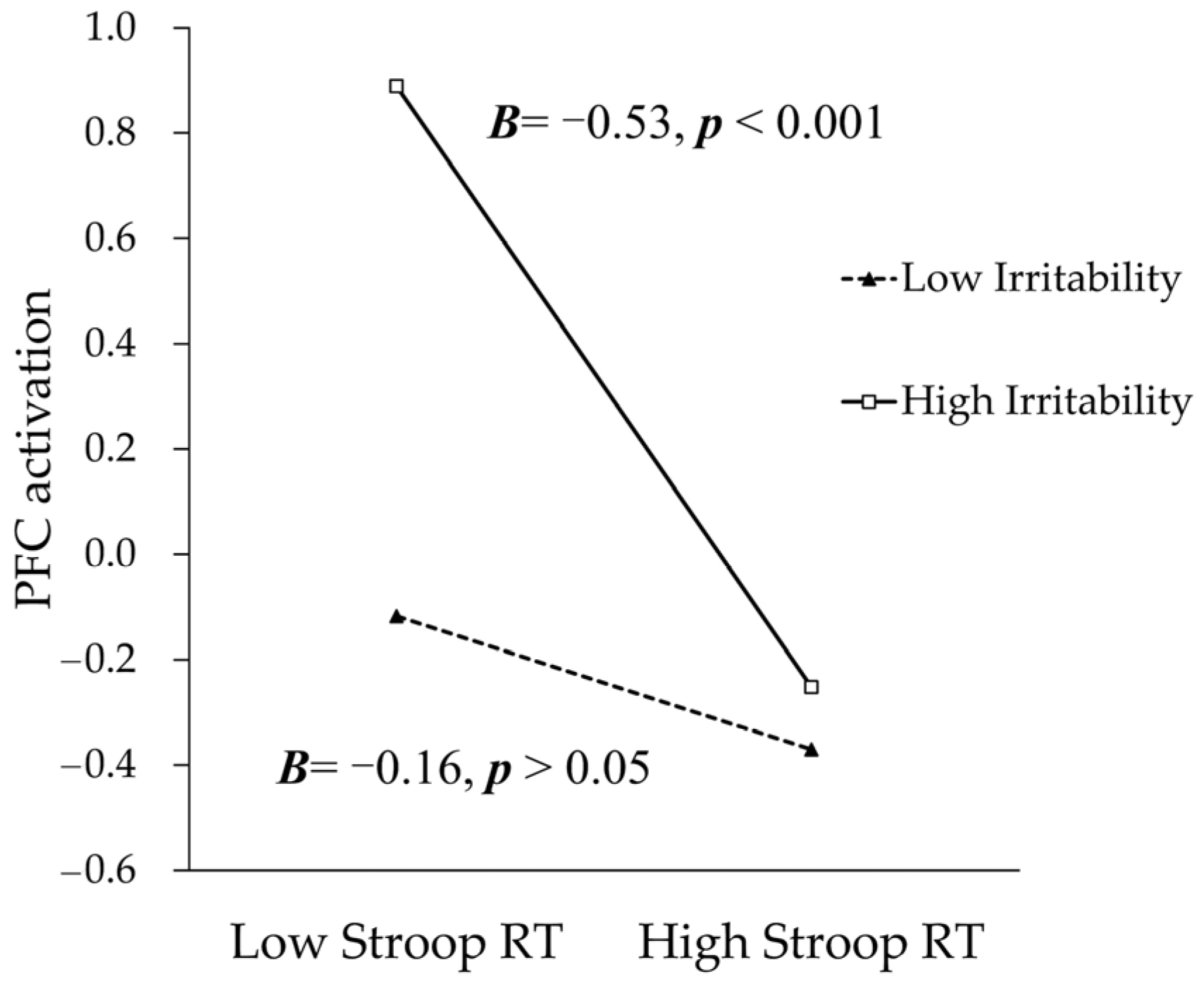
3.3. The Moderating Role of Irritability in the Association between the Cognitive Flexibility Task Performance and PFC Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, W.A. Cognitive complexity and cognitive flexibility. Sociometry 1962, 25, 405–414. [Google Scholar] [CrossRef]
- Uddin, L.Q. Cognitive and behavioural flexibility: Neural mechanisms and clinical considerations. Nat. Rev. Neurosci. 2021, 22, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, S.; Carneiro, L.; Limpo, T.; Filipe, M. Executive functions predict literacy and mathematics achievements: The unique contribution of cognitive flexibility in grades 2, 4, and 6. Child Neuropsychol. 2020, 26, 934–952. [Google Scholar] [CrossRef] [PubMed]
- Toraman, Ç.; Özdemir, H.F.; Kosan, A.M.A.; Orakci, S. Relationships between cognitive flexibility, perceived quality of faculty life, learning approaches, and academic achievement. Int. J. Instr. 2020, 13, 85–100. [Google Scholar] [CrossRef]
- Arán Filippetti, V.; Krumm, G. A hierarchical model of cognitive flexibility in children: Extending the relationship between flexibility, creativity and academic achievement. Child Neuropsychol. 2020, 26, 770–800. [Google Scholar] [CrossRef] [PubMed]
- Laureiro-Martínez, D.; Brusoni, S. Cognitive flexibility and adaptive decision-making: Evidence from a laboratory study of expert decision makers. Strateg. Manag. J. 2018, 39, 1031–1058. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Hughes, J.D.; Steinberg, B.A.; Lewis, L.D.; Heilman, K.M. Noradrenergic modulation of cognitive flexibility in problem solving. Neuroreport 1999, 10, 2763–2767. [Google Scholar] [CrossRef]
- Ciairano, S.; Bonino, S.; Miceli, R. Cognitive flexibility and social competence from childhood to early adolescence. Cogn. Creier Comport./Cogn. Brain Behav. 2006, 10, 343–366. Available online: https://www.cbbjournal.ro/index.php/en/2006/49-10-3/300-cognitive-flexibility-and-social-competence-from-childhood-to-early-adolescence (accessed on 21 March 2023).
- Veraksa, N.E.; Airapetyan, Z.V.; Gavrilova, M.N.; Fominykh, A.Y. Associations between Cognitive Outcomes and Emotion Recognition among Preschoolers. In Proceedings of the ARPHA Proceedings, Kazan Federal, Russia, 27 May–9 June 2020; pp. 2683–2693. [Google Scholar] [CrossRef]
- Odacı, H.; Cikrikci, Ö. Cognitive Flexibility Mediates the Relationship between Big Five Personality Traits and Life Satisfaction. Appl. Res. Qual. Life 2019, 14, 1229–1246. [Google Scholar] [CrossRef]
- Genet, J.J.; Siemer, M. Flexible control in processing affective and non-affective material predicts individual differences in trait resilience. Cogn. Emot. 2011, 25, 380–388. [Google Scholar] [CrossRef]
- Bunge, S.A.; Crone, E.A. Neural correlates of the development of cognitive control. In Neuroimaging in Developmental Clinical Neuroscience; Cambridge University Press: Cambridge, UK, 2009; pp. 22–37. [Google Scholar] [CrossRef]
- Prakash, R.S.; Erickson, K.I.; Colcombe, S.J.; Kim, J.S.; Voss, M.W.; Kramer, A.F. Age-related differences in the involvement of the prefrontal cortex in attentional control. Brain Cogn. 2009, 71, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.; Ischebeck, A.; Koppelstaetter, F.; Gotwald, T.; Kaufmann, L. Developmental trajectories of magnitude processing and interference control: An FMRI study. Cerebral Cortex. 2009, 19, 2755–2765. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Hiraki, K. Longitudinal development of prefrontal function during early childhood. Dev. Cogn. Neurosci. 2011, 1, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, Y.; Hiraki, K. Neural origin of cognitive shifting in young children. Proc. Natl. Acad. Sci. USA 2009, 106, 6017–6021. [Google Scholar] [CrossRef] [PubMed]
- Quiñones-Camacho, L.E.; Fishburn, F.A.; Camacho, M.C.; Wakschlag, L.S.; Perlman, S.B. Cognitive flexibility-related prefrontal activation in preschoolers: A biological approach to temperamental effortful control. Dev. Cogn. Neurosci. 2019, 38, 100651. [Google Scholar] [CrossRef]
- Li, Y.; Grabell, A.S.; Wakschlag, L.S.; Huppert, T.J.; Perlman, S.B. The neural substrates of cognitive flexibility are related to individual differences in preschool irritability: A fNIRS investigation. Dev. Cogn. Neurosci. 2017, 25, 138–144. [Google Scholar] [CrossRef]
- Ghahremani, D.G.; Monterosso, J.; Jentsch, J.D.; Bilder, R.M.; Poldrack, R.A. Neural components underlying behavioral flexibility in human reversal learning. Cerebral. Cortex. 2010, 20, 1843–1852. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, J.; Zhang, L.; Xiang, Y.-T.; Yuan, Z. Linking brain activation to topological organization in the frontal lobe as a synergistic indicator to characterize the difference between various cognitive processes of executive functions. Neurophotonics 2019, 6, 025008. [Google Scholar] [CrossRef]
- Shibuya-Tayoshi, S.; Sumitani, S.; Kikuchi, K.; Tanaka, T.; Tayoshi, S.; Ueno, S.; Ohmori, T. Activation of the prefrontal cortex during the Trail-Making Test detected with multichannel near-infrared spectroscopy. Psychiatry Clin. Neurosci. 2007, 61, 616–621. [Google Scholar] [CrossRef]
- Park, J.; Moghaddam, B. Impact of anxiety on prefrontal cortex encoding of cognitive flexibility. Neuroscience 2017, 345, 193–202. [Google Scholar] [CrossRef]
- Kennis, M.; Rademaker, A.R.; Geuze, E. Neural correlates of personality: An integrative review. Neurosci. Biobehav. Rev. 2013, 37, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Hartley, C.A.; Phelps, E.A. Anxiety and decision-making. Biol. Psychiatry 2012, 72, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Stringaris, A. Irritability in children and adolescents: A challenge for DSM-5. Eur. Child Adolesc. Psychiatry 2011, 20, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Leibenluft, E.; Stoddard, J. The developmental psychopathology of irritability. Dev. Psychopathol. 2013, 25, 1473–1487. [Google Scholar] [CrossRef]
- Blair, R.J.R.; Bashford-Largo, J.; Zhang, R.; Lukoff, J.; Elowsky, J.S.; Leibenluft, E.; Hwang, S.; Dobbertin, M.; Blair, K.S. Temporal Discounting Impulsivity and Its Association with Conduct Disorder and Irritability. J. Child Adolesc. Psychopharmacol. 2020, 30, 542–548. [Google Scholar] [CrossRef]
- Yen, J.Y.; Chang, S.J.; Long, C.Y.; Tang, T.C.; Chen, C.C.; Yen, C.F. Working memory deficit in premenstrual dysphoric disorder and its associations with difficulty in concentrating and irritability. Compr. Psychiatry 2012, 53, 540–545. [Google Scholar] [CrossRef]
- Wakschlag, L.S.; Estabrook, R.; Petitclerc, A.; Henry, D.; Burns, J.L.; Perlman, S.B.; Voss, J.L.; Pine, D.S.; Leibenluft, E.; Briggs-Gowan, M.L. Clinical Implications of a Dimensional Approach: The Normal:Abnormal Spectrum of Early Irritability. J. Am. Acad Child. Adolesc. Psychiatry 2015, 54, 626–634. [Google Scholar] [CrossRef]
- Brænden, A.; Coldevin, M.; Zeiner, P.; Stubberud, J.; Melinder, A. Executive function in children with disruptive mood dysregulation disorder compared to attention-deficit/hyperactivity disorder and oppositional defiant disorder, and in children with different irritability levels. Eur. Child Adolesc. Psychiatry 2023, 1–11. [Google Scholar] [CrossRef]
- Nguyen, L.; Xu, E.; Gotts, S.; Kircanski, K.; Shaw, P.; Brotman, M.; Pine, D.; Leibenluft, E.; Linke, J. P69. Dissociating Irritability and Rigidity in Youth During Frustration. Biol. Psychiatry 2022, 91, S115. [Google Scholar] [CrossRef]
- Grabell, A.S.; Santana, A.M.; Thomsen, K.N.; Gonzalez, K.; Zhang, Z.; Bivins, Z.; Rahman, T. Prefrontal modulation of frustration-related physiology in preschool children ranging from low to severe irritability. Dev. Cogn. Neurosci. 2022, 55, 101112. [Google Scholar] [CrossRef]
- Grabell, A.S.; Li, Y.; Barker, J.W.; Wakschlag, L.S.; Huppert, T.J.; Perlman, S.B. Evidence of Non-Linear Associations between Frustration-Related Prefrontal Cortex Activation and the Normal:Abnormal Spectrum of Irritability in Young Children. J. Abnorm. Child Psychol. 2018, 46, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.B.; Luna, B.; Hein, T.C.; Huppert, T.J. fNIRS evidence of prefrontal regulation of frustration in early childhood. Neuroimage 2014, 85 Pt 1, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.B.; Jones, B.M.; Wakschlag, L.S.; Axelson, D.; Birmaher, B.; Phillips, M.L. Neural substrates of child irritability in typically developing and psychiatric populations. Dev. Cogn. Neurosci. 2015, 14, 71–80. [Google Scholar] [CrossRef]
- Abler, B.; Walter, H.; Erk, S. Neural correlates of frustration. Neuroreport 2005, 16, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Lamm, C.; Segalowitz, S.J.; Stieben, J.; Zelazo, P.D. Neurophysiological correlates of emotion regulation in children and adolescents. J. Cogn. Neurosci. 2006, 18, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Deveney, C.M.; Briggs-Gowan, M.J.; Pagliaccio, D.; Estabrook, C.R.; Zobel, E.; Burns, J.L.; Norton, E.S.; Pine, D.S.; Brotman, M.A.; Leibenluft, E.; et al. Temporally sensitive neural measures of inhibition in preschool children across a spectrum of irritability. Dev. Psychobiol. 2019, 61, 216–227. [Google Scholar] [CrossRef]
- Tseng, W.L.; Deveney, C.M.; Stoddard, J.; Kircanski, K.; Frackman, A.E.; Yi, J.Y.; Hsu, D.; Moroney, E.; Machlin, L.; Donahue, L.; et al. Brain Mechanisms of Attention Orienting Following Frustration: Associations With Irritability and Age in Youths. Am. J. Psychiatry 2019, 176, 67–76. [Google Scholar] [CrossRef]
- Hodgdon, E.A.; Yu, Q.; Kryza-Lacombe, M.; Liuzzi, M.T.; Aspe, G.I.; Menacho, V.C.; Bozzetto, L.; Dougherty, L.; Wiggins, J.L. Irritability-related neural responses to frustrative nonreward in adolescents with trauma histories: A preliminary investigation. Dev. Psychobiol. 2021, 63, e22167. [Google Scholar] [CrossRef]
- Koenigs, M.; Tranel, D. Irrational economic decision-making after ventromedial prefrontal damage: Evidence from the Ultimatum Game. J. Neurosci. 2007, 27, 951–956. [Google Scholar] [CrossRef]
- Koenigs, M.; Young, L.; Adolphs, R.; Tranel, D.; Cushman, F.; Hauser, M.; Damasio, A. Damage to the prefrontal cortex increases utilitarian moral judgements. Nature 2007, 446, 908–911. [Google Scholar] [CrossRef]
- Adleman, N.E.; Kayser, R.; Dickstein, D.; Blair, R.J.; Pine, D.; Leibenluft, E. Neural correlates of reversal learning in severe mood dysregulation and pediatric bipolar disorder. J. Am. Acad. Child Adolesc. Psychiatry 2011, 50, 1173–1185.e1172. [Google Scholar] [CrossRef] [PubMed]
- Volckaert, A.M.S.; Noël, M.-P. Training executive function in preschoolers reduce externalizing behaviors. Trends. Neurosci. Educ. 2015, 4, 37–47. [Google Scholar] [CrossRef]
- Li, Q.; Liu, P.; Yan, N.; Feng, T. Executive function training improves emotional competence for preschool children: The roles of inhibition control and working memory. Front. Psychol. 2020, 11, 347. [Google Scholar] [CrossRef] [PubMed]
- Hawes, S.W.; Perlman, S.B.; Byrd, A.L.; Raine, A.; Loeber, R.; Pardini, D.A. Chronic anger as a precursor to adult antisocial personality features: The moderating influence of cognitive control. J. Abnorm. Psychol. 2016, 125, 64. [Google Scholar] [CrossRef]
- Stroop, J.R. Studies of interference in serial verbal reactions. J. Exp. Psychol. 1935, 18, 643. [Google Scholar] [CrossRef]
- Barker, J.W.; Aarabi, A.; Huppert, T.J. Autoregressive model based algorithm for correcting motion and serially correlated errors in fNIRS. Biomed. Opt. Express 2013, 4, 1366–1379. [Google Scholar] [CrossRef]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, D280–D298. [Google Scholar] [CrossRef]
- Delpy, D.; Cope, M. Quantification in tissue near–infrared spectroscopy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1997, 352, 649–659. [Google Scholar] [CrossRef]
- Strangman, G.; Franceschini, M.A.; Boas, D.A. Factors affecting the accuracy of near-infrared spectroscopy concentration calculations for focal changes in oxygenation parameters. Neuroimage 2003, 18, 865–879. [Google Scholar] [CrossRef]
- Huppert, T.J. Commentary on the statistical properties of noise and its implication on general linear models in functional near-infrared spectroscopy. Neurophotonics 2016, 3, 010401. [Google Scholar] [CrossRef]
- Homae, F.; Watanabe, H.; Nakano, T.; Taga, G. Prosodic processing in the developing brain. Neurosci. Res. 2007, 59, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Bray, S.; Reiss, A.L. Functional near infrared spectroscopy (NIRS) signal improvement based on negative correlation between oxygenated and deoxygenated hemoglobin dynamics. Neuroimage 2010, 49, 3039–3046. [Google Scholar] [CrossRef] [PubMed]
- Wakschlag, L.S.; Henry, D.B.; Tolan, P.H.; Carter, A.S.; Burns, J.L.; Briggs-Gowan, M.J. Putting theory to the test: Modeling a multidimensional, developmentally-based approach to preschool disruptive behavior. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 593–604.e594. [Google Scholar] [CrossRef]
- Wakschlag, L.S.; Briggs-Gowan, M.J.; Choi, S.W.; Nichols, S.R.; Kestler, J.; Burns, J.L.; Carter, A.S.; Henry, D. Advancing a multidimensional, developmental spectrum approach to preschool disruptive behavior. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 82–96.e3. [Google Scholar] [CrossRef] [PubMed]
- Wakschlag, L.S.; Choi, S.W.; Carter, A.S.; Hullsiek, H.; Burns, J.; McCarthy, K.; Leibenluft, E.; Briggs-Gowan, M.J. Defining the developmental parameters of temper loss in early childhood: Implications for developmental psychopathology. J. Child Psychol. Psychiatry Allied Discip. 2012, 53, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Zerrouk, M.; Diaz, A.; Bell, M.A. Inhibitory control moderates the association between fear and attention bias to snakes in middle childhood. Cogn. Dev. 2022, 64, 101253. [Google Scholar] [CrossRef]
- Hayes, A.F. An Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Publications: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Fishburn, F.A.; Hlutkowsky, C.O.; Bemis, L.M.; Huppert, T.J.; Wakschlag, L.S.; Perlman, S.B. Irritability uniquely predicts prefrontal cortex activation during preschool inhibitory control among all temperament domains: A LASSO approach. Neuroimage 2019, 184, 68–77. [Google Scholar] [CrossRef]
- Perlman, S.B.; Huppert, T.J.; Luna, B. Functional Near-Infrared Spectroscopy Evidence for Development of Prefrontal Engagement in Working Memory in Early Through Middle Childhood. Cerebral. Cortex. 2016, 26, 2790–2799. [Google Scholar] [CrossRef]
- Fiske, A.; de Klerk, C.; Lui, K.Y.K.; Collins-Jones, L.; Hendry, A.; Greenhalgh, I.; Hall, A.; Scerif, G.; Dvergsdal, H.; Holmboe, K. The neural correlates of inhibitory control in 10-month-old infants: A functional near-infrared spectroscopy study. Neuroimage 2022, 257, 119241. [Google Scholar] [CrossRef]
- Ishii-Takahashi, A.; Takizawa, R.; Nishimura, Y.; Kawakubo, Y.; Kuwabara, H.; Matsubayashi, J.; Hamada, K.; Okuhata, S.; Yahata, N.; Igarashi, T. Prefrontal activation during inhibitory control measured by near-infrared spectroscopy for differentiating between autism spectrum disorders and attention deficit hyperactivity disorder in adults. NeuroImage Clin. 2014, 4, 53–63. [Google Scholar] [CrossRef]
- Booth, J.R.; Burman, D.D.; Meyer, J.R.; Gitelman, D.R.; Parrish, T.B.; Mesulam, M.M. Relation between brain activation and lexical performance. Hum. Brain Mapp. 2003, 19, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, M.T.; Kryza-Lacombe, M.; Christian, I.R.; Palumbo, D.E.; Amir, N.; Wiggins, J.L. Neural and behavioral correlates of inhibitory control in youths with varying levels of irritability. J. Affect Disord. 2020, 273, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, H.J. General Features of the Model. In A Model for Personality; Eysenck, H.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 1–37. [Google Scholar]
- Kumari, V.; Ffytche, D.H.; Williams, S.C.; Gray, J.A. Personality predicts brain responses to cognitive demands. J. Neurosci. 2004, 24, 10636–10641. [Google Scholar] [CrossRef] [PubMed]
- Deveney, C.M.; Connolly, M.E.; Haring, C.T.; Bones, B.L.; Reynolds, R.C.; Kim, P.; Pine, D.S.; Leibenluft, E. Neural mechanisms of frustration in chronically irritable children. Am. J. Psychiatry 2013, 170, 1186–1194. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | B | SE | t | p | 95% Confidence Interval for B | |
|---|---|---|---|---|---|---|
| Lower Limit | Upper Limit | |||||
| Constant | 0.04 | 0.10 | 0.39 | 0.690 | −0.15 | 0.23 |
| Stroop RT | −0.35 | 0.10 | −3.65 | 0.000 | −0.54 | −0.16 |
| Irritability | 0.28 | 0.10 | 2.81 | 0.006 | 0.08 | 0.48 |
| Stroop RT × Irritability | −0.22 | 0.11 | −2.11 | 0.038 | −0.43 | −0.01 |
| R2 = 17.37%, F (3,94) = 6.58, p < 0.001 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Grabell, A.S.; Perlman, S.B. Irritability Moderates the Association between Cognitive Flexibility Task Performance and Related Prefrontal Cortex Activation in Young Children. Brain Sci. 2023, 13, 882. https://doi.org/10.3390/brainsci13060882
Li Y, Grabell AS, Perlman SB. Irritability Moderates the Association between Cognitive Flexibility Task Performance and Related Prefrontal Cortex Activation in Young Children. Brain Sciences. 2023; 13(6):882. https://doi.org/10.3390/brainsci13060882
Chicago/Turabian StyleLi, Yanwei, Adam S. Grabell, and Susan B. Perlman. 2023. "Irritability Moderates the Association between Cognitive Flexibility Task Performance and Related Prefrontal Cortex Activation in Young Children" Brain Sciences 13, no. 6: 882. https://doi.org/10.3390/brainsci13060882
APA StyleLi, Y., Grabell, A. S., & Perlman, S. B. (2023). Irritability Moderates the Association between Cognitive Flexibility Task Performance and Related Prefrontal Cortex Activation in Young Children. Brain Sciences, 13(6), 882. https://doi.org/10.3390/brainsci13060882