
Alpha and Theta Oscillations Are Causally Linked to Interference Inhibition: Evidence from High-Definition Transcranial Alternating Current Stimulation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
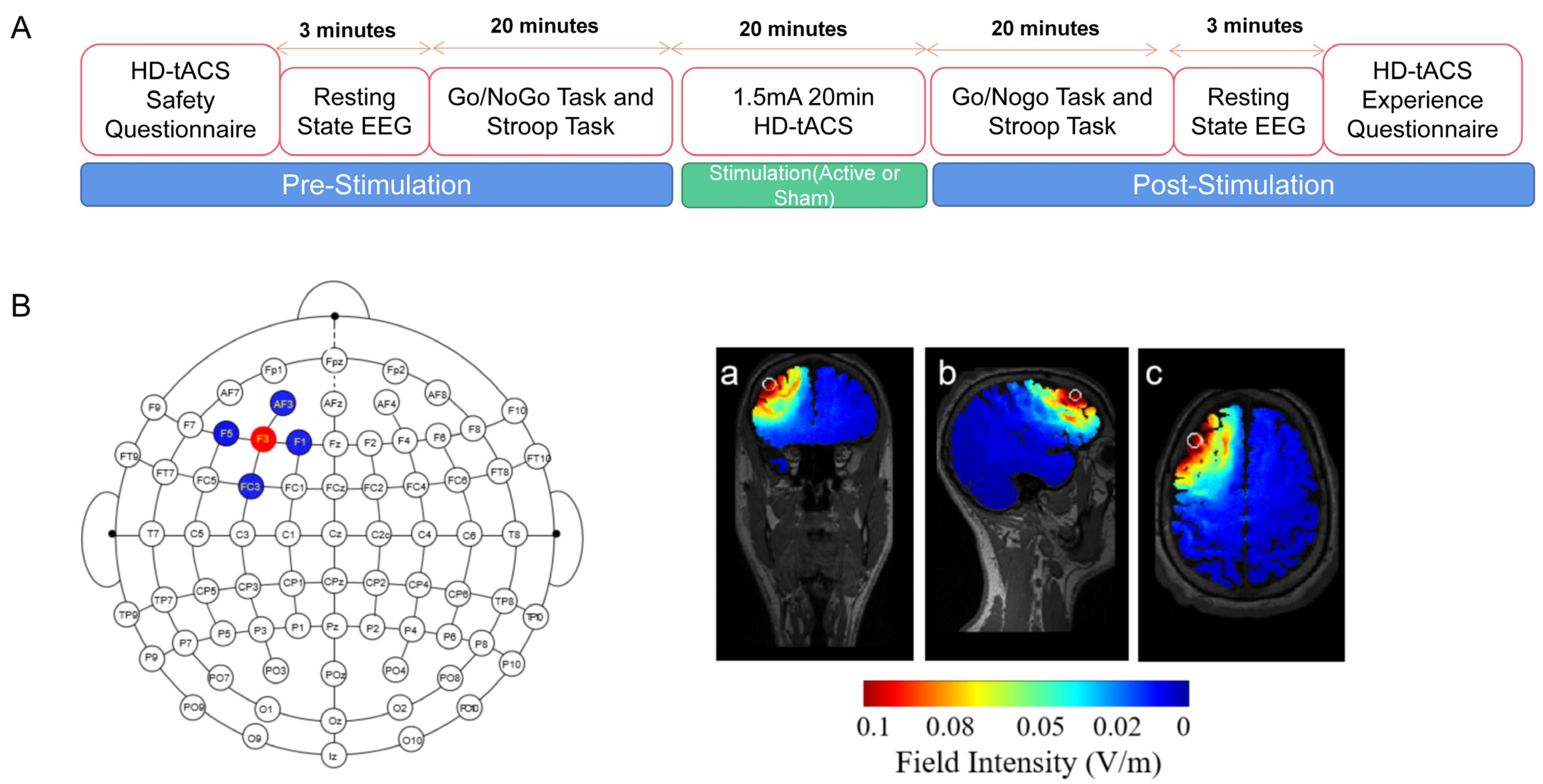
2.2. Experimental Protocol
2.3. HD-tACS
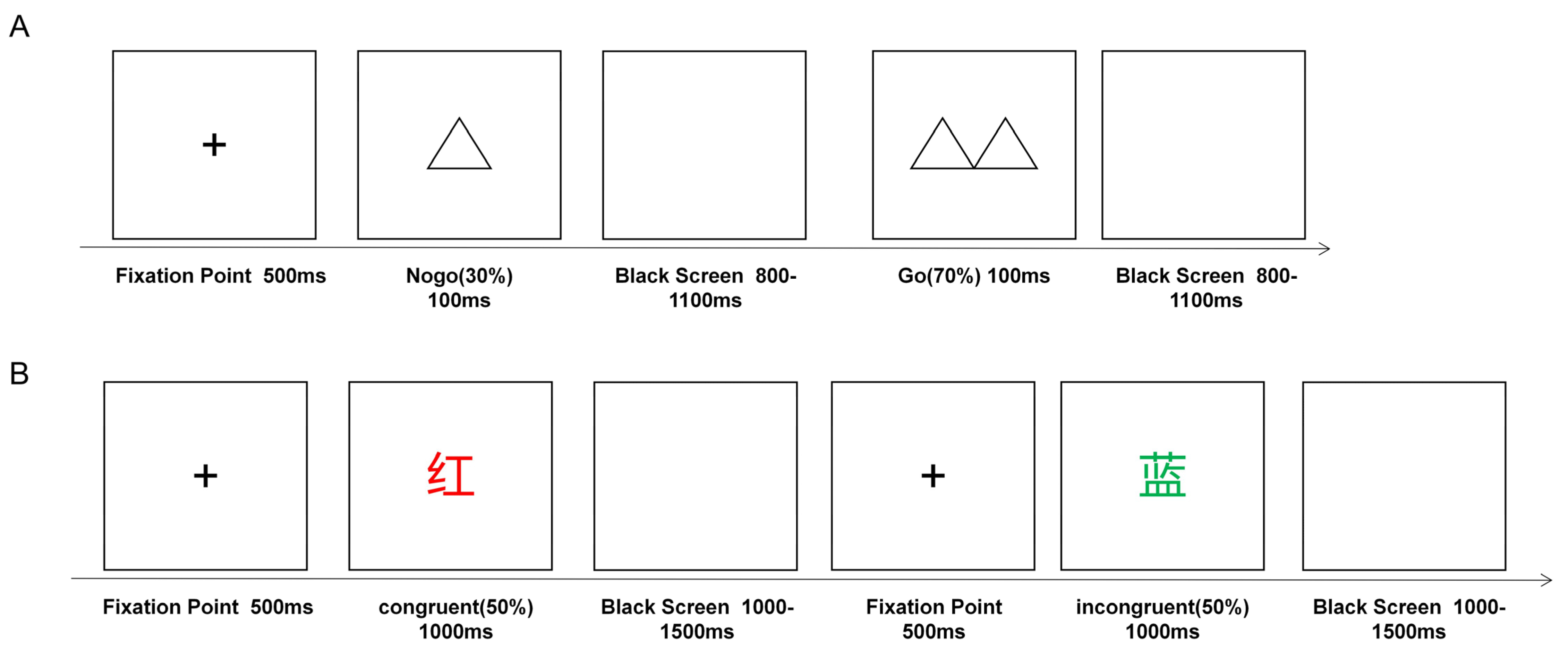
2.4. Go/NoGo Task
2.5. Color-Word Stroop Task
2.6. Resting-State EEG Recording and Preprocessing
2.7. Data Analysis: Behavioral Data
2.8. Data analysis: Resting-State EEG Data
3. Results
3.1. Demographic and Descriptive Information
3.2. Behavioral Results
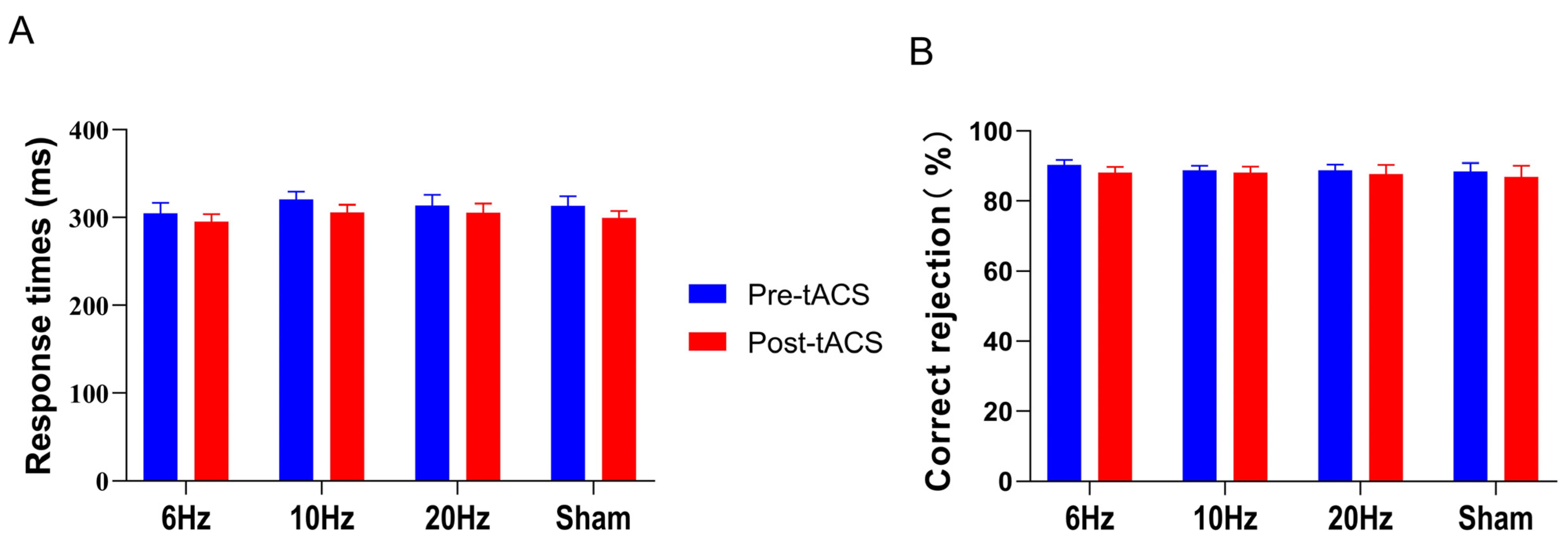
3.2.1. Effects of HD-tACS on the Go/NoGo Task
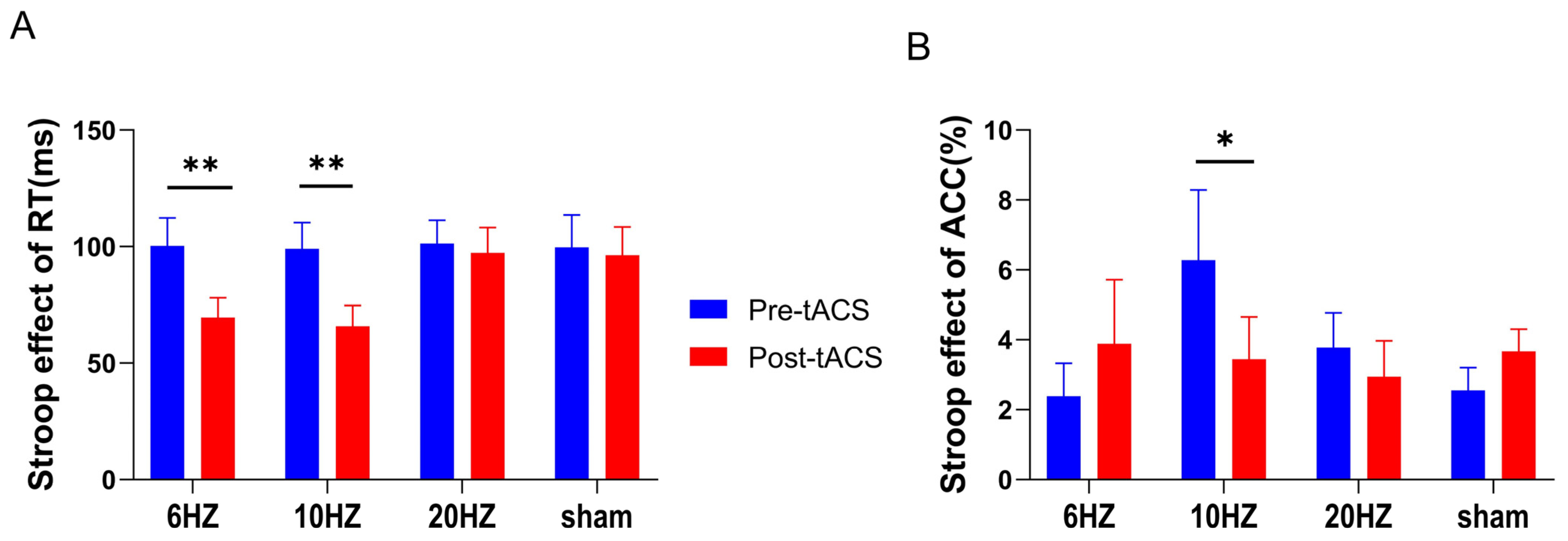
3.2.2. Effects of HD-tACS on the Color-Word Stroop Task
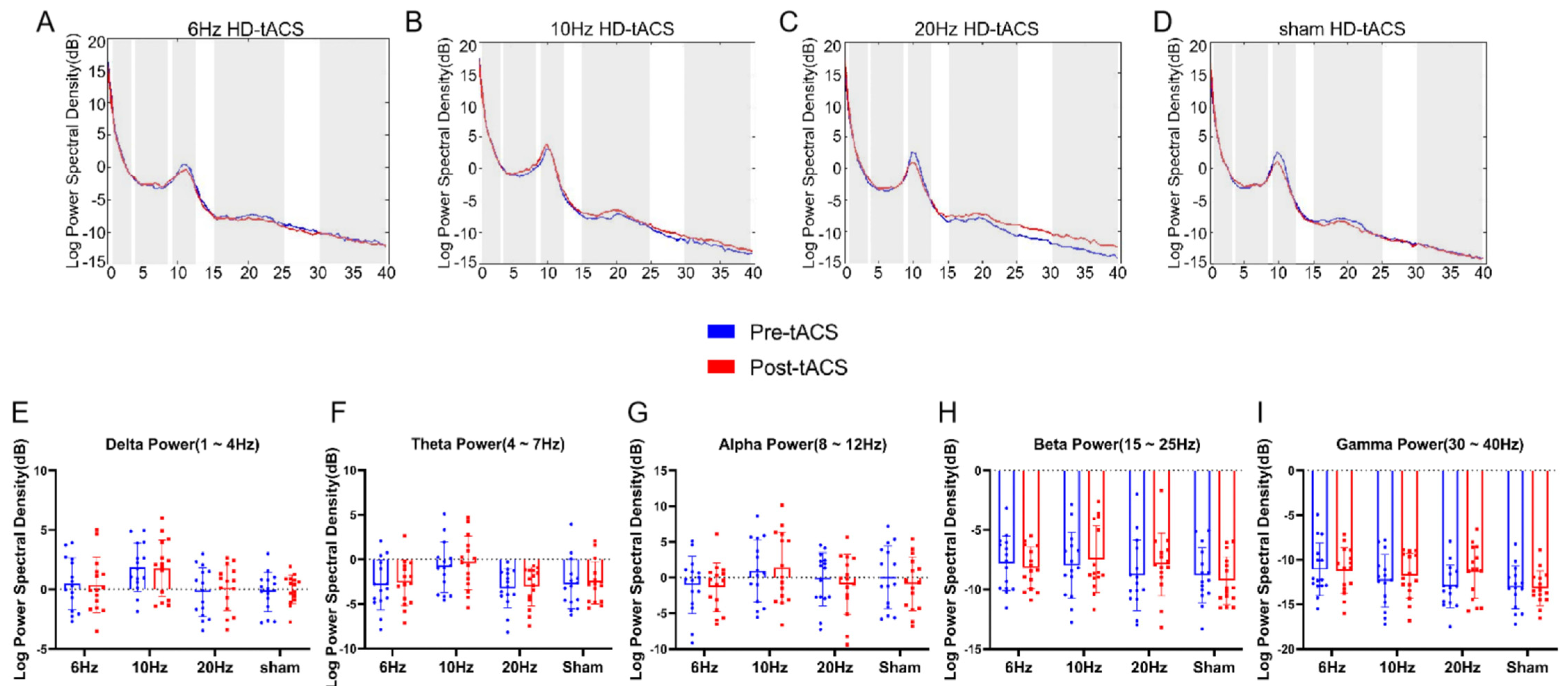
3.3. Effects of HD-tACS on EEG Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Groman, S.M.; James, A.S.; Jentsch, J.D. Poor response inhibition: At the nexus between substance abuse and attention deficit/hyperactivity disorder. Neurosci. Biobehav. Rev. 2009, 33, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.M. Contributions of Inhibitory Mechanisms to Unified Theory in Neuroscience and Psychology. Brain Cogn. 1996, 30, 127–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkley, R.A. Behavioral inhibition, sustained attention, and executive functions: Constructing a unifying theory of ADHD. Psychol. Bull. 1997, 121, 65–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, G.D.; Van Zandt, T.; Verbruggen, F.; Wagenmakers, E.J. On the ability to inhibit thought and action: General and special theories of an act of control. Psychol. Rev. 2014, 121, 66–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penolazzi, B.; Stramaccia, D.F.; Braga, M.; Mondini, S.; Galfano, G. Human memory retrieval and inhibitory control in the brain: Beyond correlational evidence. J. Neurosci. 2014, 34, 6606–6610. [Google Scholar] [CrossRef] [Green Version]
- Depue, B.E.; Burgess, G.C.; Willcutt, E.G.; Ruzic, L.; Banich, M.T. Inhibitory control of memory retrieval and motor processing associated with the right lateral prefrontal cortex: Evidence from deficits in individuals with ADHD. Neuropsychologia 2010, 48, 3909–3917. [Google Scholar] [CrossRef] [Green Version]
- Blair, C.; Razza, R.P. Relating effortful control, executive function, and false belief understanding to emerging math and literacy ability in kindergarten. Child Dev. 2007, 78, 647–663. [Google Scholar] [CrossRef]
- Bull, R.; Scerif, G. Executive Functioning as a Predictor of Children’s Mathematics Ability: Inhibition, Switching, and Working Memory. Dev. Neuropsychol. 2010, 3, 273–293. [Google Scholar] [CrossRef]
- Gilmore, C.; Attridge, N.; Clayton, S.; Cragg, L.; Johnson, S.; Marlow, N.; Simms, V.; Inglis, M.; Chambers, C. Individual differences in inhibitory control, not non-verbal number acuity, correlate with mathematics achievement. PLoS ONE 2013, 8, 67374. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhu, X.; Ziegler, A.; Shi, J. The effects of inhibitory control training for preschoolers on reasoning ability and neural activity. Sci. Rep. 2015, 5, 14200. [Google Scholar] [CrossRef] [Green Version]
- Kertzman, S.; Fluhr, A.; Vainder, M.; Abraham, W.; Dannon, P.N. The role of gender in association between inhibition capacities and risky decision making. Psychol. Res. Behav. Manag. 2018, 11, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenoy, P.; Yu, A.J. Rational decision-making in inhibitory control. Front. Hum. Neurosci. 2011, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Baler, R.D.; Volkow, N.D. Drug addiction: The neurobiology of disrupted self-control. Trends Mol. Med. 2006, 12, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K. On Conceptualizing Self-Control as More Than the Effortful Inhibition of Impulses. Personal. Soc. Psychol. Rev. 2011, 15, 352–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brydges, C.R.; Clunies-Ross, K.; Clohessy, M.; Lo, Z.L.; Nguyen, A.; Rousset, C.; Whitelaw, P.; Yeap, Y.J.; Fox, A.M. Dissociable components of cognitive control: An event-related potential (ERP) study of response inhibition and interference suppression. PLoS ONE 2012, 7, 34482. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, F.; Logan, G.D. Models of response inhibition in the stop-signal and stop-change paradigms. Neurosci. Biobehav. Rev. 2009, 33, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Bellebaum, C.; Daum, I. Cerebellar Involvement in Executive Control. Cerebellum 2007, 6, 184–192. [Google Scholar] [CrossRef]
- D’Esposito, M. From cognitive to neural models of working memory. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 761–772. [Google Scholar] [CrossRef] [Green Version]
- Fuster, J.M. Frontal lobe and cognitive development. J. Neurocytol. 2002, 31, 373–385. [Google Scholar] [CrossRef]
- Oldrati, V.; Patricelli, J.; Colombo, B.; Antonietti, A. The role of dorsolateral prefrontal cortex in inhibition mechanism: A study on cognitive reflection test and similar tasks through neuromodulation. Neuropsychologia 2016, 91, 499–508. [Google Scholar] [CrossRef]
- Brunyé, T.T.; Cantelon, J.; Holmes, A.; Taylor, H.A.; Mahoney, C.R. Mitigating Cutaneous Sensation Differences During tDCS: Comparing Sham Versus Low Intensity Control Conditions. Brain Stimul. 2014, 7, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Karuza, E.A.; Balewski, Z.Z.; Hamilton, R.H.; Medaglia, J.D.; Tardiff, N.; Thompson-Schill, S.L. Mapping the Parameter Space of tDCS and Cognitive Control via Manipulation of Current Polarity and Intensity. Front. Hum. Neurosci. 2016, 10, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friehs, M.A.; Güldenpenning, I.; Frings, C.; Weigelt, M. Electrify your Game! Anodal tDCS Increases the Resistance to Head Fakes in Basketball. J. Cogn. Enhanc. 2020, 4, 62–70. [Google Scholar] [CrossRef]
- Wiegand, A.; Sommer, A.; Nieratschker, V.; Plewnia, C. Improvement of cognitive control and stabilization of affect by prefrontal transcranial direct current stimulation (tDCS). Sci. Rep. 2019, 9, 6797. [Google Scholar] [CrossRef] [Green Version]
- Friese, M.; Binder, J.; Luechinger, R.; Boesiger, P.; Rasch, B. Suppressing emotions impairs subsequent stroop performance and reduces prefrontal brain activation. PLoS ONE 2013, 8, 60385. [Google Scholar] [CrossRef] [Green Version]
- Bechara, A. Decision making, impulse control and loss of willpower to resist drugs: A neurocognitive perspective. Nat. Neurosci. 2005, 8, 1458–1463. [Google Scholar] [CrossRef]
- Grafman, J.; Wood, J.N. Human prefrontal cortex: Processing and representational perspectives. Nature reviews. Neuroscience 2003, 4, 139–147. [Google Scholar] [CrossRef]
- Floden, D.; Stuss, D.T. Inhibitory control is slowed in patients with right superior medial frontal damage. J. Cogn. Neurosci. 2006, 18, 1843–1849. [Google Scholar] [CrossRef]
- Shimamura, A.P.; Jurica, P.J.; Mangels, J.A.; Gershberg, F.B.; Knight, R.T. Susceptibility to Memory Interference Effects following Frontal Lobe Damage: Findings from Tests of Paired-Associate Learning. J. Cogn. Neurosci. 1995, 7, 144–152. [Google Scholar] [CrossRef]
- Stramaccia, D.F.; Penolazzi, B.; Sartori, G.; Braga, M.; Mondini, S.; Galfano, G. Assessing the effects of tDCS over a delayed response inhibition task by targeting the right inferior frontal gyrus and right dorsolateral prefrontal cortex. Exp. Brain Res. 2015, 233, 2283–2290. [Google Scholar] [CrossRef]
- Qiu, J.; Li, H.; Jou, J.; Liu, J.; Luo, Y.; Feng, T.; Wu, Z.; Zhang, Q. Neural correlates of the “Aha” experiences: Evidence from an fMRI study of insight problem solving. Cortex 2010, 46, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Aziz-Zadeh, L.; Liew, S.L.; Dandekar, F. Exploring the neural correlates of visual creativity. Soc. Cogn. Affect. Neurosci. 2013, 8, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas, E.; Sejnowski, T.J. Gain modulation in the central nervous system: Where behavior, neurophysiology, and computation meet. Neuroscientist 2001, 7, 430–440. [Google Scholar] [CrossRef]
- Knyazev, G.G. Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci. Biobehav. Rev. 2007, 31, 377–395. [Google Scholar] [CrossRef] [PubMed]
- Singer, W. Neuronal synchrony: A versatile code for the definition of relations? Neuron 1999, 24, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Huster, R.J.; Enriquez-Geppert, S.; Lavallee, C.F.; Falkenstein, M.; Herrmann, C.S. Electroencephalography of response inhibition tasks: Functional networks and cognitive contributions. Int. J. Psychophysiol. 2013, 87, 217–233. [Google Scholar] [CrossRef]
- West, R.; Alain, C. Event-related neural activity associated with the Stroop task. Cogn. Brain Res. 1999, 8, 157–164. [Google Scholar] [CrossRef]
- Liotti, M.; Woldorff, M.G.; Perez, R.; Mayberg, H.S. An ERP study of the temporal course of the Stroop color-word interference effect. Neuropsychologia 2000, 38, 701–711. [Google Scholar] [CrossRef]
- West, R.; Jakubek, K.; Wymbs, N.; Perry, M.; Moore, K. Neural correlates of conflict processing. Exp. Brain Res. 2005, 167, 38–48. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Frank, M.J. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Mussel, P.; Ulrich, N.; Allen, J.J.B.; Osinsky, R.; Hewig, J. Patterns of theta oscillation reflect the neural basis of individual differences in epistemic motivation. Sci. Rep. 2016, 6, 29245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimesch, W. Alpha-band oscillations, attention, and controlled access to stored information. Trends Cogn. Sci. 2012, 16, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfurtscheller, G. Central beta rhythm during sensorimotor activities in man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 253–264. [Google Scholar] [CrossRef]
- Swann, N.; Tandon, N.; Canolty, R.; Ellmore, T.M.; McEvoy, L.K.; Dreyer, S.; DiSano, M.; Aron, A.R. Intracranial EEG reveals a time- and frequency-specific role for the right inferior frontal gyrus and primary motor cortex in stopping initiated responses. J. Neurosci. 2009, 29, 12675–12685. [Google Scholar] [CrossRef] [Green Version]
- Picazio, S.; Ponzo, V.; Koch, G. Cerebellar Control on Prefrontal-Motor Connectivity During Movement Inhibition. Cerebellum 2016, 15, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Antal, A.; Boros, K.; Poreisz, C.; Chaieb, L.; Terney, D.; Paulus, W. Comparatively weak after-effects of transcranial alternating current stimulation (tACS) on cortical excitability in humans. Brain Stimul. 2008, 1, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Sellers, K.K.; Frohlich, F. Transcranial alternating current stimulation modulates large-scale cortical network activity by network resonance. J. Neurosci. 2013, 33, 11262–11275. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Schneider, T.R.; Rach, S.; Trautmann-Lengsfeld, S.A.; Engel, A.K.; Herrmann, C.S. Entrainment of Brain Oscillations by Transcranial Alternating Current Stimulation. Curr. Biol. 2014, 24, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.; Nitsche, M.A. Effects of Transcranial Electrical Stimulation on Cognition. Clin. EEG Neurosci. 2012, 43, 192–199. [Google Scholar] [CrossRef]
- Pahor, A.; Jaušovec, N. The effects of theta transcranial alternating current stimulation (tACS) on fluid intelligence. Int. J. Psychophysiol. 2014, 93, 322–331. [Google Scholar] [CrossRef]
- Yaple, Z.; Martinez-Saito, M.; Awasthi, B.; Feurra, M.; Shestakova, A.; Klucharev, V. Transcranial Alternating Current Stimulation Modulates Risky Decision Making in a Frequency-Controlled Experiment. eNeuro 2017, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohlich, F.; McCormick, D.A. Endogenous electric fields may guide neocortical network activity. Neuron 2010, 67, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Thair, H.; Holloway, A.L.; Newport, R.; Smith, A.D. Transcranial Direct Current Stimulation (tDCS): A Beginner’s Guide for Design and Implementation. Front. Neurosci. 2017, 11, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratton, G.; Coles, M.G.; Donchin, E. A new method for off-line removal of ocular artifact. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 468–484. [Google Scholar] [CrossRef]
- Soong, A.C.; Lind, J.C.; Shaw, G.R.; Koles, Z.J. Systematic comparisons of interpolation techniques in topographic brain mapping. Electroencephalogr. Clin. Neurophysiol. 1993, 81, 185–195. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Lehr, A.; Henneberg, N.; Nigam, T.; Paulus, W.; Antal, A. Modulation of Conflict Processing by Theta-Range tACS over the Dorsolateral Prefrontal Cortex. Neural Plast. 2019, 2019, 6747049. [Google Scholar] [CrossRef]
- Klirova, M.; Vorackova, V.; Horacek, J.; Mohr, P.; Jonas, J.; Dudysova, D.U.; Kostylkova, L.; Fayette, D.; Krejcova, L.; Baumann, S.; et al. Modulating Inhibitory Control Processes Using Individualized High Definition Theta Transcranial Alternating Current Stimulation (HD theta-tACS) of the Anterior Cingulate and Medial Prefrontal Cortex. Front. Syst. Neurosci. 2021, 15, e611507. [Google Scholar] [CrossRef]
- Fusco, G.S.M.F. Research Report: Midfrontal theta transcranial alternating current stimulation modulates behavioural adjustment after error execution. Eur. J. Neurosci. 2018, 48, 3159–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Driel, J.; Sligte, I.G.; Linders, J.; Elport, D.; Cohen, M.X. Frequency Band-Specific Electrical Brain Stimulation Modulates Cognitive Control Processes. PLoS ONE 2015, 10, e0138984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglini, L.; Mena, F.; Ghiani, A.; Casco, C.; Melcher, D.; Ronconi, L. The Effect of Alpha tACS on the Temporal Resolution of Visual Perception. Front. Psychol. 2020, 11, 1765. [Google Scholar] [CrossRef] [PubMed]
- Labree, B.; Corrie, H.; Karolis, V.; Didino, D.; Cappelletti, M. Parietal alpha-based inhibitory abilities are causally linked to numerosity discrimination. Behav. Brain Res. 2020, 387, 112564. [Google Scholar] [CrossRef]
- Hopfinger, J.B.; Parsons, J.; Frohlich, F. Differential effects of 10-Hz and 40-Hz transcranial alternating current stimulation (tACS) on endogenous versus exogenous attention. Cogn. Neurosci. 2017, 8, 102–111. [Google Scholar] [CrossRef]
- Deng, Y.; Reinhart, R.M.; Choi, I.; Shinn-Cunningham, B.G. Causal links between parietal alpha activity and spatial auditory attention. Elife 2019, 8, e51184. [Google Scholar] [CrossRef] [PubMed]
- Orendacova, M.; Kvasnak, E. Effects of Transcranial Alternating Current Stimulation and Neurofeedback on Alpha (EEG) Dynamics: A Review. Front. Hum. Neurosci. 2021, 15, 628229. [Google Scholar] [CrossRef] [PubMed]
- Castellano, M.; Ibanez-Soria, D.; Kroupi, E.; Acedo, J.; Campolo, M.; Soria-Frisch, A.; Valls-Sole, J.; Verma, A.; Ruffini, G. Intermittent tACS during a visual task impacts neural oscillations and LZW complexity. Exp. Brain Res. 2020, 238, 1411–1422. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Herrmann, C.S.; Engel, A.K.; Schneider, T.R. Different coupling modes mediate cortical cross-frequency interactions. Neuroimage 2016, 140, 76–82. [Google Scholar] [CrossRef]
- Clayton, M.S.; Yeung, N.; Cohen Kadosh, R. The roles of cortical oscillations in sustained attention. Trends Cogn. Sci. 2015, 19, 188–195. [Google Scholar] [CrossRef]
- Simmonds, D.J.; Pekar, J.J.; Mostofsky, S.H. Meta-analysis of Go/No-go tasks demonstrating that fMRI activation associated with response inhibition is task-dependent. Neuropsychologia 2008, 46, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Badzakova-Trajkov, G.; Barnett, K.J.; Waldie, K.E.; Kirk, I.J. An ERP investigation of the Stroop task: The role of the cingulate in attentional allocation and conflict resolution. Brain Res. 2009, 1253, 139–148. [Google Scholar] [CrossRef]
- Daughters, S.B.; Yi, J.Y.; Phillips, R.D.; Carelli, R.M.; Frohlich, F. Alpha-tACS effect on inhibitory control and feasibility of administration in community outpatient substance use treatment. Drug Alcohol Depend. 2020, 213, 108132. [Google Scholar] [CrossRef] [PubMed]
- Neuling, T.; Rach, S.; Herrmann, C.S. Orchestrating neuronal networks: Sustained after-effects of transcranial alternating current stimulation depend upon brain states. Front. Hum. Neurosci. 2013, 7, 161. [Google Scholar] [CrossRef] [Green Version]
- Wischnewski, M.; Engelhardt, M.; Salehinejad, M.A.; Schutter, D.J.L.G.; Kuo, M.F.; Nitsche, M.A. NMDA Receptor-Mediated Motor Cortex Plasticity After 20 Hz Transcranial Alternating Current Stimulation. Cereb. Cortex 2019, 29, 2924–2931. [Google Scholar] [CrossRef] [PubMed]
- Ruhnau, P.; Neuling, T.; Fuscá, M.; Herrmann, C.S.; Demarchi, G.; Weisz, N. Eyes wide shut: Transcranial alternating current stimulation drives alpha rhythm in a state dependent manner. Sci. Rep. 2016, 6, 27138. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Hernández, S.P.; Rahman, M.S.; Voigt, K.; Malvaso, A. Inhibitory Control Development: A Network Neuroscience Perspective. Front. Psychol. 2022, 13, 651547. [Google Scholar] [CrossRef] [PubMed]
- Cragg, L. The development of stimulus and response interference control in midchildhood. Dev. Psychol. 2016, 52, 242. [Google Scholar] [CrossRef]
- Luna, B. Developmental changes in cognitive control through adolescence. Adv. Child Dev. Behav. 2009, 37, 233–278. [Google Scholar] [CrossRef]
- Kang, W.; Wang, J.; Malvaso, A. Inhibitory Control in Aging: The Compensation-Related Utilization of Neural Circuits Hypothesis. Front. Aging Neurosci. 2021, 13, 771885. [Google Scholar] [CrossRef]
- Guerra, A.; Asci, F.; Zampogna, A.; D’Onofrio, V.; Berardelli, A.; Suppa, A. The effect of gamma oscillations in boosting primary motor cortex plasticity is greater in young than older adults. Clin. Neurophysiol. 2021, 132, 1358–1366. [Google Scholar] [CrossRef]
- Antonenko, D.; Grittner, U.; Saturnino, G.; Nierhaus, T.; Thielscher, A.; Flöel, A. Inter-individual and age-dependent variability in simulated electric fields induced by conventional transcranial electrical stimulation. Neuroimage 2021, 224, 117413. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Wu, D.; Sun, K.; Chen, X.; Wang, Y.; He, Y.; Xiao, W. Alpha and Theta Oscillations Are Causally Linked to Interference Inhibition: Evidence from High-Definition Transcranial Alternating Current Stimulation. Brain Sci. 2023, 13, 1026. https://doi.org/10.3390/brainsci13071026
Zhu Y, Wu D, Sun K, Chen X, Wang Y, He Y, Xiao W. Alpha and Theta Oscillations Are Causally Linked to Interference Inhibition: Evidence from High-Definition Transcranial Alternating Current Stimulation. Brain Sciences. 2023; 13(7):1026. https://doi.org/10.3390/brainsci13071026
Chicago/Turabian StyleZhu, Yan, Di Wu, Kewei Sun, Xianglong Chen, Yifan Wang, Yang He, and Wei Xiao. 2023. "Alpha and Theta Oscillations Are Causally Linked to Interference Inhibition: Evidence from High-Definition Transcranial Alternating Current Stimulation" Brain Sciences 13, no. 7: 1026. https://doi.org/10.3390/brainsci13071026
APA StyleZhu, Y., Wu, D., Sun, K., Chen, X., Wang, Y., He, Y., & Xiao, W. (2023). Alpha and Theta Oscillations Are Causally Linked to Interference Inhibition: Evidence from High-Definition Transcranial Alternating Current Stimulation. Brain Sciences, 13(7), 1026. https://doi.org/10.3390/brainsci13071026