Age-Related Deficits in Memory Encoding and Retrieval in Word List Free Recall

{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.3. Procedure
2.4. Semantic Clustering Scores
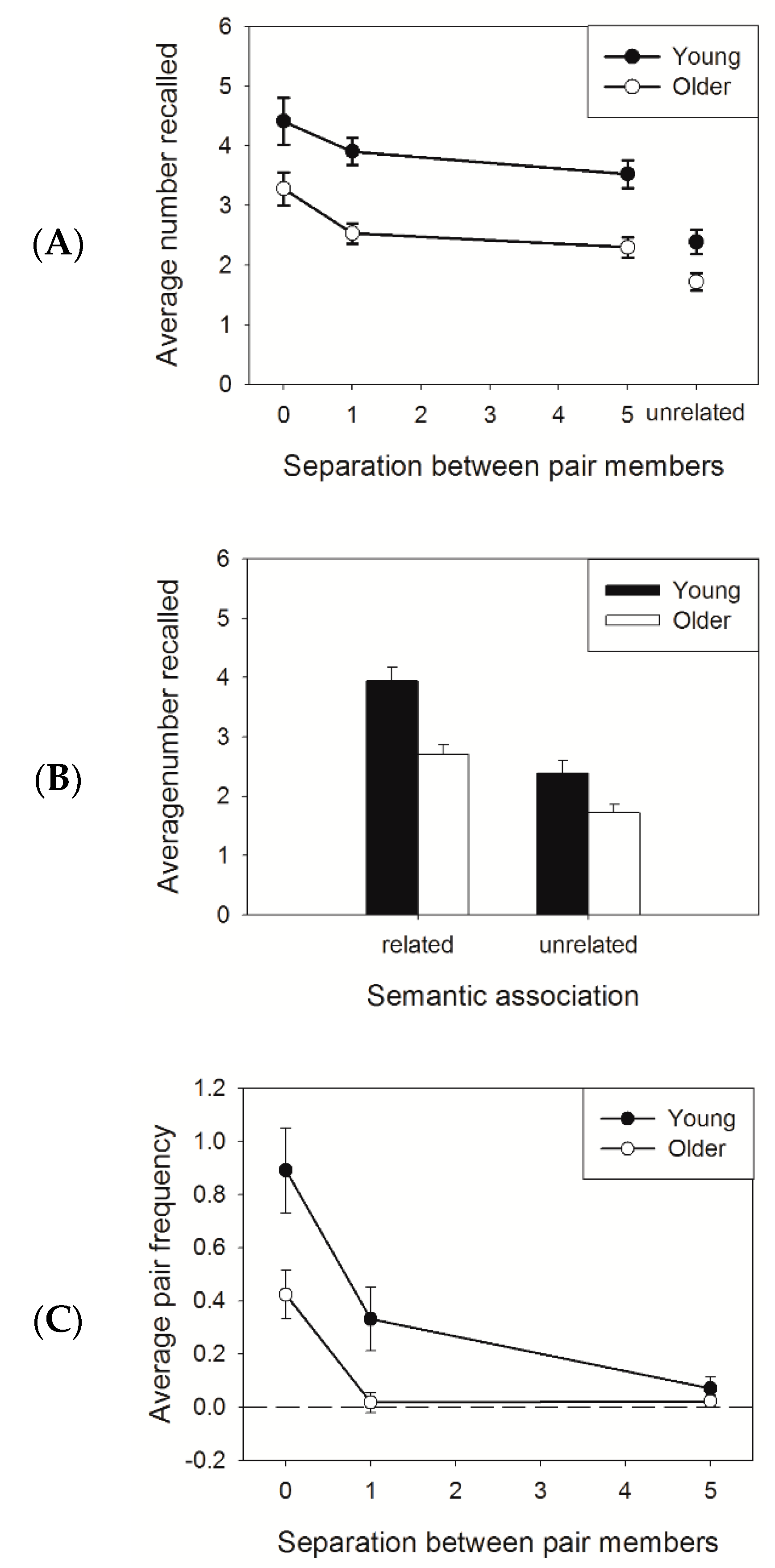
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Craik, F.I.M.; McDowd, J.M. Age differences in recall and recognition. J. Exp. Psychol. Learn. Mem. Cogn. 1987, 13, 474–479. [Google Scholar] [CrossRef]
- Kahana, M.J.; Howard, M.W.; Zaromb, F.; Wingfield, A. Age dissociates recency and lag recency effects in free recall. J. Exp. Psychol. Learn. Mem. Cogn. 2002, 3, 530–540. [Google Scholar] [CrossRef]
- Howard, M.W.; Kahana, M.J. A distributed representation of temporal context. J. Math. Psychol. 2002, 46, 269–299. [Google Scholar] [CrossRef]
- Wingfield, A.; Lindfield, K.C.; Kahana, M.J. Adult age differences in the temporal characteristics of category free recall. Psychol. Aging 1998, 13, 256–266. [Google Scholar] [CrossRef]
- Craik, F.I.M.; Jennings, J.M. Human memory. In Handbook of Aging and Cognition, 1st ed.; Craik, F.I.M., Salthouse, T.A., Eds.; Erlbaum: Hillsdale, NJ, USA, 1992; pp. 51–110. [Google Scholar]
- Rybarczyk, B.D.; Hart, R.P.; Harkins, S.W. Age and forgetting rate with pictorial stimuli. Psychol. Aging 1987, 2, 404–406. [Google Scholar] [CrossRef] [PubMed]
- West, R. An application of prefrontal cortex function theory to cognitive aging. Psychol. Bull. 1996, 120, 272–292. [Google Scholar] [CrossRef] [PubMed]
- Parkin, A.J.; Walter, B.M. Recollective experience, normal aging, and frontal dysfunction. Psychol. Aging 1992, 7, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Haug, H.; Eggers, R. Morphometry of the human cortex cerebri and human striatum during aging. Neurobiol. Aging 1991, 12, 336–338. [Google Scholar] [CrossRef]
- Cabeza, R.; Grady, C.L.; Nyberg, L.; McIntosh, A.R.; Tulving, E.; Kapur, S.; Jennings, J.M.; Houle, S.; Craik, F.I.M. Age-related differences in neural activity during memory encoding and retrieval: A positron emission tomography study. J. Neurosci. 1997, 17, 391–400. [Google Scholar] [CrossRef]
- Stebbins, G.T.; Carrillo, M.C.; Dorman, J.; Dirksen, C.; Desond, J.E.; Turner, D.A.; Bennett, D.A.; Wilson, R.S.; Glover, G.; Gabrieli, J.D. Aging effects on memory encoding in the frontal lobes. Psychol. Aging 2002, 17, 44–55. [Google Scholar] [CrossRef]
- Baldo, J.V.; Delis, D.; Kramer, J.; Shimamura, A.P. Memory performance on the California Verbal Learning Test II: Finding from patients with local frontal lesions. J. Int. Neuropsychol. Soc. 2002, 8, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Janowsky, J.S.; Shimamura, A.P.; Squire, L.R. Source memory impairment in patients with frontal lobe lesions. Neuropsychologia 1989, 27, 1043–1056. [Google Scholar] [CrossRef]
- Simons, J.S.; Verfaellie, M.; Galton, C.J.; Miller, B.L.; Hodges, J.R.; Graham, K.S. Recollection-based memory in frontotemporal dementia: Implications for theories of long-term memory. Brain 2002, 125, 2523–2536. [Google Scholar] [CrossRef]
- Moscovitch, M. Amnesia. In The International Encyclopedia of Social and Behavioural Sciences; Smesler, N.B., Baltes, O.B., Eds.; Pergamon/Elsevier Science: Oxford, UK, 2004; pp. 1–26. [Google Scholar]
- Moscovitch, M.; Westmacott, R.; Gilboa, A.; Addis, D.R.; Rosenbaum, R.S.; Viskontas, I.; Priselac, S.; Svoboda, E.; Ziegler, M.; Black, S.; et al. Hippocampal complex contribution to retention and retrieval of recent and remote episodic and semantic memories: Evidence from behavioral and neuroimaging studies of healthy and brain-damaged people. In Dynamic Cognitive Processes; Ohta, N., MacLeod, C.M., Uttl, B., Eds.; Springer: Tokyo, Japan, 2005; pp. 333–380. [Google Scholar]
- Nadel, L.; Moscovitch, M. Memory consolidation, retrograde amnesia and the hippocampal complex. Curr. Opin. Neurobiol. 1997, 7, 217–227. [Google Scholar] [CrossRef]
- Nadel, L.; Samsonovich, A.; Ryan, L.; Moscovitch, M. Multiple trace theory of human memory: Computational, neuroimaging, and neuropsychological results. Hippocampus 2000, 10, 352–368. [Google Scholar] [CrossRef]
- Simons, J.S.; Spiers, H.J. Prefrontal and medial temporal lobe interactions in long-term memory. Nat. Rev. Neurosci. 2003, 4, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Stricker, J.L.; Brown, G.G.; Wixted, J.T.; Baldo, J.V.; Delis, D. New semantic and serial clustering indices for the California Verbal Learning Test 2: Background, rationale, and formulae. J. Int. Neuropsychol. Soc. 2002, 8, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Taconnat, L.; Raz, N.; Tocze, C.; Bouazzaoui, B.; Sauzeon, H.; Fay, S.; Isingrini, M. Ageing and organisation strategies in free recall: the role of cognitive flexibility. Eur. J. Cogn. Psychol. 2009, 21, 347–365. [Google Scholar] [CrossRef]
- Backman, L.; Larsson, M. Recall of organizable words and objects in adulthood: Influences of instructions, retention interval, and retrieval cues. J. Gerontol. 1992, 47, P273–P278. [Google Scholar] [CrossRef] [PubMed]
- Verhaeghen, P.; Marcoen, A. More or less the same? A memorability analysis on episodic memory tasks in young and older adults. J. Gerontol. 1993, 48, P172–P178. [Google Scholar] [CrossRef] [PubMed]
- Elhalal, A.; Davelaar, E.J.; Usher, M. The role of the frontal cortex in memory: An investigation of the Von Restorff effect. Front. Hum. Neurosci. 2014, 8, 410. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.A.; Evans, J.D.; Miller, S.W.; Heaton, R.K. Demographically corrected norms for the California Verbal Learning Test. J. Clin. Exp. Neuropsychol. 2000, 22, 80–94. [Google Scholar] [CrossRef]
- Glanzer, M. Distance between related words in free recall: Trace of the STS. J. Verbal Learn. Verbal Behav. 1969, 8, 105–111. [Google Scholar] [CrossRef]
- Davelaar, E.J.; Haarmann, H.J.; Goshen-Gottstein, Y.; Usher, M. Semantic similarity dissociates short from long-term recency effects: Testing a neurocomputational model of list memory. Mem. Cogn. 2006, 34, 323–334. [Google Scholar] [CrossRef]
- Haarmann, H.J.; Usher, M. Maintenance of semantic information in capacity-limited item short-term memory. Psychol. Bull. Rev. 2001, 8, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Cowan, N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behav. Brain Sci. 2001, 24, 87–185. [Google Scholar] [CrossRef]
- Raaijmakers, J.G.W.; Shiffrin, R.M. Search of associative memory. Psychol. Rev. 1981, 88, 93–134. [Google Scholar] [CrossRef]
- Nelson, D.L.; McEvoy, C.L.; Schreiber, T.A. The University of South Florida word association, rhyme, and word fragment norms. Available online: http://www.usf.edu/FreeAssociation/ (accessed on 30 January 2008).
- Bousfield, A.K.; Bousfield, W.A. Measurement of clustering and of sequential constancies in repeated free recall. Psychol. Rep. 1966, 19, 935–942. [Google Scholar] [CrossRef]
- Delis, D.C.; Kramer, J.H.; Kaplan, E.; Ober, B. California Verbal Learning Test: Adult Version Manual, 1st ed.; The Psychological Corporation: San Antonio, TX, USA, 1987. [Google Scholar]
- Sternberg, R.J.; Tulving, E. The measurement of subjective organization in free recall. Psychol. Bull. 1977, 84, 539–556. [Google Scholar] [CrossRef]
- Naveh-Benjamin, M. Adult age differences in memory performance: Tests of an associative deficit hypothesis. J. Exp. Psychol. Learn. Mem. Cogn. 2000, 26, 1170–1187. [Google Scholar] [CrossRef]
- Haarmann, H.J.; Ashling, G.E.; Davelaar, E.J.; Usher, M. Age-related declines in context maintenance and semantic short-term memory. Q. J. Exp. Psychol. 2005, 58, 34–53. [Google Scholar] [CrossRef] [PubMed]
- Hirst, W.; Volpe, B.T. Memory strategies with brain damage. Brain Cogn. 1988, 8, 379–408. [Google Scholar] [CrossRef]
- Tulving, E.; Kapur, S.; Craik, F.I.M.; Moscovitch, M.; Houle, S. Hemispheric encoding/retrieval asymmetry in episodic memory: Positron emission tomography findings. Proc. Natl. Acad. Sci. 1994, 91, 2016–2020. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Lim, J. A computational model of prefrontal control in free recall: Strategic memory use in the California Verbal Learning Task. J. Cogn. Neurosci. 2003, 15, 821–832. [Google Scholar] [CrossRef]
- Moscovitch, M.; Winocur, G. Frontal lobes, memory and aging. Ann. N. Y. Acad. Sci. 1995, 769, 119–150. [Google Scholar] [CrossRef]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadar, D.; Usher, M.; Davelaar, E.J. Age-Related Deficits in Memory Encoding and Retrieval in Word List Free Recall. Brain Sci. 2018, 8, 211. https://doi.org/10.3390/brainsci8120211
Cadar D, Usher M, Davelaar EJ. Age-Related Deficits in Memory Encoding and Retrieval in Word List Free Recall. Brain Sciences. 2018; 8(12):211. https://doi.org/10.3390/brainsci8120211
Chicago/Turabian StyleCadar, Dorina, Marius Usher, and Eddy J. Davelaar. 2018. "Age-Related Deficits in Memory Encoding and Retrieval in Word List Free Recall" Brain Sciences 8, no. 12: 211. https://doi.org/10.3390/brainsci8120211
APA StyleCadar, D., Usher, M., & Davelaar, E. J. (2018). Age-Related Deficits in Memory Encoding and Retrieval in Word List Free Recall. Brain Sciences, 8(12), 211. https://doi.org/10.3390/brainsci8120211