The Sound of Words Evokes Affective Brain Responses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Stimuli
2.2. Participants
2.3. Procedure
2.4. fMRI data Acquisition
2.5. Post-Scan Tests
2.5.1. Unannounced Recognition Test
2.5.2. Ratings
2.6. fMRI Preprocessing
2.7. fMRI Analysis
3. Results
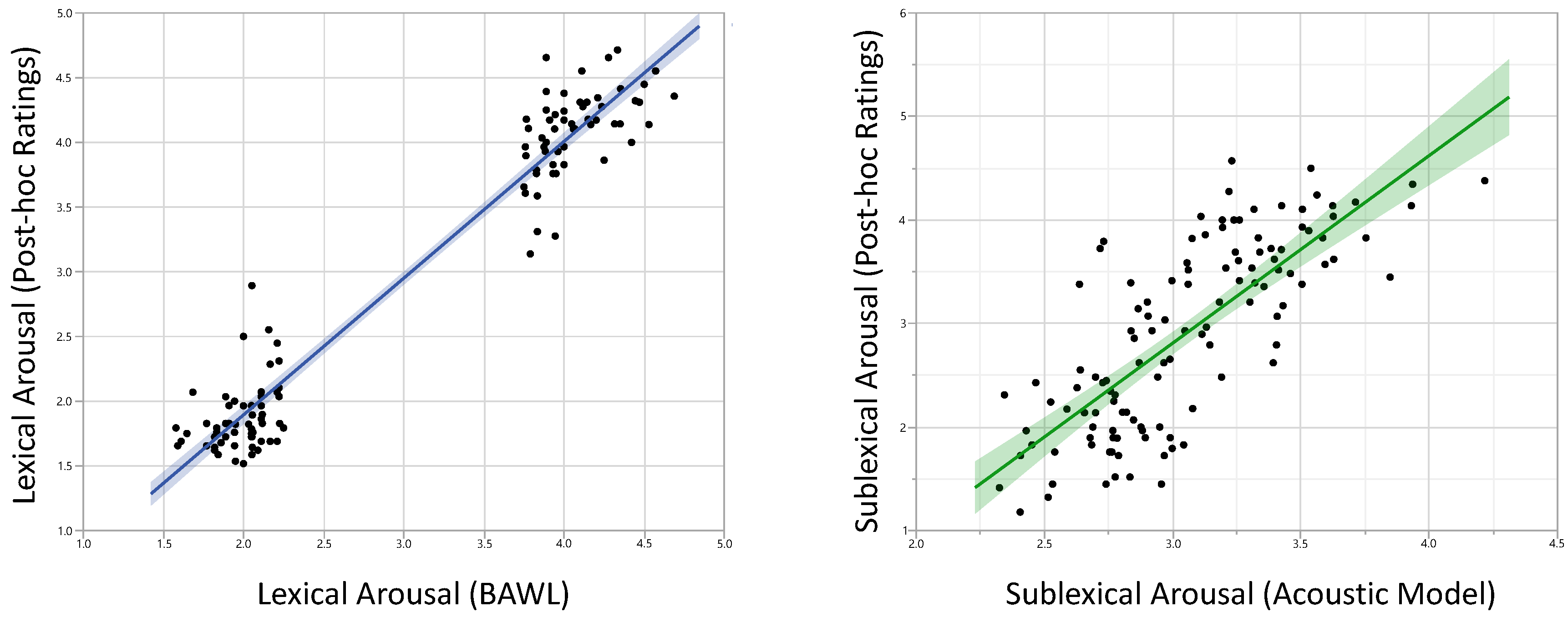
3.1.Behavioral Results
3.1.1. Recognition Test
3.1.2. Ratings
3.2. Neuroimaging Results
3.2.1. Main Effect of all Words Compared to SCN
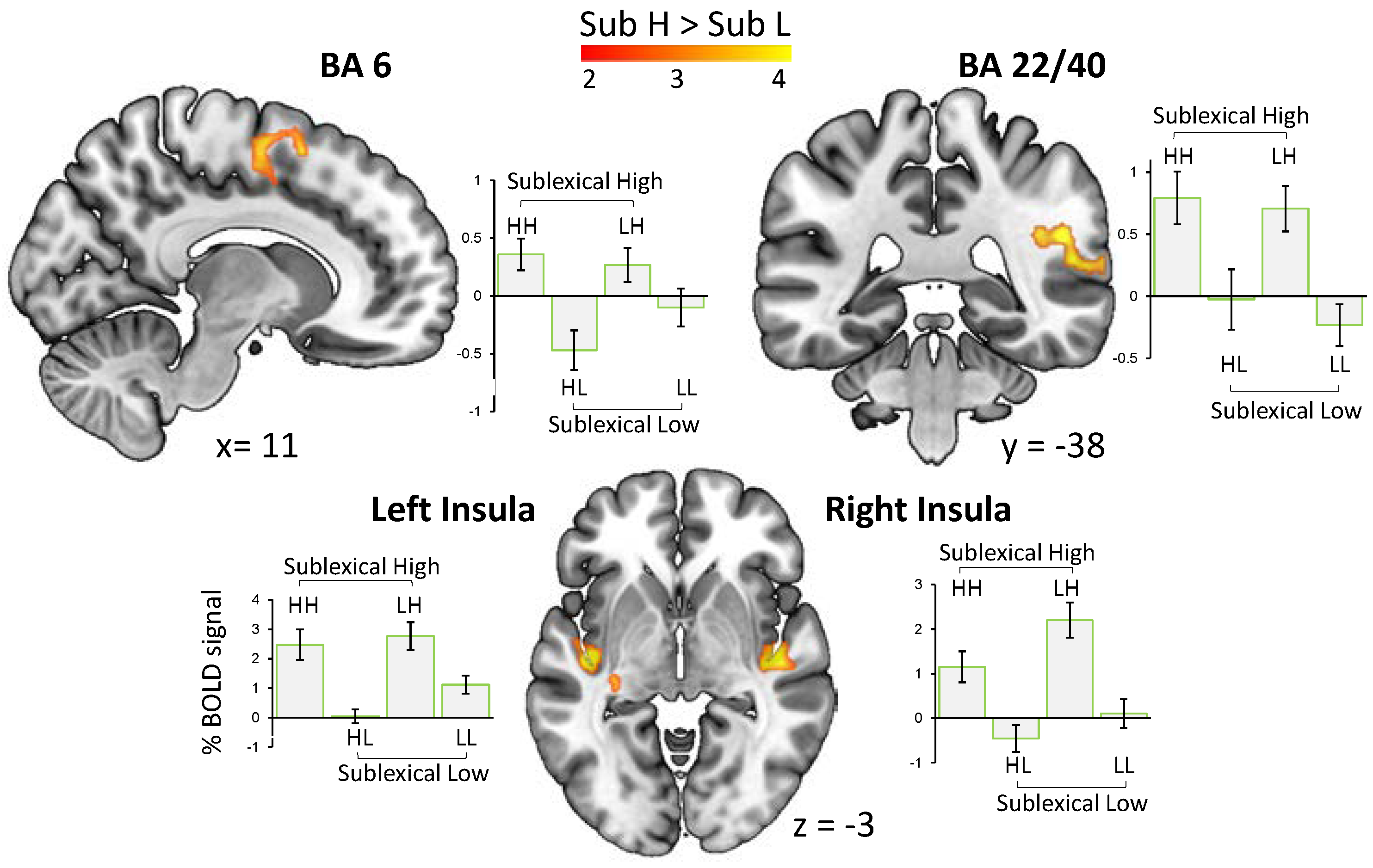
3.2.2. Main Effect of the Category Lexical Arousal
3.3.3. Main Effect of the Category Sublexical Arousal
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Saussure, F. Course in General Linguistics; Columbia University Press: New York, NY, USA, 2011. [Google Scholar]
- Aryani, A.; Jacobs, A.M.; Conrad, M. Extracting salient sublexical units from written texts: “Emophon,” a corpus-based approach to phonological iconicity. Front. Psychol. 2013, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Aryani, A.; Kraxenberger, M.; Ullrich, S.; Jacobs, A.M.; Conrad, M. Measuring the Basic Affective Tone of Poems via Phonological Saliency and Iconicity. Psychol. Aesthet. Creat. Arts 2016, 10, 191–204. [Google Scholar] [CrossRef]
- Jacobs, A.M. Neurocognitive poetics: Methods and models for investigating the neuronal and cognitive-affective bases of literature reception. Front. Hum. Neurosci. 2015, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Jakobson, R. Quest for the Essence of Language. Diogenes 1965, 13, 21–37. [Google Scholar] [CrossRef]
- Schrott, R.; Jacobs, A.M. Gehirn und Gedicht: Wie wir Unsere Wirklichkeiten Konstruieren; Hanser: Munich, Germany, 2011. [Google Scholar]
- Jacobs, A.M.; Võ, M.L.H.; Briesemeister, B.B.; Conrad, M.; Hofmann, M.J.; Kuchinke, L.; Lüdtke, J.; Braun, M. 10 years of BAWLing into affective and aesthetic processes in reading: What are the echoes? Front. Psychol. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Aryani, A.; Conrad, M.; Schmidtcke, D.; Jacobs, A.M. Why “piss” is ruder than “pee”? The role of sound in affective meaning making. PsyArXiv 2018. [Google Scholar] [CrossRef]
- Jacobs, A.M. Quantifying the Beauty of Words: A Neurocognitive Poetics Perspective. Front. Hum. Neurosci. 2017, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.; Kotz, S.A.; Schmidtke, D.S.; Aryani, A.; Conrad, M. Phonological iconicity electrifies: An ERP study on affective sound-to-meaning correspondences in German. Front. Psychol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wundt, W.M. Völkerpsychologie; Eine Untersuchung der Entwicklungsgesetze von Sprache, Mythus und Sitte: Bd. Die Kunst; Wilhelm Engelmann: Lemgo, Germany, 1908; Volume 3. [Google Scholar]
- Osgood, C.E. The nature and measurement of meaning. Psychol. Bull. 1952, 49, 197. [Google Scholar] [CrossRef] [PubMed]
- Weninger, F.; Eyben, F.; Schuller, B.W.; Mortillaro, M.; Scherer, K.R. On the acoustics of emotion in audio: What speech, music, and sound have in common. Front. Psychol. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frühholz, S.; Trost, W.; Kotz, S.A. The sound of emotions-Towards a unifying neural network perspective of affective sound processing. Neurosci. Biobehav. Rev. 2016, 68, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Juslin, P.N.; Laukka, P. Communication of Emotions in Vocal Expression and Music Performance: Different Channels, Same Code? Psychol. Bull. 2003, 129, 770–814. [Google Scholar] [CrossRef] [PubMed]
- Võ, M.L.H.; Conrad, M.; Kuchinke, L.; Urton, K.; Hofmann, M.J.; Jacobs, A.M. The Berlin Affective Word List Reloaded (BAWL-R). Behav. Res. Methods 2009, 41, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified segmentation. Neuroimage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J. A fast diffeomorphic image registration algorithm. Neuroimage 2007, 38, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.P.; Friston, K.J. Generalisability, random E ects & population inference. Neuroimage 1998, 7, S754. [Google Scholar]
- Lieberman, M.D.; Cunningham, W.A. Type I and Type II error concerns in fMRI research: Re-balancing the scale. Soc. Cogn. Affect. Neurosci. 2009, 4, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Kuchinke, L.; Jacobs, A.M.; Grubich, C.; Võ, M.L.H.; Conrad, M.; Herrmann, M. Incidental effects of emotional valence in single word processing: An fMRI study. Neuroimage 2005, 28, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, M.; Hofmann, M.J.; Briesemeister, B.B.; Jacobs, A.M. Mixing positive and negative valence: Affective-semantic integration of bivalent words. Sci. Rep. 2016, 6, 30718. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.A.; Critchley, H.D.; Rotshtein, P.; Dolan, R.J. Neural correlates of processing valence and arousal in affective words. Cereb. Cortex 2007, 17, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V.; Lakoff, G. The brain’s concepts: The role of the sensory-motor system in conceptual knowledge. Cogn. Neuropsychol. 2005, 22, 455–479. [Google Scholar] [CrossRef] [PubMed]
- Vigliocco, G.; Meteyard, L.; Andrews, M.; Kousta, S. Toward a theory of semantic representation. Lang. Cogn. 2009, 1, 219–247. [Google Scholar] [CrossRef]
- Glenberg, A.M. Embodiment as a unifying perspective for psychology. Wiley Interdiscip. Rev. Cogn. Sci. 2010, 1, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.M.; Hofmann, M.J.; Kinder, A. On elementary affective decisions: To like or not to like, that is the question. Front. Psychol. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ethofer, T.; Anders, S.; Wiethoff, S.; Erb, M.; Herbert, C.; Saur, R.; Grodd, W.; Wildgruber, D. Effects of prosodic emotional intensity on activation of associative auditory cortex. Neuroreport 2006, 17, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Wiethoff, S.; Wildgruber, D.; Kreifelts, B.; Becker, H.; Herbert, C.; Grodd, W.; Ethofer, T. Cerebral processing of emotional prosody-influence of acoustic parameters and arousal. Neuroimage 2008, 39, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Belin, P.; McAdams, S.; Smith, B.; Savel, S.; Thivard, L.; Samson, S.; Samson, Y. The functional anatomy of sound intensity discrimination. J. Neurosci. 1998, 18, 6388–6394. [Google Scholar] [CrossRef] [PubMed]
- Wildgruber, D.; Ackermann, H.; Kreifelts, B.; Ethofer, T. Cerebral processing of linguistic and emotional prosody: FMRI studies. Prog. Brain Res. 2006, 4, 249–268. [Google Scholar]
- Frühholz, S.; Trost, W.; Grandjean, D. The role of the medial temporal limbic system in processing emotions in voice and music. Prog. Neurobiol. 2014, 123, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Trost, W.; Ethofer, T.; Zentner, M.; Vuilleumier, P. Mapping aesthetic musical emotions in the brain. Cereb. Cortex 2012, 22, 2769–2783. [Google Scholar] [CrossRef] [PubMed]
- Mirz, F.; Gjedde, A.; Sødkilde-Jrgensen, H.; Pedersen, C.B. Functional brain imaging of tinnitus-like perception induced by aversive auditory stimuli. Neuroreport 2000, 11, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Leitman, D.I.; Wolf, D.H.; Ragland, J.D.; Laukka, P.; Loughead, J.; Valdez, J.N.; Javitt, D.C.; Turetsky, B.; Gur, R. “It’s not what you say, but how you say it”: A reciprocal temporo-frontal network for affective prosody. Front. Hum. Neurosci. 2010, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Frühholz, S.; Grandjean, D. Towards a fronto-temporal neural network for the decoding of angry vocal expressions. Neuroimage 2012, 62, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Löfberg, O.; Julkunen, P.; Pääkkönen, A.; Karhu, J. The auditory-evoked arousal modulates motor cortex excitability. Neuroscience 2014, 274, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Zald, D.H.; Pardo, J.V. The neural correlates of aversive auditory stimulation. Neuroimage 2002, 16, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Janata, P.; Tomic, S.T.; Haberman, J.M. Sensorimotor coupling in music and the psychology of the groove. J. Exp. Psychol. Gen. 2012, 141, 54–75. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | Word Category | Inferential Statistics | |||||||
|---|---|---|---|---|---|---|---|---|---|
| HH | HL | LH | LL | ||||||
| M | SD | M | SD | M | SD | M | SD | ||
| Lexical Arousal | 4.07 | 0.24 | 4.04 | 0.22 | 1.99 | 0.16 | 1.99 | 0.18 | F(3,116) = 983, p < 0.0001 |
| Lexical Valence | −1.83 | 0.52 | −1.83 | 0.51 | 0.22 | 0.36 | 0.18 | 0.37 | F(3,116) = 205, p < 0.0001 |
| Sublexical Arousal | 3.36 | 0.31 | 2.76 | 0.19 | 3.30 | 0.27 | 2.77 | 0.21 | F(3,116) = 50.5, p < 0.0001 |
| Word Frequency | 0.64 | 0.75 | 0.74 | 0.76 | 0.57 | 0.78 | 0.51 | 0.75 | F(3,116) = 0.47, p = 0.69 |
| Imageability | 4.78 | 1.01 | 4.56 | 1.0 | 4.93 | 0.90 | 5.02 | 1.16 | F(3,116) = 1.11, p = 0.34 |
| Number of Syllables | 1.86 | 0.73 | 2.1 | 0.54 | 2.0 | 0.69 | 2.03 | 0.61 | F(3,116) = 0.68, p = 0.56 |
| Number of Phonemes | 5.3 | 1.36 | 5.23 | 1.10 | 5.13 | 0.89 | 4.93 | 1.20 | F(3,116) = 0.57, p = 0.63 |
| duration (ms) | 873 | 116 | 850 | 102 | 826 | 108 | 836 | 100 | F(3,116) = 1.06, p = 0.36 |
| Contrast | Anatomical Definition | MNI Coordinates | Z | K | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| LexH > LexL | L/R | Medial Frontal Gyrus (BA 9) | −3 | 56 | 20 | 5.12 | 4079 |
| L | IFG (BA 47), Temporal Pole (BA 38) | −30 | 21 | −17 | 4.48 | 672 | |
| L/R | Cuneus, Precuneus (BA 7, BA31) | −3 | −68 | 32 | 4.01 | 694 | |
| L | Posterior Cingulate Cortex (BA 23) | −8 | −47 | 26 | 3.90 | 492 | |
| LexL > LexH | L | Middle Occipital Gyrus (BA 37, 19) | −53 | −60 | −11 | 5.88 | 1244 |
| R | Middle Occipital Gyrus (BA 37) | 56 | −57 | −8 | 3.88 | 515 | |
| L | Somatosensory Cortex (BA 5) | −21 | −47 | 54 | 4.39 | 717 | |
| SubH > SubL | L | Posterior Insula (BA 13) | −42 | −15 | −1.5 | 4.86 | 861 |
| R | Posterior Insula (BA 13) | 39 | −15 | 1.5 | 4.78 | 943 | |
| R | Superior Temporal Area (BA 40, BA 22) | 51 | −38 | 24 | 4.58 | 852 | |
| R | Supplementary and Premotor area (BA 6) | 12 | −6 | 54 | 3.87 | 524 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aryani, A.; Hsu, C.-T.; Jacobs, A.M. The Sound of Words Evokes Affective Brain Responses. Brain Sci. 2018, 8, 94. https://doi.org/10.3390/brainsci8060094
Aryani A, Hsu C-T, Jacobs AM. The Sound of Words Evokes Affective Brain Responses. Brain Sciences. 2018; 8(6):94. https://doi.org/10.3390/brainsci8060094
Chicago/Turabian StyleAryani, Arash, Chun-Ting Hsu, and Arthur M. Jacobs. 2018. "The Sound of Words Evokes Affective Brain Responses" Brain Sciences 8, no. 6: 94. https://doi.org/10.3390/brainsci8060094
APA StyleAryani, A., Hsu, C. -T., & Jacobs, A. M. (2018). The Sound of Words Evokes Affective Brain Responses. Brain Sciences, 8(6), 94. https://doi.org/10.3390/brainsci8060094