More than Meets the Mind’s Eye? Preliminary Observations Hint at Heterogeneous Alpha Neuromarkers for Visual Attention


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
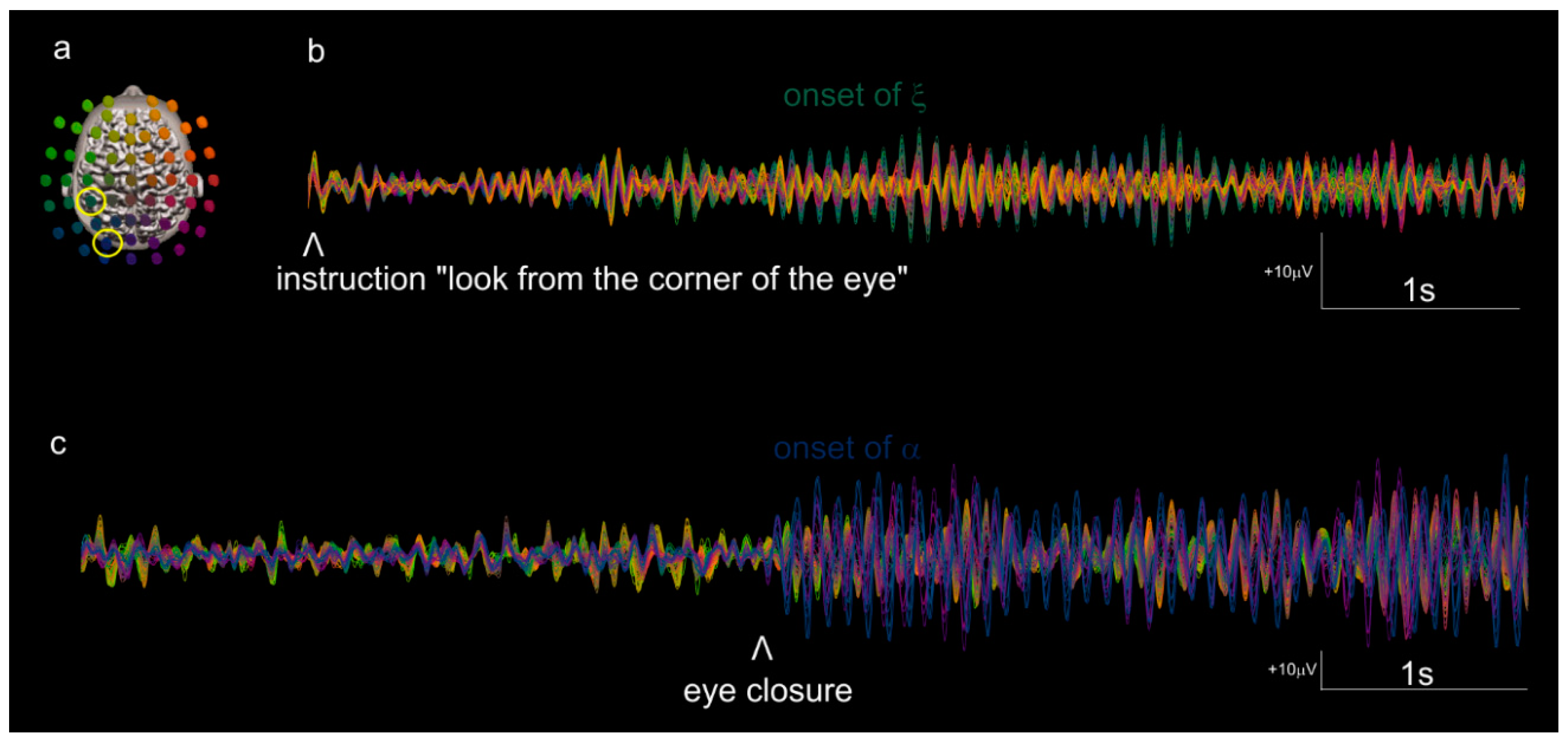
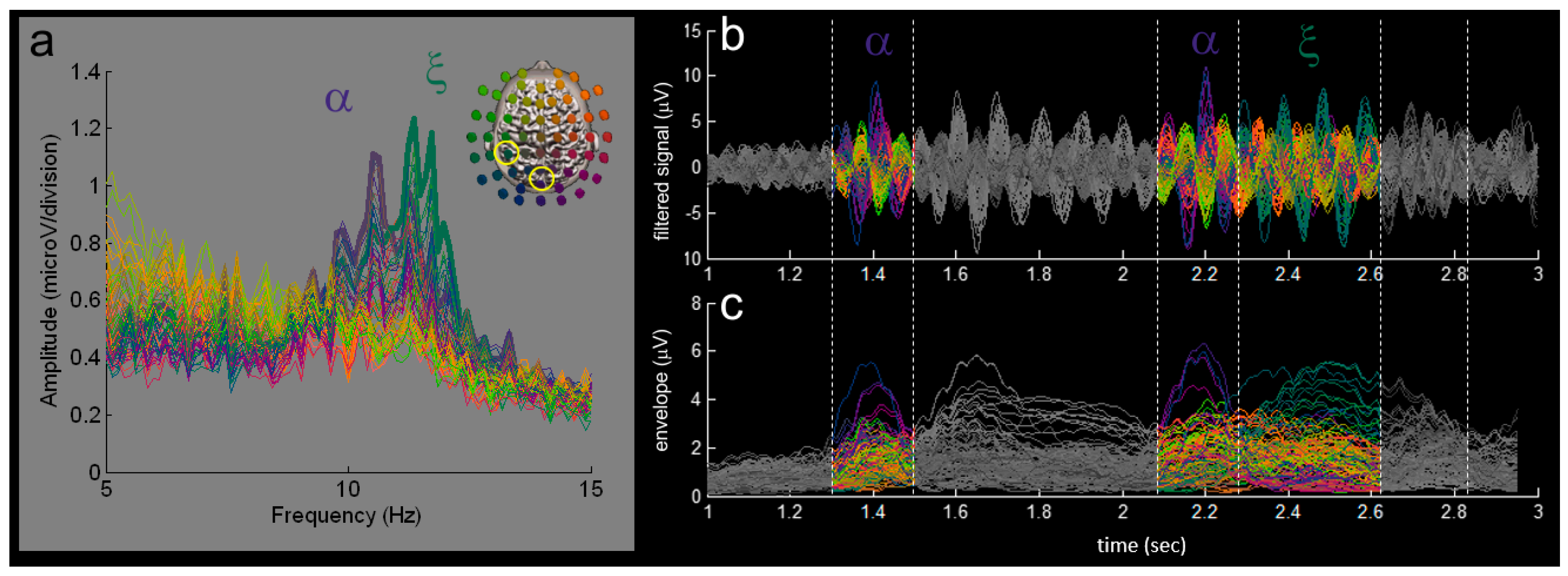
3. Results
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Posner, M.I.; Snyder, C.R.; Davidson, B.J. Attention and the detection of signals. J. Exp. Psychol. Gen. 1980, 109, 160. [Google Scholar] [CrossRef]
- James, W. The Principles of Psychology; Henry Holt and Company: New York, NY, USA, 1890; pp. 402–458. [Google Scholar]
- Von Helmholtz, H. (1896/1989). Physiological Optics (1896-2nd German Edition, translated by M. Mackeben, from Nakayama and Mackeben. Vis. Res. 1989, 29, 1631–1647. [Google Scholar]
- Hurlbert, A.; Poggio, T. Spotlight on attention. Trends Neurosci. 1985, 8, 309–311. [Google Scholar] [CrossRef]
- Johnston, W.A.; Dark, V.J. Selective attention. Annu. Rev. Psychol. 1986, 37, 43–75. [Google Scholar] [CrossRef]
- Driver, J.; Baylis, G.C. Movement and visual attention: The spotlight metaphor breaks down. J. Exp. Psychol. Hum. Percept. Perform. 1989, 15, 448. [Google Scholar] [CrossRef] [PubMed]
- Cave, K.R.; Bichot, N.P. Visuospatial attention: Beyond a spotlight model. Psychon. Bull. Rev. 1999, 6, 204–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, F. Splitting the spotlight of visual attention. Neuron 2004, 42, 524–526. [Google Scholar] [CrossRef]
- Posner, M.I.; Petersen, S.E. The attention system of the human brain. Annu. Rev. Neurosci. 1990, 13, 25–42. [Google Scholar] [CrossRef]
- Raz, A.; Buhle, J. Typologies of attentional networks. Nat. Rev. Neurosci. 2006, 7, 367. [Google Scholar] [CrossRef]
- Capotosto, P.; Babiloni, C.; Romani, G.L.; Corbetta, M. Frontoparietal cortex controls spatial attention through modulation of anticipatory alpha rhythms. J. Neurosci. 2009, 29, 5863–5872. [Google Scholar] [CrossRef]
- Berger, H. Über das elektrenkephalogramm des menschen. Eur. Arch. Psychiatry Clin. Neurosci. 1929, 87, 527–570. [Google Scholar] [CrossRef]
- Gloor, P. Hans Berger on the electroencephalogram of man. The fourteen original reports on the human electroencephalogram. Electroencephalogr. Clin. Neurophysiol. 1969, 28, 507. [Google Scholar]
- Berger, H. Ueber das Elektrenkephalogramm des Menschen. J. Psychol. Neurol. 1930, 40, 160–179. [Google Scholar]
- Adrian, E.D.; Matthews, B.H. The Berger rhythm: Potential changes from the occipital lobes in man. Brain 1934, 57, 355–385. [Google Scholar] [CrossRef]
- La Vaque, T.J. The history of EEG hans berger: Psychophysiologist. A historical vignette. J. Neurother. 1999, 3, 1–9. [Google Scholar] [CrossRef]
- Millett, D. Hans Berger: From psychic energy to the EEG. Perspect. Biol. Med. 2001, 44, 522–542. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Gómez, C.M.; Salas, R.M. Spectral structure and brain mapping of human alpha activities in different arousal states. Neuropsychobiology 1999, 39, 110–116. [Google Scholar] [CrossRef]
- Shaw, J.C. The Brain’s Alpha Rhythms and the Mind; BV Elsevier Science: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Jensen, O.; Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: Gating by inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef]
- Foxe, J.J.; Snyder, A.C. The role of alpha-band brain oscillations as a sensory suppression mechanism during selective attention. Front. Psychol. 2011, 2, 154. [Google Scholar] [CrossRef]
- Keitel, C.; Keitel, A.; Benwell, C.S.; Daube, C.; Thut, G.; Gross, J. Stimulus-driven brain rhythms within the alpha band: The attentional-modulation conundrum. J. Neurosci. 2019, 39, 3119–3129. [Google Scholar] [CrossRef]
- Ray, W.J.; Cole, H.W. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Doppelmayr, M.; Russegger, H.; Pachinger, T.; Schwaiger, J. Induced alpha band power changes in the human EEG and attention. Neurosci. Lett. 1998, 244, 73–76. [Google Scholar] [CrossRef]
- Foxe, J.J.; Simpson, G.V.; Ahlfors, S.P. Parieto-occipital ∼ 10 Hz activity reflects anticipatory state of visual attention mechanisms. Neuroreport 1998, 9, 3929–3933. [Google Scholar] [CrossRef] [PubMed]
- Worden, M.S.; Foxe, J.J.; Wang, N.; Simpson, G.V. Anticipatory biasing of visuospatial attention indexed by retinotopically specific-band electroencephalography increases over occipital cortex. J. Neurosci. 2000, 20, RC63. [Google Scholar] [CrossRef] [PubMed]
- Sauseng, P.; Klimesch, W.; Stadler, W.; Schabus, M.; Doppelmayr, M.; Hanslmayr, S.; Gruber, W.R.; Birbaumer, N. A shift of visual spatial attention is selectively associated with human EEG alpha activity. Eur. J. Neurosci. 2005, 22, 2917–2926. [Google Scholar] [CrossRef] [PubMed]
- Thut, G.; Nietzel, A.; Brandt, S.A.; Pascual-Leone, A. α-Band electroencephalographic activity over occipital cortex indexes visuospatial attention bias and predicts visual target detection. J. Neurosci. 2006, 26, 9494–9502. [Google Scholar] [CrossRef]
- Doesburg, S.M.; Roggeveen, A.B.; Kitajo, K.; Ward, L.M. Large-scale gamma-band phase synchronization and selective attention. Cereb. Cortex 2008, 18, 386–396. [Google Scholar] [CrossRef]
- Wyart, V.; Tallon-Baudry, C. Neural dissociation between visual awareness and spatial attention. J. Neurosci. 2008, 28, 2667–2679. [Google Scholar] [CrossRef]
- Pantazis, D.; Simpson, G.V.; Weber, D.L.; Dale, C.L.; Nichols, T.E.; Leahy, R.M. A novel ANCOVA design for analysis of MEG data with application to a visual attention study. NeuroImage 2009, 44, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Snyder, A.C.; Molholm, S.; Foxe, J.J. Oscillatory alpha-band mechanisms and the deployment of spatial attention to anticipated auditory and visual target locations: Supramodal or sensory-specific control mechanisms? J. Neurosci. 2011, 31, 9923–9932. [Google Scholar] [CrossRef]
- Rohenkohl, G.; Nobre, A.C. Alpha oscillations related to anticipatory attention follow temporal expectations. J. Neurosci. 2011, 31, 14076–14084. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, S.; D’Zmura, M.; Srinivasan, R. Lateralization of frequency-specific networks for covert spatial attention to auditory stimuli. Brain Topogr. 2012, 25, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.D.; Vaina, L.M. Functional roles of 10 Hz alpha-band power modulating engagement and disengagement of cortical networks in a complex visual motion task. PLoS ONE 2014, 9, e107715. [Google Scholar] [CrossRef] [PubMed]
- Ikkai, A.; Dandekar, S.; Curtis, C.E. Lateralization in alpha-band oscillations predicts the locus and spatial distribution of attention. PLoS ONE 2016, 11, e0154796. [Google Scholar] [CrossRef] [PubMed]
- Samaha, J.; Gosseries, O.; Postle, B.R. Distinct oscillatory frequencies underlie excitability of human occipital and parietal cortex. J. Neurosci. 2017, 37, 2824–2833. [Google Scholar] [CrossRef] [PubMed]
- Kizuk, S.A.; Mathewson, K.E. Power and phase of alpha oscillations reveal an interaction between spatial and temporal visual attention. J. Cogn. Neurosci. 2017, 29, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Cosmelli, D.; López, V.; Lachaux, J.P.; López-Calderón, J.; Renault, B.; Martinerie, J.; Aboitiz, F. Shifting visual attention away from fixation is specifically associated with alpha band activity over ipsilateral parietal regions. Psychophysiology 2011, 48, 312–322. [Google Scholar] [CrossRef]
- Treder, M.S.; Bahramisharif, A.; Schmidt, N.M.; Van Gerven, M.A.; Blankertz, B. Brain-computer interfacing using modulations of alpha activity induced by covert shifts of attention. J. Neuroeng. Rehabil. 2011, 8, 24. [Google Scholar] [CrossRef]
- Martel, A.; Dähne, S.; Blankertz, B. EEG predictors of covert vigilant attention. J. Neural Eng. 2014, 11, 035009. [Google Scholar] [CrossRef]
- Brüers, S.; VanRullen, R. Alpha power modulates perception independently of endogenous factors. Front. Neurosci. 2018, 12, 279. [Google Scholar] [CrossRef]
- Lehmann, D. Multichannel topography of human alpha EEG fields. Electroencephalogr. Clin. Neurophysiol. 1971, 31, 439–449. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Schimke, H.; Pachinger, T. Alpha frequency, reaction time, and the speed of processing information. J. Clin. Neurophysiol. 1996, 13, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, T.; Runnals, S. Evaluation of attention and alertness with a stimulus-brain feedback loop. Electroencephalogr. Clin. Neurophysiol. 1962, 14, 847–852. [Google Scholar] [CrossRef]
- Cooper, N.R.; Croft, R.J.; Dominey, S.J.; Burgess, A.P.; Gruzelier, J.H. Paradox lost? Exploring the role of alpha oscillations during externally vs. internally directed attention and the implications for idling and inhibition hypotheses. Int. J. Psychophysiol. 2003, 47, 65–74. [Google Scholar] [CrossRef]
- Rihs, T.A.; Michel, C.M.; Thut, G. A bias for posterior α-band power suppression versus enhancement during shifting versus maintenance of spatial attention. Neuroimage 2009, 44, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Benedek, M.; Schickel, R.J.; Jauk, E.; Fink, A.; Neubauer, A.C. Alpha power increases in right parietal cortex reflects focused internal attention. Neuropsychologia 2014, 56, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiloni, C.; Miniussi, C.; Babiloni, F.; Carducci, F.; Cincotti, F.; Del Percio, C.; Sirello, G.; Fracassi, C.; Nobre, A.C.; Rossini, P.M. Sub-second “temporal attention” modulates alpha rhythms. A high-resolution EEG study. Cogn. Brain Res. 2004, 19, 259–268. [Google Scholar] [CrossRef]
- Boncompte, G.; Villena-González, M.; Cosmelli, D.; López, V. Spontaneous alpha power lateralization predicts detection performance in an un-cued signal detection task. PLoS ONE 2016, 11, e0160347. [Google Scholar] [CrossRef]
- Foster, J.J.; Sutterer, D.W.; Serences, J.T.; Vogel, E.K.; Awh, E. Alpha-band oscillations enable spatially and temporally resolved tracking of covert spatial attention. Psychol. Sci. 2017, 28, 929–941. [Google Scholar] [CrossRef]
- Doesburg, S.M.; Bedo, N.; Ward, L.M. Top-down alpha oscillatory network interactions during visuospatial attention orienting. Neuroimage 2016, 132, 512–519. [Google Scholar] [CrossRef]
- van Schouwenburg, M.R.; Zanto, T.P.; Gazzaley, A. Spatial attention and the effects of frontoparietal alpha band stimulation. Front. Hum. Neurosci. 2017, 10, 658. [Google Scholar] [CrossRef] [PubMed]
- Proskovec, A.L.; Heinrichs-Graham, E.; Wiesman, A.I.; McDermott, T.J.; Wilson, T.W. Oscillatory dynamics in the dorsal and ventral attention networks during the reorienting of attention. Hum. Brain Mapp. 2018, 39, 2177–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognoli, E.; Lagarde, J.; DeGuzman, G.C.; Kelso, J.A.S. The phi complex as a neuromarker of human social coordination. Proc. Natl. Acad. Sci. USA 2007, 104, 8190–8195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognoli, E.; Kelso, J.A.S. The coordination dynamics of social neuromarkers. Front. Hum. Neurosci. 2015, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Tognoli, E.; Kelso, J.A.S. The metastable brain. Neuron 2014, 81, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Bazanova, O.M.; Vernon, D. Interpreting EEG alpha activity. Neurosci. Biobehav. Rev. 2014, 44, 94–110. [Google Scholar] [CrossRef] [Green Version]
- Calderon, R. Brain computer interface and neuroprosthetics. ProQuest 2007. Available online: http://purl.flvc.org/fau/fd/FA00012509 (accessed on 2 November 2019).
- Chatrian, G.E.; Lettich, E.; Nelson, P.L. Ten percent electrode system for topographic studies of spontaneous and evoked EEG activities. Am. J. EEG Technol. 1985, 25, 83–92. [Google Scholar] [CrossRef]
- Picton, T.W.; Bentin, S.; Berg, P.; Donchin, E.; Hillyard, S.A.; Johnson, R.; Miller, G.A.; Ritter, W.; Ruchkin, D.S.; Rugg, M.D.; et al. Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology 2000, 37, 127–152. [Google Scholar] [CrossRef]
- Tognoli, E.; Kelso, J.A.S. Brain coordination dynamics: True and false faces of phase synchrony and metastability. Prog. Neurobiol. 2009, 87, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Tognoli, E.; Kelso, J.A.S. Spectral dissociation of lateralized pairs of brain rhythms. arXiv 2013, arXiv:1310.7662. [Google Scholar]
- Zani, A.; Proverbio, A.M. How voluntary orienting of attention and alerting modulate costs of conflict processing. Sci. Rep. 2017, 7, 46701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulke, L.V.; Atkinson, J.; Braddick, O. Neural differences between covert and overt attention studied using EEG with simultaneous remote eye tracking. Front. Hum. Neurosci. 2016, 10, 592. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tognoli, E. More than Meets the Mind’s Eye? Preliminary Observations Hint at Heterogeneous Alpha Neuromarkers for Visual Attention. Brain Sci. 2019, 9, 307. https://doi.org/10.3390/brainsci9110307
Tognoli E. More than Meets the Mind’s Eye? Preliminary Observations Hint at Heterogeneous Alpha Neuromarkers for Visual Attention. Brain Sciences. 2019; 9(11):307. https://doi.org/10.3390/brainsci9110307
Chicago/Turabian StyleTognoli, Emmanuelle. 2019. "More than Meets the Mind’s Eye? Preliminary Observations Hint at Heterogeneous Alpha Neuromarkers for Visual Attention" Brain Sciences 9, no. 11: 307. https://doi.org/10.3390/brainsci9110307
APA StyleTognoli, E. (2019). More than Meets the Mind’s Eye? Preliminary Observations Hint at Heterogeneous Alpha Neuromarkers for Visual Attention. Brain Sciences, 9(11), 307. https://doi.org/10.3390/brainsci9110307