Matrix- and Technology-Dependent Stability and Bioaccessibility of Strawberry Anthocyanins during Storage


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Chemicals
2.2. Preparation of Juice Formulation
2.3. Processing Equipment and Conditions
2.4. Sample Handling and Storage
2.5. In-Vitro Digestion for Bioaccessibility
2.6. Microbiological, Chemical and Physical Analyses
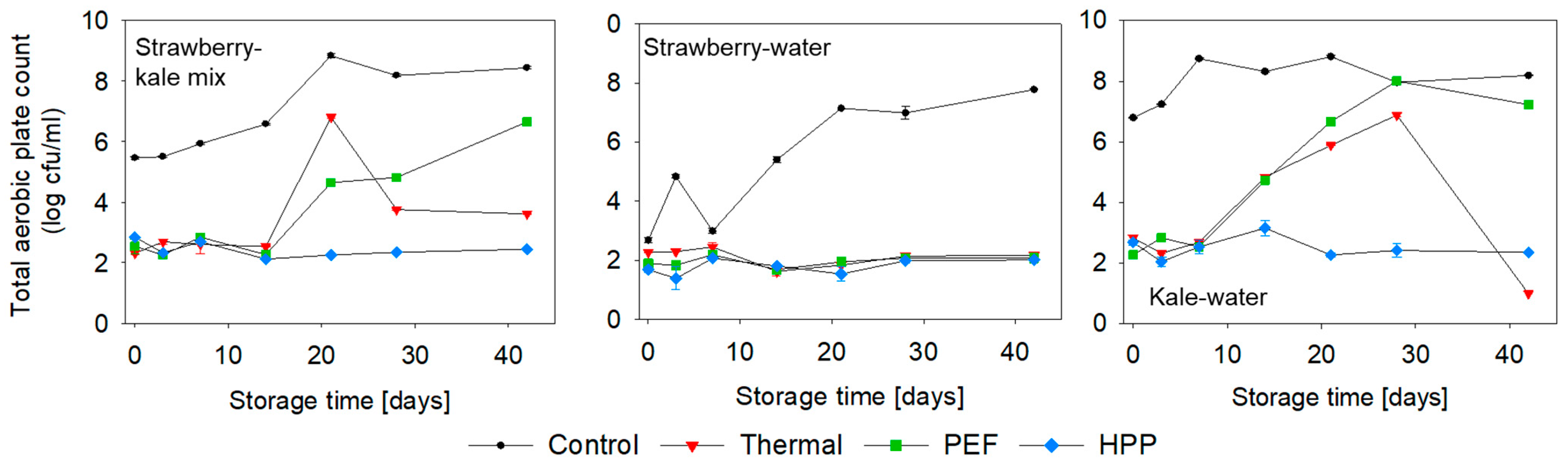
2.6.1. Microbiological Quality
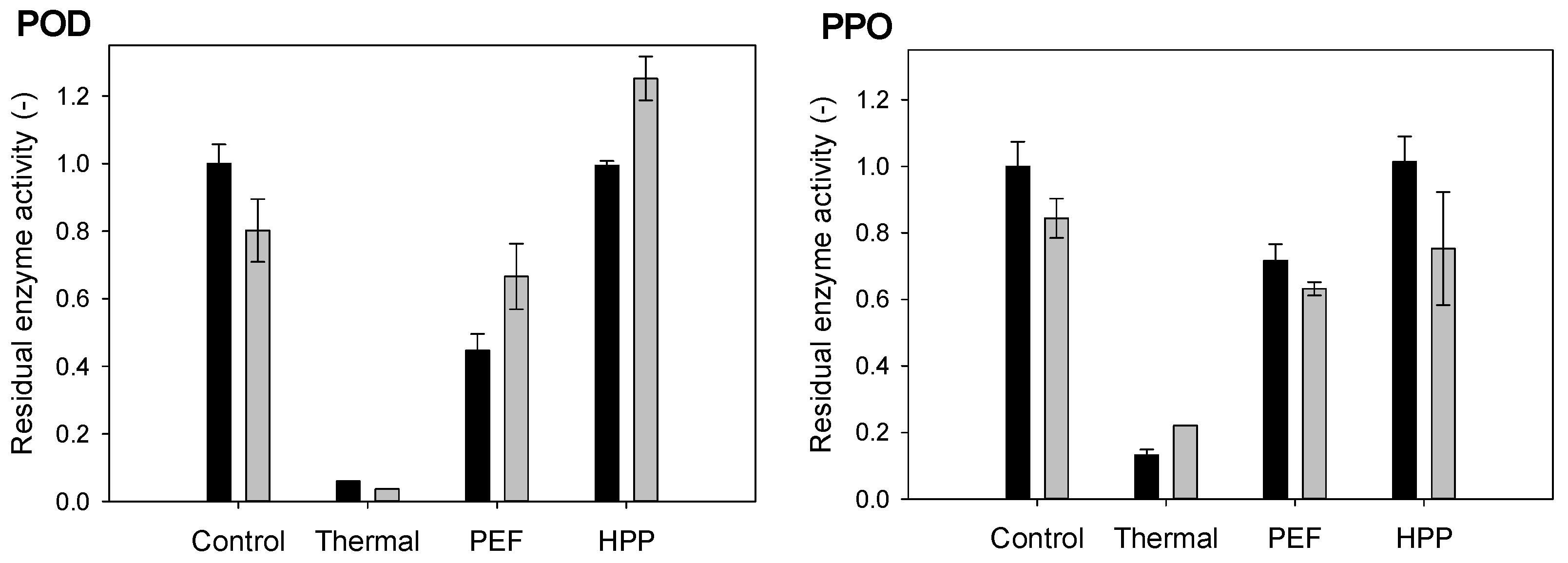
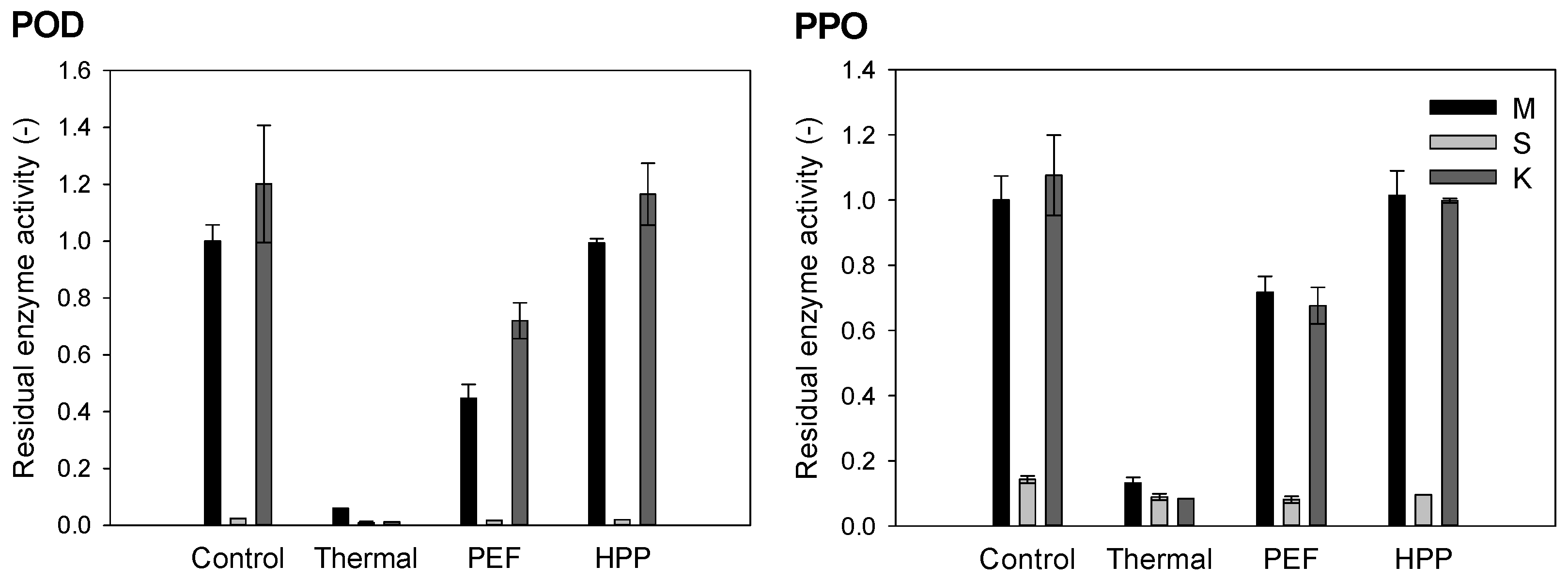
2.6.2. Residual Enzyme Activity Measurements of Peroxidase (POD) and Polyphenoloxidase (PPO)
2.6.3. Color Determination (CIE-L*a*b*)
2.6.4. Ascorbic Acid/Dehydroascorbic Acid
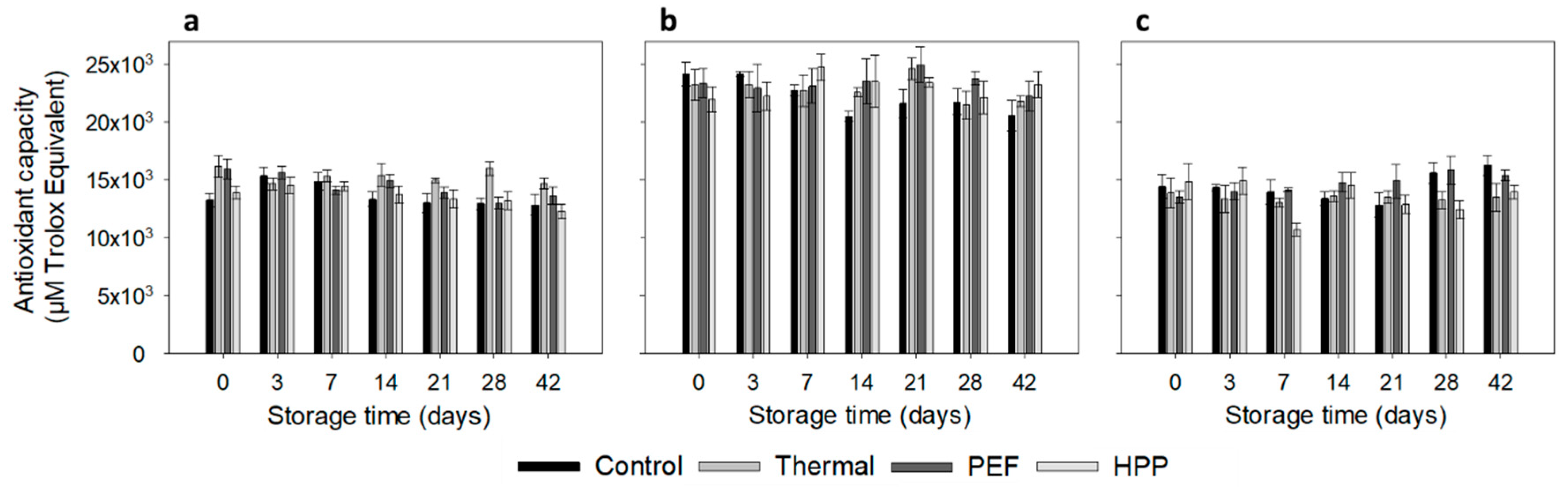
2.6.5. Antioxidant Capacity (ORAC)
2.6.6. Total Monomeric Anthocyanin Content
2.6.7. Anthocyanin Kinetics
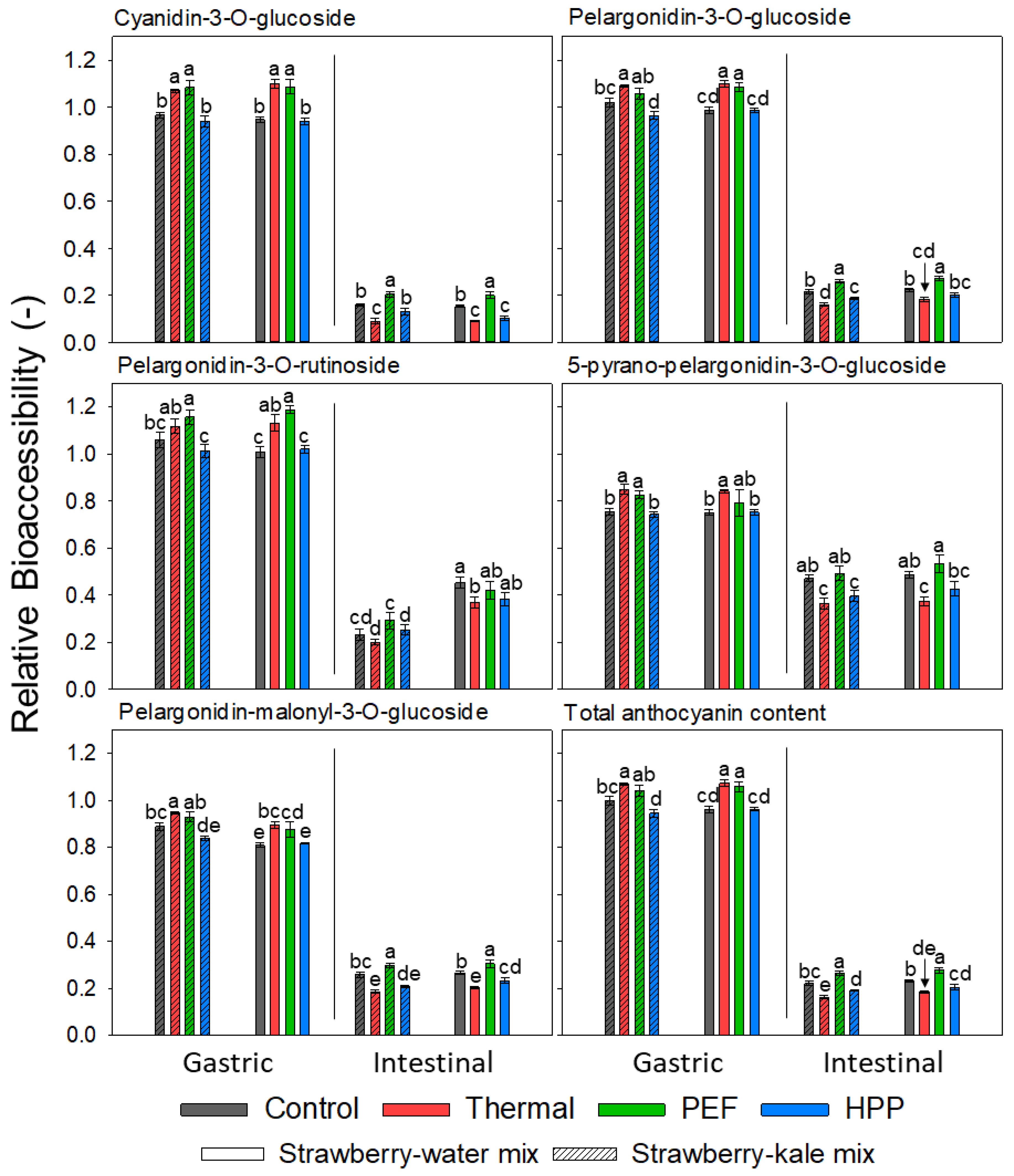
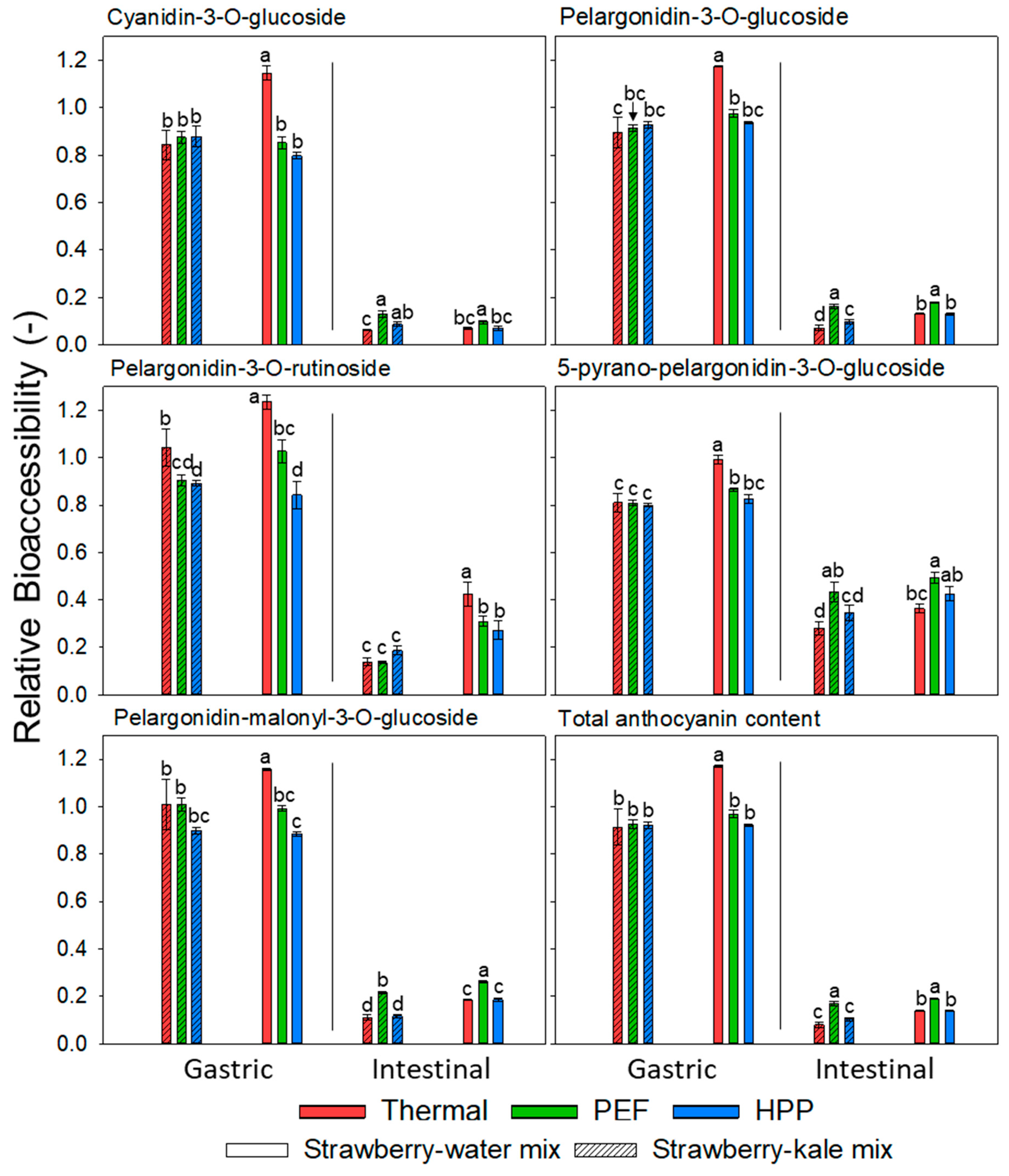
2.6.8. Relative Bioaccessibility of Anthocyanins
2.7. Statistical Analyses
3. Results and Discussion
3.1. Microbiological Quality
3.2. Residual Enzyme Activity of Peroxidase (POD) and Polyphenoloxidase (PPO)
3.3. Ascorbic Acid/Dehydroascorbic Acid
3.4. Color Difference Dependent on Processing during Storage
3.5. Antioxidant Capacity
3.6. Anthocyanin Degradation Kinetics during Storage
3.7. Relative Bioaccessibility (BA) of Anthocyanins before and after Storage
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes da Silva, F.L.; Escribano-Bailón, M.T.; Pérez Alonso, J.J.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanin pigments in strawberry. LWT Food Sci. Technol. 2007, 40, 374–382. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Rein, M.J.; Heinonen, M. Stability and Enhancement of Berry Juice Color. J. Agric. Food Chem. 2004, 52, 3106–3114. [Google Scholar] [CrossRef] [PubMed]
- Hanuka Katz, I.; Eran Nagar, E.; Okun, Z.; Shpigelman, A. The Link between Polyphenol Structure, Antioxidant Capacity and Shelf-Life Stability in the Presence of Fructose and Ascorbic Acid. Molecules 2020, 25, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, R.; Okun, Z.; Shpigelman, A. The Influence of Chemical Structure and the Presence of Ascorbic Acid on Anthocyanins Stability and Spectral Properties in Purified Model Systems. Foods 2019, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Cheftel, J.C. Review: High-pressure, microbial inactivation and food preservation. Food Sci. Technol. Int. 1995, 1, 75–90. [Google Scholar] [CrossRef]
- Castro, A.J.; Barbosa-Cánovas, G.v.; Swanson, B.G. Microbial inactivation of foods by pulsed electric fields. J. Food Process. Preserv. 1993, 17, 47–73. [Google Scholar] [CrossRef]
- Hoover, D.G.; Metrick, C.; Papineau, A.M.; Farkas, D.F.; Knorr, D. Biological effects of high hydrostatic pressure on food microorganisms. Food Technol. 1989, 43, 99–107. [Google Scholar]
- Oliveira, A.; Coelho, M.; Alexandre, E.M.C.; Almeida, D.P.F.; Pintado, M. Long-Term Frozen Storage and Pasteurization Effects on Strawberry Polyphenols Content. Food Bioprocess Technol. 2015, 8, 1838–1844. [Google Scholar] [CrossRef]
- da Silveira, T.F.F.; Cristianini, M.; Kuhnle, G.G.; Ribeiro, A.B.; Filho, J.T.; Godoy, H.T. Anthocyanins, non-anthocyanin phenolics, tocopherols and antioxidant capacity of açaí juice (Euterpe oleracea) as affected by high pressure processing and thermal pasteurization. Innov. Food Sci. Emerg. Technol. 2019, 55, 88–96. [Google Scholar] [CrossRef]
- Ma, Y.; Yin, X.; Bi, X.; Su, F.; Liang, Z.; Luo, M.; Fu, D.; Xing, Y.; Che, Z. Physicochemical properties and bioactive compounds of fermented pomegranate juice as affected by high-pressure processing and thermal treatment. Int. J. Food Prop. 2019, 22, 1250–1269. [Google Scholar] [CrossRef] [Green Version]
- Marszałek, K.; Mitek, M.; Skąpska, S. The effect of thermal pasteurization and high pressure processing at cold and mild temperatures on the chemical composition, microbial and enzyme activity in strawberry purée. Innov. Food Sci. Emerg. Technol. 2015, 27, 48–56. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; Da Pieve, S.; Butler, F.; Da, S.; Butler, F. Impact of high pressure processing on total antioxidant activity, phenolic, ascorbic acid, anthocyanin content and colour of strawberry and blackberry purées. Innov. Food Sci. Emerg. Technol. 2009, 10, 308–313. [Google Scholar] [CrossRef]
- Torres, B.; Tiwari, B.K.; Patras, A.; Cullen, P.J.; Brunton, N.; O’Donnell, C.P.; Donnell, C.P.O. Stability of anthocyanins and ascorbic acid of high pressure processed blood orange juice during storage. Innov. Food Sci. Emerg. Technol. 2011, 12, 93–97. [Google Scholar] [CrossRef]
- Odriozola-Serrano, I.; Soliva-Fortuny, R.; Gimeno-Añó, V.; Martín-Belloso, O. Kinetic study of anthocyanins, vitamin C, and antioxidant capacity in strawberry juices treated by high-intensity pulsed electric fields. J. Agric. Food Chem. 2008, 56, 8387–8393. [Google Scholar] [CrossRef]
- Odriozola-Serrano, I.; Soliva-Fortuny, R.; Martín-Belloso, O. Phenolic acids, flavonoids, vitamin C and antioxidant capacity of strawberry juices processed by high-intensity pulsed electric fields or heat treatments. Eur. Food Res. Technol. 2008, 228, 239–248. [Google Scholar] [CrossRef]
- Dobson, G.; McDougall, G.J.; Stewart, D.; Cubero, M.Á.; Karjalainen, R.O. Effects of Juice Matrix and Pasteurization on Stability of Black Currant Anthocyanins during Storage. J. Food Sci. 2017, 82, 44–52. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; De Ancos, B.; Sánchez-Moreno, C.; Cano, M.P.; Elez-Martínez, P.; Martín-Belloso, O. Impact of food matrix and processing on the in vitro bioaccessibility of vitamin C, phenolic compounds, and hydrophilic antioxidant activity from fruit juice-based beverages. J. Funct. Foods 2015, 14, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Morales-De La Peña, M.; Salvia-Trujillo, L.; Rojas-Graü, M.A.; Martín-Belloso, O. Changes on phenolic and carotenoid composition of high intensity pulsed electric field and thermally treated fruit juice-soymilk beverages during refrigerated storage. Food Chem. 2011, 129, 982–990. [Google Scholar] [CrossRef]
- Stübler, A.-S.; Lesmes, U.; Heinz, V.; Rauh, C.; Shpigelman, A.; Aganovic, K. Digestibility, antioxidative activity and stability of plant protein-rich products after processing and formulation with polyphenol-rich juices: Kale and kale–strawberry as a model. Eur. Food Res. Technol. 2019, 245, 2499–2514. [Google Scholar] [CrossRef]
- Stübler, A.-S.; Lesmes, U.; Juadjur, A.; Heinz, V.; Rauh, C.; Shpigelman, A.; Aganovic, K. Impact of pilot-scale processing (thermal, PEF, HPP) on the stability and bioaccessibility of polyphenols and proteins in mixed protein and polyphenol-rich juice systems. Innov. Food Sci. Emerg. Technol. 2020, 64, 102426. [Google Scholar] [CrossRef]
- Chung, C.; Rojanasasithara, T.; Mutilangi, W.; McClements, D.J. Enhanced stability of anthocyanin-based color in model beverage systems through whey protein isolate complexation. Food Res. Int. 2015, 76, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Rojanasasithara, T.; Mutilangi, W.; McClements, D.J. Stability improvement of natural food colors: Impact of amino acid and peptide addition on anthocyanin stability in model beverages. Food Chem. 2017, 218, 277–284. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; Tiwari, B.K.; Butler, F. Stability and Degradation Kinetics of Bioactive Compounds and Colour in Strawberry Jam during Storage. Food Bioprocess Technol. 2011, 4, 1245–1252. [Google Scholar] [CrossRef]
- Buvé, C.; Kebede, B.T.; De Batselier, C.; Carrillo, C.; Pham, H.T.T.; Hendrickx, M.; Grauwet, T.; Van Loey, A. Kinetics of colour changes in pasteurised strawberry juice during storage. J. Food Eng. 2018, 216, 42–51. [Google Scholar] [CrossRef]
- Bonerz, D.; Würth, K.; Will, F.; Patz, C.; Quast, P.; Hillebrand, S.; Winterhalter, P.; Dietrich, H. Aging process in strawberry juices and concentrates: Degradation kinetics of monomeric anthocyanins and the formation of new aging pigments. Dtsch. Leb. 2011, 107, 24–32. [Google Scholar]
- Alighourchi, H.; Barzegar, M. Some physicochemical characteristics and degradation kinetic of anthocyanin of reconstituted pomegranate juice during storage. J. Food Eng. 2009, 90, 179–185. [Google Scholar] [CrossRef]
- Cao, X.; Bi, X.; Huang, W.; Wu, J.; Hu, X.; Liao, X. Changes of quality of high hydrostatic pressure processed cloudy and clear strawberry juices during storage. Innov. Food Sci. Emerg. Technol. 2012, 16, 181–190. [Google Scholar] [CrossRef]
- Verbeyst, L.; Oey, I.; Van der Plancken, I.; Hendrickx, M.; Van Loey, A. Kinetic study on the thermal and pressure degradation of anthocyanins in strawberries. Food Chem. 2010, 123, 269–274. [Google Scholar] [CrossRef]
- Su, G.; Zhu, S.; Xu, M.; Ramaswamy, H.S.; Lin, Y.; Yu, Y. Pressure Degradation Kinetics of Anthocyanin Pigment and Visual Color of Chinese Bayberry Juice. Int. J. Food Prop. 2016, 19, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, M.; Ludikhuyze, L.; Van Den Broeck, I.; Weemaes, C. Effects of high pressure on enzymes related to food quality. Trends Food Sci. Technol. 1998, 9, 197–203. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, X.S.; Chen, F.; Wu, J.H.; Liao, X.J.; Wang, Z.F. Stability and colour characteristics of PEF-treated cyanidin-3-glucoside during storage. Food Chem. 2008, 106, 669–676. [Google Scholar] [CrossRef]
- Eran Nagar, E.; Berenshtein, L.; Katz, I.H.; Lesmes, U.; Okun, Z.; Shpigelman, A. The impact of chemical structure on polyphenol bioaccessibility, as a function of processing, cell wall material and pH: A model system. J. Food Eng. 2020, 289, 110304. [Google Scholar] [CrossRef]
- Carrillo, C.; Buvé, C.; Panozzo, A.; Grauwet, T.; Hendrickx, M. Role of structural barriers in the in vitro bioaccessibility of anthocyanins in comparison with carotenoids. Food Chem. 2017, 227, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Nagar, E.E.; Okun, Z.; Shpigelman, A. Digestive fate of polyphenols: Updated view of the influence of chemical structure and the presence of cell wall material. Curr. Opin. Food Sci. 2020, 31, 38–46. [Google Scholar] [CrossRef]
- Buniowska, M.; Carbonell-Capella, J.M.; Frigola, A.; Esteve, M.J. Bioaccessibility of bioactive compounds after non-thermal processing of an exotic fruit juice blend sweetened with Stevia rebaudiana. Food Chem. 2017, 221, 1834–1842. [Google Scholar] [CrossRef]
- Oksuz, T.; Tacer-Caba, Z.; Nilufer-Erdil, D.; Boyacioglu, D. Changes in bioavailability of sour cherry (Prunus cerasus L.) phenolics and anthocyanins when consumed with dairy food matrices. J. Food Sci. Technol. 2019, 56, 4177–4188. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. In vitro bioaccessibility of health-related compounds as affected by the formulation of fruit juice- and milk-based beverages. Food Res. Int. 2014, 62, 771–778. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Yue-Ming, J.; Zauberman, G.; Fuchs, Y. Partial purification and some properties of polyphenol oxidase extracted from litchi fruit pericarp. Postharvest Biol. Technol. 1997, 10, 221–228. [Google Scholar] [CrossRef]
- Cserhalmi, Z.; Sass-Kiss, Á.; Tóth-Markus, M.; Lechner, N. Study of pulsed electric field treated citrus juices. Innov. Food Sci. Emerg. Technol. 2006, 7, 49–54. [Google Scholar] [CrossRef]
- Drlange. Colour review. Drlange Appl. Rep. No. 8.0e 1994. [Google Scholar]
- Wibowo, S.; Afuape, A.L.; de Man, S.; Bernaert, N.; Van Droogenbroeck, B.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Thermal processing of kale purée: The impact of process intensity and storage on different quality related aspects. Innov. Food Sci. Emerg. Technol. 2019, 58, 102213. [Google Scholar] [CrossRef]
- Kneifel, W.; Sommer, R. HPLC-Methode zur Bestimmung von Vitamin C in Milch, Molke und Molkegetränken. Z. Lebensm. Unters. Forsch. 1985, 181, 107–110. [Google Scholar] [CrossRef]
- Juadjur, A.; Winterhalter, P. Development of a novel adsorptive membrane chromatographic method for the fractionation of polyphenols from bilberry. J. Agric. Food Chem. 2012, 60, 2427–2433. [Google Scholar] [CrossRef]
- Wibowo, S.; Essel, E.A.; De Man, S.; Bernaert, N.; Van Droogenbroeck, B.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Comparing the impact of high pressure, pulsed electric field and thermal pasteurization on quality attributes of cloudy apple juice using targeted and untargeted analyses. Innov. Food Sci. Emerg. Technol. 2019, 54, 64–77. [Google Scholar] [CrossRef]
- Sánchez-Vega, R.; Rodríguez-Roque, M.J.; Elez-Martínez, P.; Martín-Belloso, O. Impact of critical high-intensity pulsed electric field processing parameters on oxidative enzymes and color of broccoli juice. J. Food Process. Preserv. 2020, 44, 1–10. [Google Scholar] [CrossRef]
- Heinz, V.; Alvarez, I.; Angersbach, A.; Knorr, D. Preservation of liquid foods by high intensity pulsed electric fields - Basic concepts for process design. Trends Food Sci. Technol. 2001, 12, 103–111. [Google Scholar] [CrossRef]
- Aguiló-Aguayo, I.; Soliva-Fortuny, R.; Martín-Belloso, O. High-intensity pulsed electric fields processing parameters affecting polyphenoloxidase activity of strawberry juice. J. Food Sci. 2010, 75, 641–646. [Google Scholar] [CrossRef]
- Kamat, S.S.; Dash, K.K.; Balasubramaniam, V.M. Quality changes in combined pressure-thermal treated acidified vegetables during extended ambient temperature storage. Innov. Food Sci. Emerg. Technol. 2018, 49, 146–157. [Google Scholar] [CrossRef]
- Terefe, N.S.; Yang, Y.H.; Knoerzer, K.; Buckow, R.; Versteeg, C. High pressure and thermal inactivation kinetics of polyphenol oxidase and peroxidase in strawberry puree. Innov. Food Sci. Emerg. Technol. 2010, 11, 52–60. [Google Scholar] [CrossRef]
- Roidoung, S.; Dolan, K.D.; Siddiq, M. Gallic acid as a protective antioxidant against anthocyanin degradation and color loss in vitamin-C fortified cranberry juice. Food Chem. 2016, 210, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Rimbach, G. Ascorbigen: Chemistry, occurrence, and biologic properties. Clin. Dermatol. 2009, 27, 217–224. [Google Scholar] [CrossRef]
- Hrncirik, K.; Valusek, J.; Velisek, J. Investigation of ascorbigen as a breakdown product of glucobrassicin autolysis in Brassica vegetables. Eur. Food Res. Technol. 2001, 212, 576–581. [Google Scholar] [CrossRef]
- Hrnčiřík, K.; Valušek, J.; Velíšek, J. A study on the formation and stability of ascorbigen in an aqueous system. Food Chem. 1998, 63, 349–355. [Google Scholar] [CrossRef]
- Aaby, K.; Grimsbo, I.H.; Hovda, M.B.; Rode, T.M. Effect of high pressure and thermal processing on shelf life and quality of strawberry purée and juice. Food Chem. 2018, 260, 115–123. [Google Scholar] [CrossRef]
- Chen, J.; Tao, X.Y.; Sun, A.D.; Wang, Y.; Liao, X.J.; Li, L.N.; Zhang, S. Influence of pulsed electric field and thermal treatments on the quality of blueberry juice. Int. J. Food Prop. 2014, 17, 1419–1427. [Google Scholar] [CrossRef]
- Kumar, R.; Vijayalakshmi, S.; Rajeshwara, R.; Sunny, K.; Nadanasabapathi, S. Effect of storage on thermal, pulsed electric field and combination processed mango nectar. J. Food Meas. Charact. 2019, 13, 131–143. [Google Scholar] [CrossRef]
- Dziadek, K.; Kopeć, A.; Dróżdż, T.; Kiełbasa, P.; Ostafin, M.; Bulski, K.; Oziembłowski, M. Effect of pulsed electric field treatment on shelf life and nutritional value of apple juice. J. Food Sci. Technol. 2019, 56, 1184–1191. [Google Scholar] [CrossRef] [Green Version]
- You, Y.; Li, N.; Han, X.; Guo, J.; Zhao, Y.; Liu, G.; Huang, W.; Zhan, J. Influence of different sterilization treatments on the color and anthocyanin contents of mulberry juice during refrigerated storage. Innov. Food Sci. Emerg. Technol. 2018, 48, 1–10. [Google Scholar] [CrossRef]
- Rodrigo, D.; van Loey, A.; Hendrickx, M. Combined thermal and high pressure colour degradation of tomato puree and strawberry juice. J. Food Eng. 2007, 79, 553–560. [Google Scholar] [CrossRef]
- Sulaiman, A.; Farid, M.; Silva, F.V.M. Strawberry puree processed by thermal, high pressure, or power ultrasound: Process energy requirements and quality modeling during storage. Food Sci. Technol. Int. 2017, 23, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Arts, M.J.T.J.; Haenen, G.R.M.M.; Wilms, L.C.; Beetstra, S.A.J.N.; Heijnen, C.G.M.; Voss, H.P.; Bast, A. Interactions between flavonoids and proteins: Effect on the total antioxidant capacity. J. Agric. Food Chem. 2002, 50, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Perales, S.; Lagarda, M.J.; Barberá, R.; Clemente, G.; Farré, R. Influence of storage and in vitro gastrointestinal digestion on total antioxidant capacity of fruit beverages. J. Food Compos. Anal. 2011, 24, 87–94. [Google Scholar] [CrossRef]
- Joubran, Y.; Moscovici, A.; Lesmes, U. Antioxidant activity of bovine alpha lactalbumin Maillard products and evaluation of their in vitro gastro-duodenal digestive proteolysis. Food Funct. 2015, 6, 1229–1240. [Google Scholar] [CrossRef]
- Andrés, V.; Villanueva, M.J.; Tenorio, M.D. The effect of high-pressure processing on colour, bioactive compounds, and antioxidant activity in smoothies during refrigerated storage. Food Chem. 2016, 192, 328–335. [Google Scholar] [CrossRef]
- Tadapaneni, R.K.; Banaszewski, K.; Patazca, E.; Edirisinghe, I.; Cappozzo, J.; Jackson, L.; Burton-Freeman, B. Effect of high-pressure processing and milk on the anthocyanin composition and antioxidant capacity of strawberry-based beverages. J. Agric. Food Chem. 2012, 60, 5795–5802. [Google Scholar] [CrossRef]
- Cao, S.; Liu, L.; Pan, S.; Lu, Q.; Xu, X. A comparison of two determination methods for studying degradation kinetics of the major anthocyanins from blood orange. J. Agric. Food Chem. 2009, 57, 245–249. [Google Scholar] [CrossRef]
- Sinela, A.M.; Rawat, N.; Mertz, C.; Achir, N.; Fulcrand, H.; Dornier, M. Anthocyanins degradation during storage of Hibiscus sabdariffa extract and evolution of its degradation products. Food Chem. 2017, 214, 234–241. [Google Scholar] [CrossRef]
- Marszałek, K.; Woźniak, Ł.; Skąpska, S.; Mitek, M. High pressure processing and thermal pasteurization of strawberry purée: Quality parameters and shelf life evaluation during cold storage. J. Food Sci. Technol. 2017, 54, 832–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Lin, Y.; Zhan, Y.; He, J.; Zhu, S. Effect of high pressure processing on the stability of anthocyanin, ascorbic acid and color of Chinese bayberry juice during storage. J. Food Eng. 2013, 119, 701–706. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; O’Donnell, C.; Tiwari, B.K. Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- Mena, P.; Martí, N.; García-Viguera, C. Varietal blends as a way of optimizing and preserving the anthocyanin content of pomegranate (Punica granatum L.) juices. J. Agric. Food Chem. 2014, 62, 6936–6943. [Google Scholar] [CrossRef]
- Loypimai, P.; Moongngarm, A.; Chottanom, P. Thermal and pH degradation kinetics of anthocyanins in natural food colorant prepared from black rice bran. J. Food Sci. Technol. 2016, 53, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, K.; Howard, L.R.; Brownmiller, C.; Prior, R.L. Changes in Chokeberry (Aronia melanocarpa L.) polyphenols during juice processing and storage. J. Agric. Food Chem. 2014, 62, 4018–4025. [Google Scholar] [CrossRef]
- Rein, M.J.; Ollilainen, V.; Vahermo, M.; Yli-Kauhaluoma, J.; Heinonen, M. Identification of novel pyranoanthocyanins in berry juices. Eur. Food Res. Technol. 2005, 220, 239–244. [Google Scholar] [CrossRef]
- Yang, P.; Yuan, C.; Wang, H.; Han, F.; Liu, Y.; Wang, L.; Liu, Y. Stability of anthocyanins and their degradation products from cabernet sauvignon red wine under gastrointestinal pH and temperature conditions. Molecules 2018, 23, 354. [Google Scholar] [CrossRef] [Green Version]
- Marquez, A.; Serratosa, M.P.; Merida, J. Influence of bottle storage time on colour, phenolic composition and sensory properties of sweet red wines. Food Chem. 2013, 146, 507–514. [Google Scholar] [CrossRef]
- Heaney, R.P. Bioavailability of Nutrients and Other Bioactive Components from Dietary Supplements Factors Influencing the Measurement of Bioavailability, Taking Calcium as a Model 1. J. Nutr. 2001, 131, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F. The stomach as a site for anthocyanins absorption from food. FEBS Lett. 2003, 544, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Welch, C.; Wu, Q.; Simon, J. Recent Advances in Anthocyanin Analysis and Characterization. Curr. Anal. Chem. 2008, 4, 75–101. [Google Scholar] [CrossRef] [Green Version]
- Bermúdez-Soto, M.J.; Tomás-Barberán, F.A.; García-Conesa, M.T. Stability of polyphenols in chokeberry (Aronia melanocarpa) subjected to in vitro gastric and pancreatic digestion. Food Chem. 2007, 102, 865–874. [Google Scholar] [CrossRef]
- McDougall, G.J.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Assessing potential bioavailability of raspberry anthocyanins using an in vitro digestion system. J. Agric. Food Chem. 2005, 53, 5896–5904. [Google Scholar] [CrossRef] [PubMed]
- Sengul, H.; Surek, E.; Nilufer-Erdil, D. Investigating the effects of food matrix and food components on bioaccessibility of pomegranate (Punica granatum) phenolics and anthocyanins using an in-vitro gastrointestinal digestion model. Food Res. Int. 2014, 62, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Rein, M.J. Copigmentation Reactions and Color Stability of Berry Anthocyanins; Faculty of Agriculture and Forestry of the University of Helsinki: Helsinki, Finland, 2005; ISBN 9521022922. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Strawberry-Kale Mix | Strawberry-Water | Kale-Water |
|---|---|---|---|
| Total soluble solids (°Brix) | 6.78 ± 0.24 | 4.60 ± 0.04 | 2.71 ± 0.08 |
| Protein content (g/100 g) | 1.34 ± 0.08 | 0.36 ± 0.03 | 1.03 ± 0.04 |
| Sugar (g/100 g) | 3.77 ± 0.04 | 3.02 ± 0.01 | 0.67 ± 0.02 |
| After Processing (Day 0) | After Storage (Day 42) | ||
|---|---|---|---|
| Strawberry–kale mix | Control | 4.94 ± 0.23 bc | 1.16 ± 0.01 d |
| Thermal | 5.70 ± 0.11 a | 2.00 ± 0.00 a | |
| PEF | 5.08 ± 0.23 abc | 1.55 ± 0.13 bc | |
| HPP | 4.33 ± 0.04 cd | 1.35 ± 0.01 cd | |
| Strawberry–water mix | Control | 4.92 ± 0.45 bc | 1.54 ± 0.04 bc |
| Thermal | 5.33 ± 0.03 ab | 1.84 ± 0.13 ab | |
| PEF | 4.96 ± 0.19 bc | 1.53 ± 0.10 bc | |
| HPP | 4.18 ± 0.17 d | 1.45 ± 0.12 cd |
| ΔE | Thermal | PEF | HPP |
|---|---|---|---|
| Strawberry–water mix | 4.04 ± 0.04 b | 4.68 ± 0.01 a | 0.79 ± 0.06 c |
| Strawberry–kale mix | 8.22 ± 0.09 a | 7.67 ± 0.09 b | 0.86 ± 0.06 c |
| Kale–water mix | 5.03 ± 0.25 a | 3.90 ± 0.23 b | 0.61 ± 0.30 c |
| Day 3 | Day 7 | Day 14 | Day 21 | Day 28 | Day 42 | |
|---|---|---|---|---|---|---|
| Control | 1.31 ± 0.19 f | 2.23 ± 0.12 e | 3.03 ± 0.07 d | 3.66 ± 0.02 c | 4.40 ± 0.01 b | 5.75 ± 0.04 a |
| Thermal | 0.63 ± 0.05 c | 1.71 ± 0.05 a | 2.09 ± 0.43 a | 2.07 ± 0.39 a | 1.81 ± 0.04 a | 1.04 ± 0.12 b |
| PEF | 1.17 ± 0.06 d | 4.33 ± 0.04 a | 4.31 ± 0.16 ab | 4.05 ± 0.04 b | 4.05 ± 0.07 b | 3.04 ± 0.05 c |
| HPP | 0.76 ± 0.08 d | 1.39 ± 0.20 c | 2.33 ± 0.19 b | 2.40 ± 0.14 b | 2.59 ± 0.02 b | 3.53 ± 0.05 a |
| Formulation | ProCessing | Cyanidin-3-O-Glucoside | Pelargonidin-3-O-Glucoside | Pelargonidin-3-O-Rutinoside | 5-Pyrano-Pelargonidin-3-O-Glucoside | Pelargonidin-3-O-Malonyl-Glucoside | Total AnthocyAnin Content (Sum of AnThocyanins) | Monomeric AnThocyanin Content |
|---|---|---|---|---|---|---|---|---|
| Strawberry–kale mix | Control | 4.40 ± 0.36 aB | 3.64 ± 0.16 aC | 3.56 ± 0.04 aC | 2.45 ± 0.14 aD | 4.47 ± 0.18 aB | 3.71 ± 0.09 a | 2.86 ± 0.03 a |
| Thermal | 3.18 ± 0.02 bA | 2.67 ± 0.01 bBC | 1.96 ± 0.02 bD | 1.86 ± 0.03 bD | 2.81 ± 0.01 bB | 2.69 ± 0.01 b | 2.44 ± 0.06 b | |
| PEF | 2.83 ± 0.21 bA | 2.42 ± 0.27 bcA | 1.71 ± 0.21 cB | 1.57 ± 0.24 bcB | 2.64 ± 0.19 bA | 2.22 ± 0.27 c | 2.20 ± 0.08 c | |
| HPP | 2.77 ± 0.06 bB | 2.21 ± 0.04 cC | 1.75 ± 0.03 bcD | 1.51 ± 0.05 cdE | 2.86 ± 0.03 bB | 2.29 ± 0.04 c | 1.77 ± 0.05 d | |
| Strawberry–water | Control | 1.89 ± 0.05 cB | 1.17 ± 0.02 dC | 1.10 ± 0.02 dC | 1.14 ± 0.05 eC | 1.87 ± 0.03 cB | 1.28 ± 0.03 d | 1.34 ± 0.12 e |
| Thermal | 1.35 ± 0.03 dA | 0.98 ± 0.06 dB | 0.61 ± 0.06 fC | 0.76 ± 0.02 fC | 1.03 ± 0.06 eB | 1.00 ± 0.06 d | 0.65 ± 0.04 f | |
| PEF | 1.75 ± 0.04 cdA | 1.13 ± 0.03 dCD | 0.86 ± 0.03 deE | 1.01 ± 0.05 efD | 1.30 ± 0.04 deB | 1.18 ± 0.03 d | 1.16 ± 0.12 e | |
| HPP | 1.71 ± 0.06 cdA | 1.11 ± 0.05 dCD | 0.86 ± 0.04 eD | 1.22 ± 0.18 deBC | 1.47 ± 0.04 dAB | 1.18 ± 0.04 d | 1.19 ± 0.04 e |
| Formulation | Processing | Pearson Correlation |
|---|---|---|
| Strawberry-water | Control | 0.782 (<0.05) |
| Thermal | 0.421 (>0.05) | |
| PEF | 0.528 (<0.05) | |
| HPP | 0.647 (<0.05) | |
| Strawberry-kale mix | Control | −0.767 (<0.05) |
| Thermal | −0.236 (>0.05) | |
| PEF | −0.640 (<0.05) | |
| HPP | −0.956 (<0.05) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stübler, A.-S.; Böhmker, L.; Juadjur, A.; Heinz, V.; Rauh, C.; Shpigelman, A.; Aganovic, K. Matrix- and Technology-Dependent Stability and Bioaccessibility of Strawberry Anthocyanins during Storage. Antioxidants 2021, 10, 30. https://doi.org/10.3390/antiox10010030
Stübler A-S, Böhmker L, Juadjur A, Heinz V, Rauh C, Shpigelman A, Aganovic K. Matrix- and Technology-Dependent Stability and Bioaccessibility of Strawberry Anthocyanins during Storage. Antioxidants. 2021; 10(1):30. https://doi.org/10.3390/antiox10010030
Chicago/Turabian StyleStübler, Anna-Sophie, Lena Böhmker, Andreas Juadjur, Volker Heinz, Cornelia Rauh, Avi Shpigelman, and Kemal Aganovic. 2021. "Matrix- and Technology-Dependent Stability and Bioaccessibility of Strawberry Anthocyanins during Storage" Antioxidants 10, no. 1: 30. https://doi.org/10.3390/antiox10010030
APA StyleStübler, A. -S., Böhmker, L., Juadjur, A., Heinz, V., Rauh, C., Shpigelman, A., & Aganovic, K. (2021). Matrix- and Technology-Dependent Stability and Bioaccessibility of Strawberry Anthocyanins during Storage. Antioxidants, 10(1), 30. https://doi.org/10.3390/antiox10010030