Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Culture
2.3. Cell Viability
2.4. Zebrafish Husbandry and Embryos Collection
2.5. Animals and Experimental Diets
2.6. NO Measurement
2.7. ROS Measurement
2.8. RNA Extraction, RT-PCR and Real-Time qPCR
2.9. Enzyme-Linked Immunosorbent Measurement (ELISA)
2.10. Western Blotting
2.11. Tissue Histology
2.12. Statistical Analysis
3. Results
3.1. Cytotoxic Effect of BG In Vitro
3.2. Inhibitory Effect of BG on Nitric Oxide (NO) Production
3.3. Inhibitory Effect of BG on H2O2-Induced ROS Production
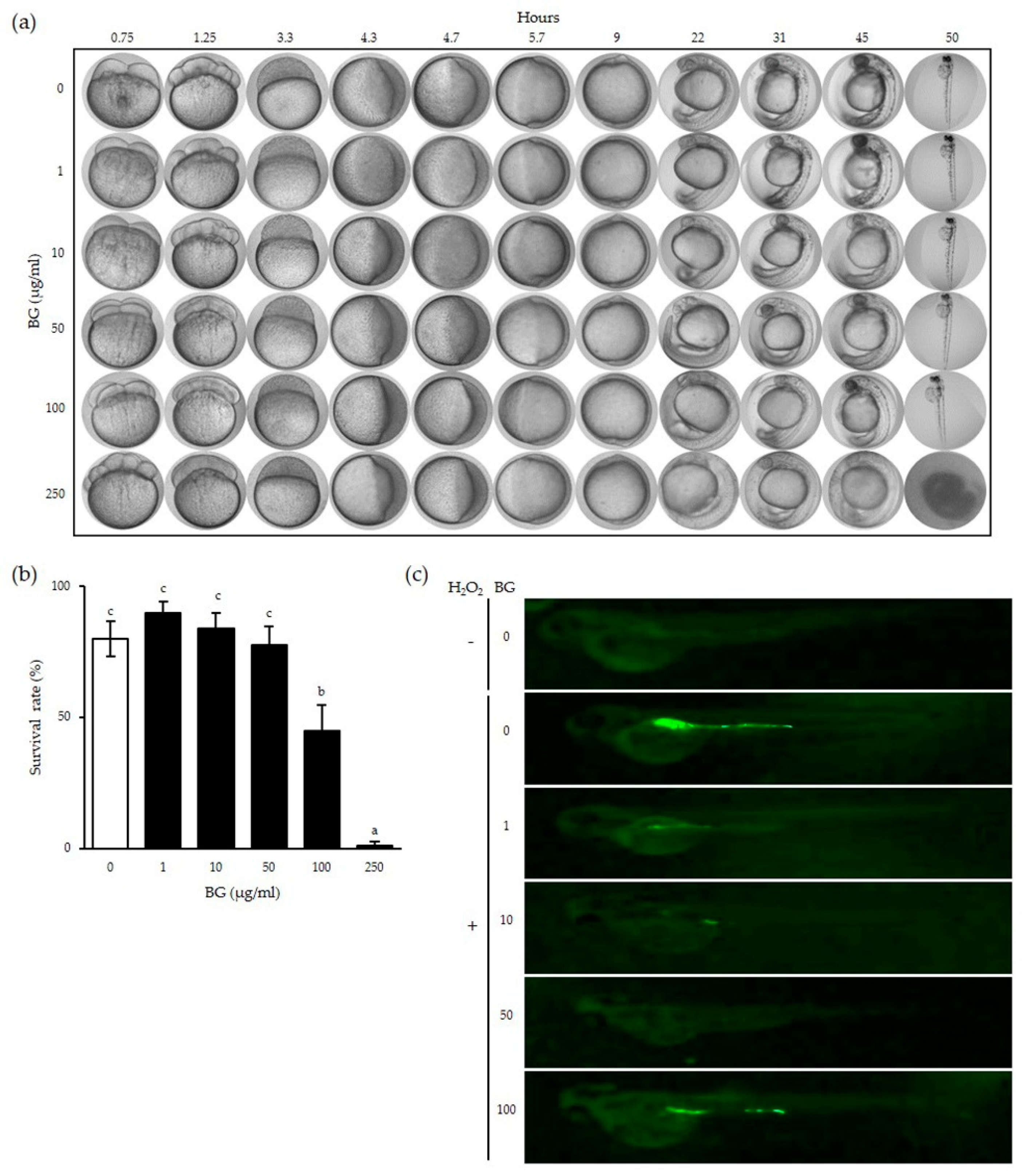
3.4. Cytotoxic and Inhibitory Effects of BG on ROS Production in Zebrafish Embryos
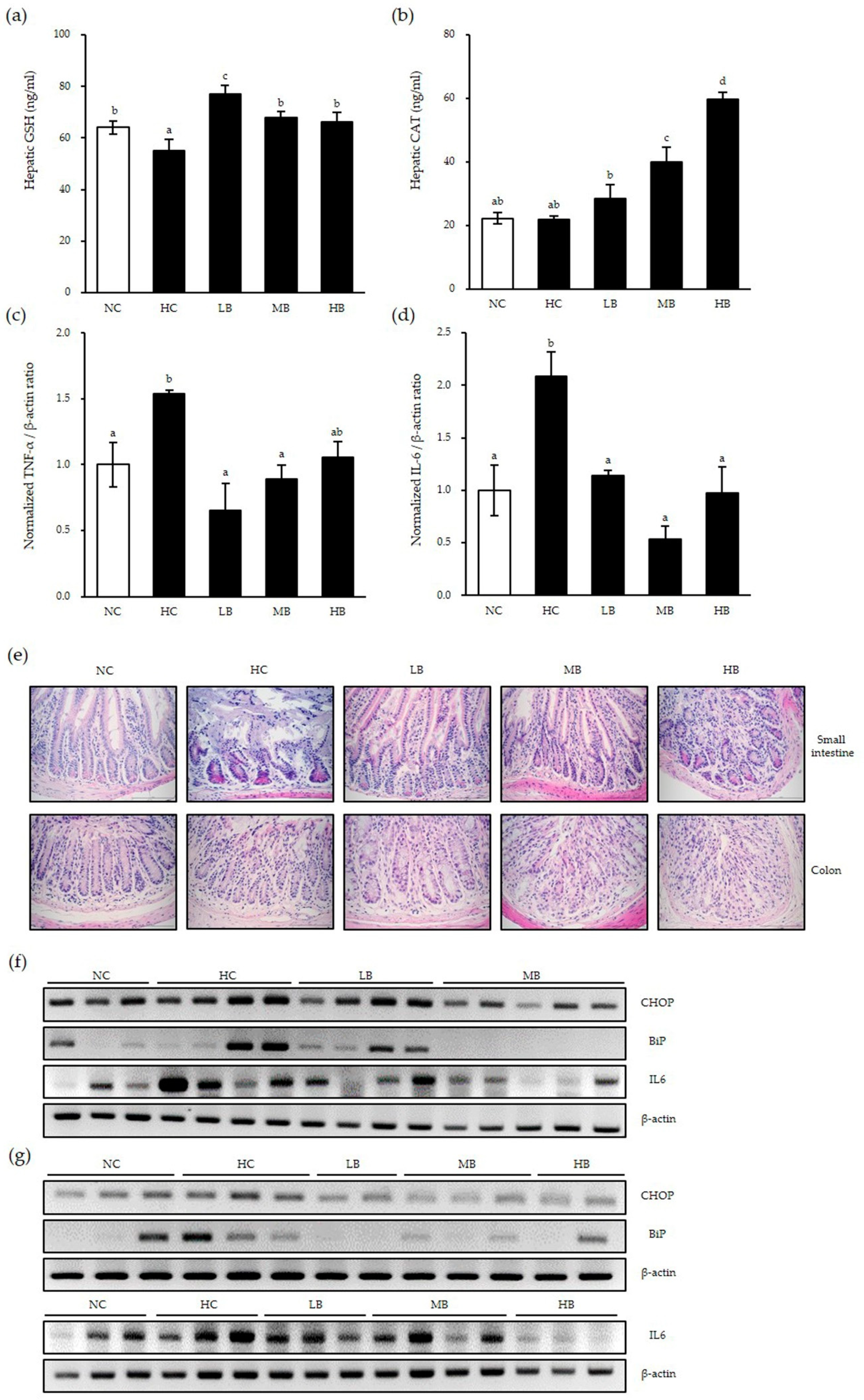
3.5. Inhibitory Effect of BG on a NAFLD Mouse Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bak, M.-J.; Jeong, J.-H.; Kang, H.-S.; Jin, K.-S.; Ok, S.; Jeong, W.-S. Cedrela sinensis leaves suppress oxidative stress and expressions of iNOS and COX-2 via MAPK signaling pathways in RAW 264.7 cells. J. Food Sci. Nutr. 2009, 14, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Yarlagadda, K.; Hassani, J.; Foote, I.P.; Markowitz, J.J. The role of nitric oxide in melanoma. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Nahrevanian, H.J. Involvement of nitric oxide and its up/down stream molecules in the immunity against parasitic infections. Braz. J. Infect. Dis. 2009, 13, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Jabs, T.J. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive Oxygen Species (ROS)––Induced genetic and epigenetic alterations in human carcinogenesis. Mutat. Res. 2011, 711, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J.N. Oxidative stress and aging: Catalase is a longevity determinant enzyme. Nature 2000, 408, 239. [Google Scholar] [CrossRef]
- Hancock, J.; Desikan, R.; Neill, S. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 29, 345–350. [Google Scholar] [CrossRef]
- Schreck, R.; Grassmann, R.; Fleckenstein, B.; Baeuerle, P.J. Antioxidants selectively suppress activation of NF-kappa B by human T-cell leukemia virus type I Tax protein. J. Virol. 1992, 66, 6288–6293. [Google Scholar] [CrossRef] [Green Version]
- Schreck, R.; Rieber, P.; Baeuerle, P.A. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar] [CrossRef]
- Schröder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. 2005, 569, 29–63. [Google Scholar] [CrossRef]
- Zeeshan, H.M.A.; Lee, G.H.; Kim, H.-R.; Chae, H.J. Endoplasmic reticulum stress and associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Cullinan, S.B.; Diehl, J.A. Coordination of ER and oxidative stress signaling: The PERK/Nrf2 signaling pathway. Int. J. Biochem. Cell Biol. 2006, 38, 317–332. [Google Scholar] [CrossRef]
- Guo, F.; Lin, E.A.; Liu, P.; Lin, J.; Liu, C.J. XBP1U inhibits the XBP1S-mediated upregulation of the iNOS gene expression in mammalian ER stress response. Cell Signal. 2010, 22, 1818–1828. [Google Scholar]
- Kim, J.-H. Cardiovascular diseases and Panax ginseng: A review on molecular mechanisms and medical applications. J. Ginseng Res. 2012, 36, 16. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.R.; Yun, B.S.; In, O.H.; Sung, C.K. Comparative study of Korean white, red, and black ginseng extract on cholinesterase inhibitory activity and cholinergic function. J. Ginseng Res. 2011, 35, 421. [Google Scholar] [CrossRef] [Green Version]
- Metwaly, A.M.; Lianlian, Z.; Luqi, H.; Deqiang, D.J. Black ginseng and its saponins: Preparation, phytochemistry and pharmacological effects. Molecules 2019, 24, 1856. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Yoo, J.-H.; Lee, Y.-S.; Park, E.-J.; Lee, H.-J. Ameliorative effects of black ginseng on nonalcoholic fatty liver disease in free fatty acid–induced HepG2 cells and high-fat/high-fructose diet-fed mice. J. Ginseng Res. 2020, 44, 350–361. [Google Scholar] [CrossRef]
- Park, S.-J.; Park, M.; Sharma, A.; Kim, K.; Lee, H.-J. Black Ginseng and Ginsenoside Rb1 Promote Browning by Inducing UCP1 Expression in 3T3-L1 and Primary White Adipocytes. Nutrients 2019, 11, 2747. [Google Scholar]
- Postlethwait, J.H.; Woods, I.G.; Ngo-Hazelett, P.; Yan, Y.-L.; Kelly, P.D.; Chu, F.; Huang, H.; Hill-Force, A.; Talbot, W.S. Zebrafish comparative genomics and the origins of vertebrate chromosomes. Genome Res. 2000, 10, 1890–1902. [Google Scholar] [CrossRef] [Green Version]
- Trede, N.S.; Langenau, D.M.; Traver, D.; Look, A.T.; Zon, L.I. The use of zebrafish to understand immunity. Immunity 2004, 20, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Vickers, N.J. Animal communication: When I’m calling you, will you answer too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef]
- Balakumar, M.; Raji, L.; Prabhu, D.; Sathishkumar, C.; Prabu, P.; Mohan, V.; Balasubramanyam, M.J. High-fructose diet is as detrimental as high-fat diet in the induction of insulin resistance and diabetes mediated by hepatic/pancreatic endoplasmic reticulum (ER) stress. Mol. Cell. Biochem. 2016, 423, 93–104. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Yang, A.Y.; Cheng, D.C.; Boyanapalli, S.S.-S.; Su, Z.-Y.; Khor, T.O.; Gao, S.; Wang, J.; Jiang, Z.-H.; Kong, A.-N. Pharmacodynamics of ginsenosides: Antioxidant activities, activation of Nrf2, and potential synergistic effects of combinations. Chem. Res. Toxicol. 2012, 25, 1574–1580. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.S.; Yamabe, N.; Kim, H.Y.; Okamoto, T.; Sei, Y.; Yokozawa, T.J. Increase in the free radical scavenging activities of American ginseng by heat processing and its safety evaluation. J. Ethnopharmacol. 2007, 113, 225–232. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Lima, J.L. Fluorescence probes used for detection of reactive oxygen species. Biochem. Biophys. Res. Commun. 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Caldefie-Chézet, F.; Walrand, S.; Moinard, C.; Tridon, A.; Chassagne, J.; Vasson, M.-P. Is the neutrophil reactive oxygen species production measured by luminol and lucigenin chemiluminescence intra or extracellular? Comparison with DCFH-DA flow cytometry and cytochrome c reduction. Clin. Chim. Acta 2002, 319, 9–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Diet | Number of Mice |
|---|---|---|
| NC | Normal drinking water | 10 |
| HC | 45% high-fat diet with 10% fructose in the drinking water | 9 |
| LB | 45% high-fat diet supplemented with 0.5% BG with 10% fructose in the drinking water | 9 |
| MB | 45% high-fat diet supplemented with 1% BG with 10% fructose in the drinking water | 9 |
| HB | 45% high-fat diet supplemented with 2% BG with 10% fructose in the drinking water | 9 |
| Total | 46 |
| Gene | Accession Number | Primer Sequence (5′–3′) | bp | |
|---|---|---|---|---|
| RT-PCR | BiP | NM_001163434.1 | Forward: CTG GGT ACA TTT GAT CTG ACT GG Reverse: GCA TCC TGG TGG CTT TCC AGC CAT TC | 398 |
| CHOP | NM_007837.4 | Forward: CAC ATC CCA AAG CCC TCG CTC TC Reverse: TCA TGC TTG GTG CAG GCT GAC CAT | 286 | |
| IL-6 | NM_001314054.1 | Forward: CCG GAG AGG AGA CTT CAC AG Reverse: GGA AAT TGG GGT AGG AAG GA | 421 | |
| β-actin | NM_007393.5 | Forward: TCT CCA GCA ACG AGG AGA AT Reverse: TGT GAT CTG AAA CCT GCT GC | 348 | |
| Real-time qPCR | TNF-α | NM_013693.3 | Forward: TGT CCC TTT CAC TCA CTG GC Reverse: CAT CTT TTG GGG GAG TGC CT | |
| IL-6 | NM_001314054.1 | Forward: GGG ACT GAT GCT GGT GAC AA Reverse: TCC ACG ATT TCC CAG AGA ACA | ||
| β-actin | NM_007393.5 | Forward: GACGTTGACATCCGTAAAG Reverse: CAGTAACAGTCCGCCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, M.-Y.; Lee, S.R.; Hwang, H.-J.; Yoon, J.-G.; Lee, H.-J.; Cho, J.A. Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway. Antioxidants 2021, 10, 62. https://doi.org/10.3390/antiox10010062
An M-Y, Lee SR, Hwang H-J, Yoon J-G, Lee H-J, Cho JA. Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway. Antioxidants. 2021; 10(1):62. https://doi.org/10.3390/antiox10010062
Chicago/Turabian StyleAn, Mi-Yeong, So Rok Lee, Hye-Jeong Hwang, Ju-Gyeong Yoon, Hae-Jeung Lee, and Jin Ah Cho. 2021. "Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway" Antioxidants 10, no. 1: 62. https://doi.org/10.3390/antiox10010062
APA StyleAn, M. -Y., Lee, S. R., Hwang, H. -J., Yoon, J. -G., Lee, H. -J., & Cho, J. A. (2021). Antioxidant and Anti-Inflammatory Effects of Korean Black Ginseng Extract through ER Stress Pathway. Antioxidants, 10(1), 62. https://doi.org/10.3390/antiox10010062