Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer



Abstract
:1. Introduction
1.1. ROS and Its Physiological Role
1.2. Oxidative Stress in Cancer
2. ROS and Associated Programmed Cell Death (PCD) Pathways
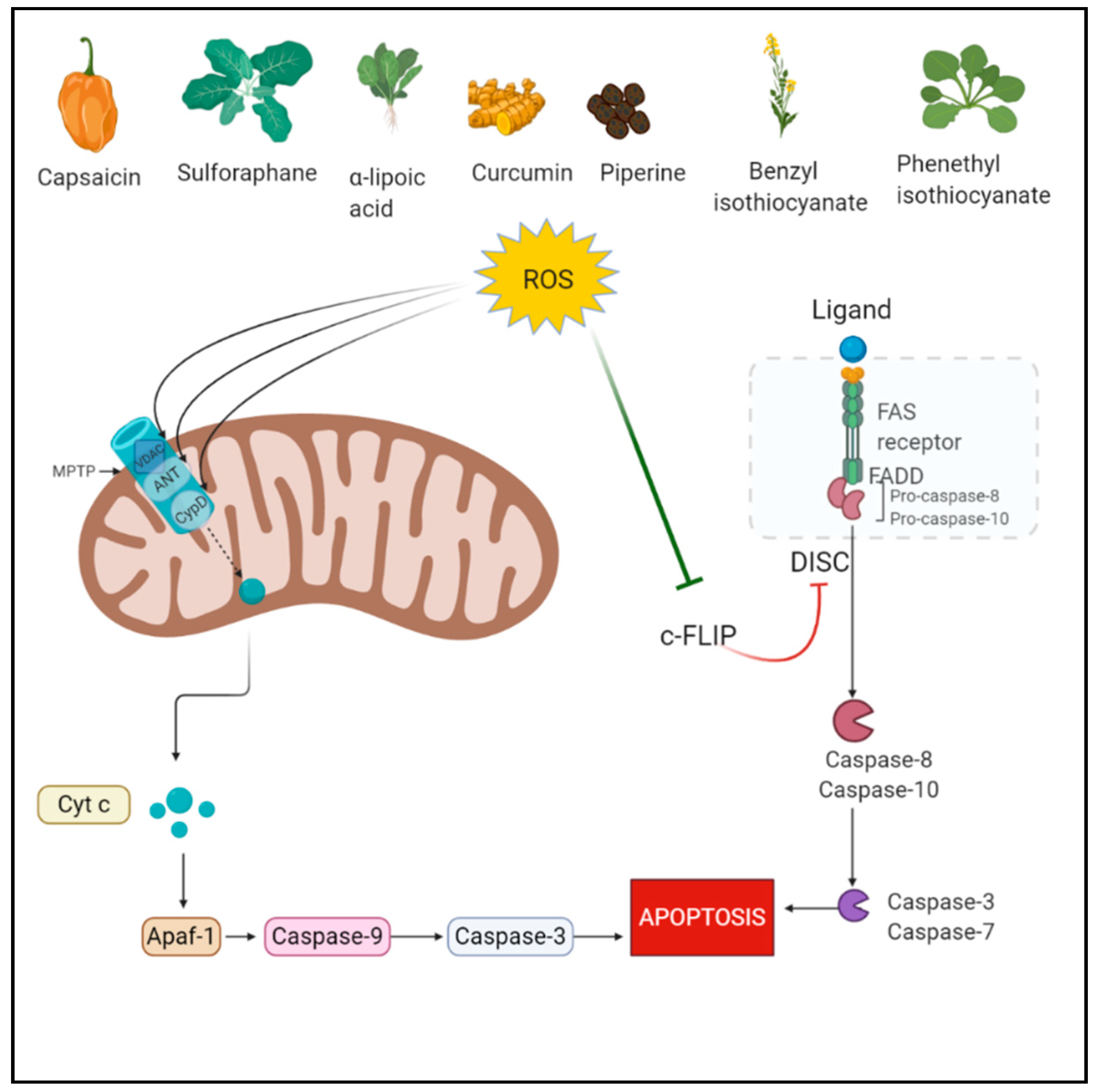
2.1. Apoptosis
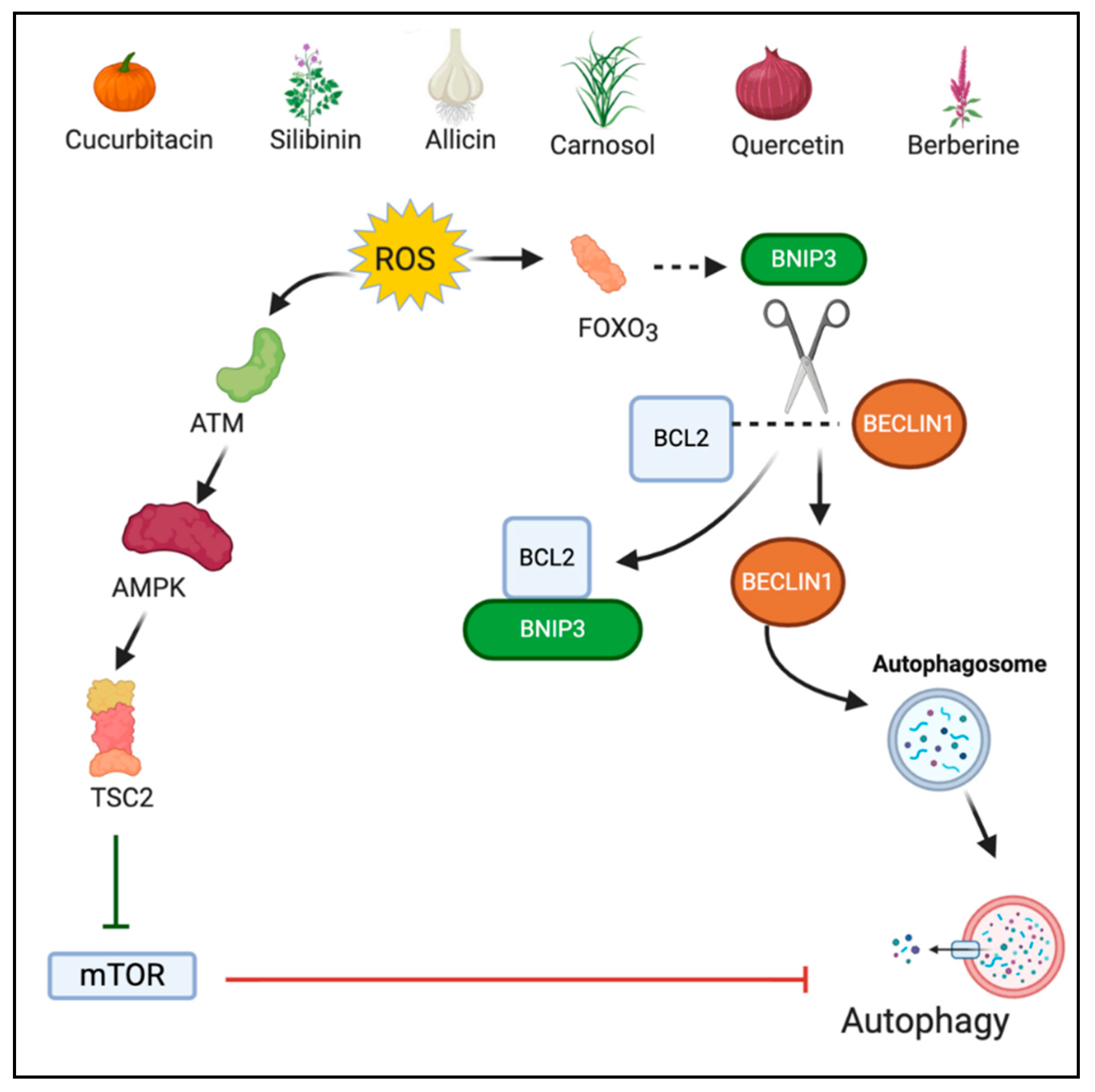
2.2. Autophagy
2.3. Ferroptosis
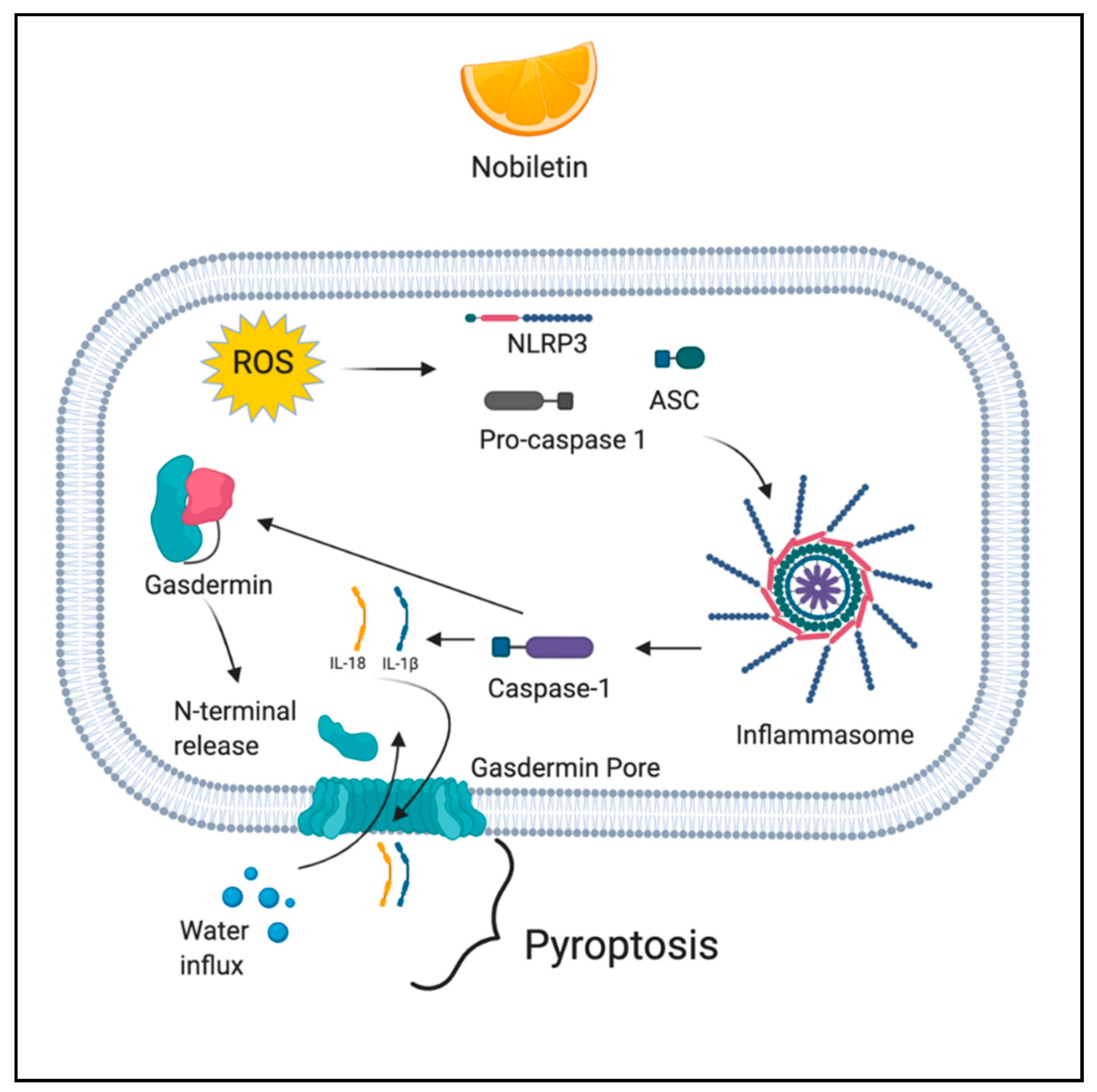
2.4. Pyroptosis
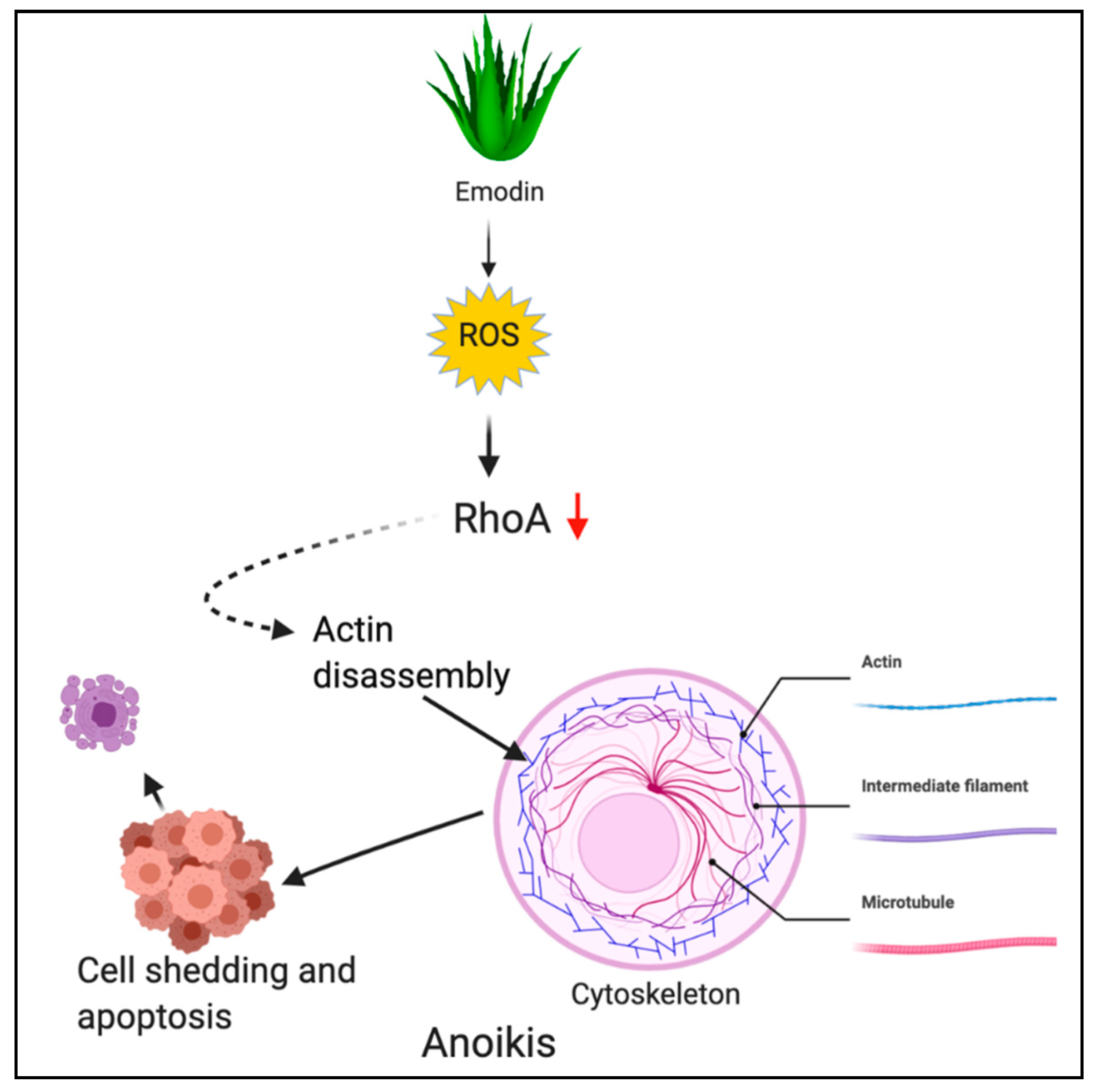
2.5. Anoikis
3. Phytochemicals: A Promising Role in ROS Mediated Cancer Cell Death
3.1. Phytochemicals Acting via ROS-Mediated Apoptosis
3.1.1. Capsaicin
3.1.2. Sulforaphane
3.1.3. α-Lipoic Acid
3.1.4. Benzyl Isothiocyanate (BITC)
3.1.5. Phenethyl Isothiocyanate (PEITC)
3.1.6. Piperine
3.1.7. Curcumin
3.2. Phytochemicals Acting via ROS-Mediated Ferroptosis
3.2.1. Withaferin A
3.2.2. Bromelain
3.2.3. Ruscogenin
3.2.4. Oridonin
3.3. Phytochemicals Acting via ROS-Mediated Autophagy
3.3.1. Cucurbitacin B
3.3.2. Silibinin
3.3.3. Allicin
3.3.4. Carnosol
3.3.5. Quercetin
3.3.6. Berberine
3.4. Phytochemicals Acting via ROS-Mediated Pyroptosis
Nobiletin
3.5. Phytochemicals Acting via ROS-Mediated Anoikis
Emodin
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell. Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, L.R.; Peng, J.; Chilian, W.M. Reactive Oxygen Species: The Good and the Bad. In Reactive Oxygen Species (ROS) in Living Cells; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Funato, Y. Regulation of intracellular signalling through cysteine oxidation by reactive oxygen species. J. Biochem. 2012, 151, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Oswald, M.C.W.; Garnham, N.; Sweeney, S.T.; Landgraf, M. Regulation of neuronal development and function by ROS. FEBS Lett. 2018, 592, 679–691. [Google Scholar] [CrossRef]
- Wilson, C.; Gonzalez-Billault, C. Regulation of cytoskeletal dynamics by redox signaling and oxidative stress: Implications for neuronal development and trafficking. Front. Cell. Neurosci. 2015, 9, 381. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, H.; Li, H.; Kalyanaraman, B.; Nicolosi, A.C.; Gutterman, D.D. Mitochondrial sources of H2O2 generation play a key role in flow-mediated dilation in human coronary resistance arteries. Circ. Res. 2003, 93, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushio-Fukai, M.; Alexander, R.W. Reactive oxygen species as mediators of angiogenesis signaling: Role of NAD(P)H oxidase. Mol. Cell. Biochem. 2004, 264, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Rocic, P.; Pung, Y.F.; Belmadani, S.; Carrao, A.C.; Ohanyan, V.; Chilian, W.M. Redox-dependent mechanisms in coronary collateral growth: The “redox window” hypothesis. Antioxid. Redox Signal. 2009, 11, 1961–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accardi, M.V.; Daniels, B.A.; Brown, P.M.; Fritschy, J.M.; Tyagarajan, S.K.; Bowie, D. Mitochondrial reactive oxygen species regulate the strength of inhibitory GABA-mediated synaptic transmission. Nat. Commun. 2014, 5, 3168. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C. Antimicrobial actions of reactive oxygen species. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupre-Crochet, S.; Erard, M.; Nubetae, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef]
- Kuiper, J.W.; Sun, C.; Magalhaes, M.A.; Glogauer, M. Rac regulates PtdInsP(3) signaling and the chemotactic compass through a redox-mediated feedback loop. Blood 2011, 118, 6164–6171. [Google Scholar] [CrossRef] [Green Version]
- Tavassolifar, M.J.; Vodjgani, M.; Salehi, Z.; Izad, M. The Influence of Reactive Oxygen Species in the Immune System and Pathogenesis of Multiple Sclerosis. Autoimmune Dis. 2020, 2020, 5793817. [Google Scholar] [CrossRef]
- Lee, K.; Esselman, W.J. cAMP potentiates H(2)O(2)-induced ERK1/2 phosphorylation without the requirement for MEK1/2 phosphorylation. Cell. Signal. 2001, 13, 645–652. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pr. Res. Clin. Obs. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, L.; Zhou, T.; Pannell, B.K.; Ziegler, A.C.; Best, T.M. Biological and physiological role of reactive oxygen species—The good, the bad and the ugly. Acta Physiol. 2015, 214, 329–348. [Google Scholar] [CrossRef] [PubMed]
- Leslie, N.R.; Bennett, D.; Lindsay, Y.E.; Stewart, H.; Gray, A.; Downes, C.P. Redox regulation of PI 3-kinase signalling via inactivation of PTEN. EMBO J. 2003, 22, 5501–5510. [Google Scholar] [CrossRef] [PubMed]
- Okoh, V.O.; Felty, Q.; Parkash, J.; Poppiti, R.; Roy, D. Reactive oxygen species via redox signaling to PI3K/AKT pathway contribute to the malignant growth of 4-hydroxy estradiol-transformed mammary epithelial cells. PLoS ONE 2013, 8, e54206. [Google Scholar] [CrossRef]
- Weinberg, F.; Hamanaka, R.; Wheaton, W.W.; Weinberg, S.; Joseph, J.; Lopez, M.; Kalyanaraman, B.; Mutlu, G.M.; Budinger, G.R.; Chandel, N.S. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc. Natl. Acad. Sci. USA 2010, 107, 8788–8793. [Google Scholar] [CrossRef] [Green Version]
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar] [CrossRef]
- Vafa, O.; Wade, M.; Kern, S.; Beeche, M.; Pandita, T.K.; Hampton, G.M.; Wahl, G.M. c-Myc can induce DNA damage, increase reactive oxygen species, and mitigate p53 function: A mechanism for oncogene-induced genetic instability. Mol. Cell 2002, 9, 1031–1044. [Google Scholar] [CrossRef]
- Cat, B.; Stuhlmann, D.; Steinbrenner, H.; Alili, L.; Holtkotter, O.; Sies, H.; Brenneisen, P. Enhancement of tumor invasion depends on transdifferentiation of skin fibroblasts mediated by reactive oxygen species. J. Cell Sci. 2006, 119, 2727–2738. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Subbaram, S.; Carrico, P.M.; Melendez, J.A. Redox-control of matrix metalloproteinase-1: A critical link between free radicals, matrix remodeling and degenerative disease. Respir. Physiol. Neurobiol. 2010, 174, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Ozben, T. Oxidative stress and apoptosis: Impact on cancer therapy. J. Pharm. Sci. 2007, 96, 2181–2196. [Google Scholar] [CrossRef]
- Kim, B.; Srivastava, S.K.; Kim, S.H. Caspase-9 as a therapeutic target for treating cancer. Expert Opin. Ther. Targets 2015, 19, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Madesh, M.; Hajnoczky, G. VDAC-dependent permeabilization of the outer mitochondrial membrane by superoxide induces rapid and massive cytochrome c release. J. Cell Biol. 2001, 155, 1003–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yi, J. Cancer cell killing via ROS: To increase or decrease, that is the question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef]
- Kumar, R.; Herbert, P.E.; Warrens, A.N. An introduction to death receptors in apoptosis. Int. J. Surg. 2005, 3, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Safa, A.R.; Day, T.W.; Wu, C.H. Cellular FLICE-like inhibitory protein (C-FLIP): A novel target for cancer therapy. Curr. Cancer Drug Targets 2008, 8, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Azad, N.; Kongkaneramit, L.; Chen, F.; Lu, Y.; Jiang, B.H.; Rojanasakul, Y. The Fas death signaling pathway connecting reactive oxygen species generation and FLICE inhibitory protein down-regulation. J. Immunol. 2008, 180, 3072–3080. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptations. Nat. Cell. Biol 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Ranjan, A.; Kaushik, I.; Srivastava, S.K. Pimozide Suppresses the Growth of Brain Tumors by Targeting STAT3-Mediated Autophagy. Cells 2020, 9, 2141. [Google Scholar] [CrossRef] [PubMed]
- Cordani, M.; Donadelli, M.; Strippoli, R.; Bazhin, A.V.; Sanchez-Alvarez, M. Interplay between ROS and Autophagy in Cancer and Aging: From Molecular Mechanisms to Novel Therapeutic Approaches. Oxid. Med. Cell. Longev. 2019, 2019, 8794612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.F.; Liu, H.; Luo, X.J.; Zhao, Z.; Zou, Z.Y.; Li, J.; Lin, X.J.; Liang, Y. The roles of reactive oxygen species (ROS) and autophagy in the survival and death of leukemia cells. Crit. Rev. Oncol. Hematol. 2017, 112, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Corradetti, M.N.; Inoki, K.; Guan, K.L. TSC2: Filling the GAP in the mTOR signaling pathway. Trends Biochem. Sci. 2004, 29, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.; Kim, J.; Walker, C.L. ATM engages the TSC2/mTORC1 signaling node to regulate autophagy. Autophagy 2010, 6, 672–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowolny, G.; Aucello, M.; Rizzuto, E.; Beccafico, S.; Mammucari, C.; Boncompagni, S.; Belia, S.; Wannenes, F.; Nicoletti, C.; Del Prete, Z.; et al. Skeletal muscle is a primary target of SOD1G93A-mediated toxicity. Cell Metab. 2008, 8, 425–436. [Google Scholar] [CrossRef]
- Burton, T.R.; Gibson, S.B. The role of Bcl-2 family member BNIP3 in cell death and disease: NIPping at the heels of cell death. Cell Death Differ. 2009, 16, 515–523. [Google Scholar] [CrossRef] [Green Version]
- White, E.; DiPaola, R.S. The double-edged sword of autophagy modulation in cancer. Clin. Cancer Res. 2009, 15, 5308–5316. [Google Scholar] [CrossRef] [Green Version]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Ribas, V.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Glutathione and mitochondria. Front. Pharm. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Tian, S.; Pan, Y.; Li, W.; Wang, Q.; Tang, Y.; Yu, T.; Wu, X.; Shi, Y.; Ma, P.; et al. Pyroptosis: A new frontier in cancer. Biomed. Pharm. 2020, 121, 109595. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, P.Y.; Bao, W.; Chen, S.J.; Wu, F.S.; Zhu, P.Y. Hydrogen inhibits endometrial cancer growth via a ROS/NLRP3/caspase-1/GSDMD-mediated pyroptotic pathway. BMC Cancer 2020, 20, 28. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, A.P. Anoikis. Cell Death Differ. 2005, 12 (Suppl. 2), 1473–1477. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Srivastava, S.K. Critical role of STAT3 in melanoma metastasis through anoikis resistance. Oncotarget 2014, 5, 7051–7064. [Google Scholar] [CrossRef] [Green Version]
- Fofaria, N.M.; Srivastava, S.K. STAT3 induces anoikis resistance, promotes cell invasion and metastatic potential in pancreatic cancer cells. Carcinogenesis 2015, 36, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Kandala, P.K.; Srivastava, S.K. Diindolylmethane-mediated Gli1 protein suppression induces anoikis in ovarian cancer cells in vitro and blocks tumor formation ability in vivo. J. Biol. Chem. 2012, 287, 28745–28754. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Gupta, N.; Fofaria, N.M.; Ranjan, A.; Srivastava, S.K. HER2-mediated GLI2 stabilization promotes anoikis resistance and metastasis of breast cancer cells. Cancer Lett. 2019, 442, 68–81. [Google Scholar] [CrossRef]
- Cai, J.; Niu, X.; Chen, Y.; Hu, Q.; Shi, G.; Wu, H.; Wang, J.; Yi, J. Emodin-induced generation of reactive oxygen species inhibits RhoA activation to sensitize gastric carcinoma cells to anoikis. Neoplasia 2008, 10, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boreddy, S.R.; Srivastava, S.K. Pancreatic cancer chemoprevention by phytochemicals. Cancer Lett. 2013, 334, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Cordell, G.A.; Araujo, O.E. Capsaicin: Identification, nomenclature, and pharmacotherapy. Ann. Pharm. 1993, 27, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.C.; Fofaria, N.M.; Gupta, P.; Ranjan, A.; Kim, S.H.; Srivastava, S.K. Inhibition of beta-catenin signaling suppresses pancreatic tumor growth by disrupting nuclear beta-catenin/TCF-1 complex: Critical role of STAT-3. Oncotarget 2015, 6, 11561–11574. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, K.C.; Fofaria, N.M.; Gupta, P.; Srivastava, S.K. CBP-mediated FOXO-1 acetylation inhibits pancreatic tumor growth by targeting SirT. Mol. Cancer Ther. 2014, 13, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, K.C.; Boreddy, S.R.; Srivastava, S.K. Role of mitochondrial electron transport chain complexes in capsaicin mediated oxidative stress leading to apoptosis in pancreatic cancer cells. PLoS ONE 2011, 6, e20151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Humphreys, I.; Sahu, R.P.; Shi, Y.; Srivastava, S.K. In vitro and in vivo induction of apoptosis by capsaicin in pancreatic cancer cells is mediated through ROS generation and mitochondrial death pathway. Apoptosis 2008, 13, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane: Translational research from laboratory bench to clinic. Nutr. Rev. 2013, 71, 709–726. [Google Scholar] [CrossRef]
- Singh, S.V.; Srivastava, S.K.; Choi, S.; Lew, K.L.; Antosiewicz, J.; Xiao, D.; Zeng, Y.; Watkins, S.C.; Johnson, C.S.; Trump, D.L.; et al. Sulforaphane-induced cell death in human prostate cancer cells is initiated by reactive oxygen species. J. Biol. Chem. 2005, 280, 19911–19924. [Google Scholar] [CrossRef] [Green Version]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Pibiri, M.; Sulas, P.; Camboni, T.; Leoni, V.P.; Simbula, G. alpha-Lipoic acid induces Endoplasmic Reticulum stress-mediated apoptosis in hepatoma cells. Sci. Rep. 2020, 10, 7139. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.C.; Srivastava, S.K.; Boreddy, S.R. Molecular Targets of Benzyl Isothiocyanates in Pancreatic Cancer; IntechOpen: London, UK, 2012. [Google Scholar]
- Boreddy, S.R.; Pramanik, K.C.; Srivastava, S.K. Pancreatic tumor suppression by benzyl isothiocyanate is associated with inhibition of PI3K/AKT/FOXO pathway. Clin. Cancer Res. 2011, 17, 1784–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boreddy, S.R.; Sahu, R.P.; Srivastava, S.K. Benzyl isothiocyanate suppresses pancreatic tumor angiogenesis and invasion by inhibiting HIF-alpha/VEGF/Rho-GTPases: Pivotal role of STAT-3. PLoS ONE 2011, 6, e25799. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.; Sahu, R.P.; Kandala, P.K.; Srivastava, S.K. Benzyl isothiocyanate-mediated inhibition of histone deacetylase leads to NF-kappaB turnoff in human pancreatic carcinoma cells. Mol. Cancer Ther. 2010, 9, 1596–1608. [Google Scholar] [CrossRef] [Green Version]
- Sahu, R.P.; Epperly, M.W.; Srivastava, S.K. Benzyl isothiocyanate sensitizes human pancreatic cancer cells to radiation therapy. Front. Biosci. 2009, 1, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Wicker, C.A.; Sahu, R.P.; Kulkarni-Datar, K.; Srivastava, S.K.; Brown, T.L. BITC Sensitizes Pancreatic Adenocarcinomas to TRAIL-induced Apoptosis. Cancer Growth Metastasis 2010, 2009, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Sahu, R.P.; Zhang, R.; Batra, S.; Shi, Y.; Srivastava, S.K. Benzyl isothiocyanate-mediated generation of reactive oxygen species causes cell cycle arrest and induces apoptosis via activation of MAPK in human pancreatic cancer cells. Carcinogenesis 2009, 30, 1744–1753. [Google Scholar] [CrossRef] [Green Version]
- Loganathan, S.; Kandala, P.K.; Gupta, P.; Srivastava, S.K. Inhibition of EGFR-AKT axis results in the suppression of ovarian tumors in vitro and in preclinical mouse model. PLoS ONE 2012, 7, e43577. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Oh, J.; Kim, M.; Jin, E.J. Bromelain effectively suppresses Kras-mutant colorectal cancer by stimulating ferroptosis. Anim. Cells Syst. 2018, 22, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Wright, S.E.; Srivastava, S.K. PEITC treatment suppresses myeloid derived tumor suppressor cells to inhibit breast tumor growth. Oncoimmunology 2015, 4, e981449. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Kim, B.; Kim, S.H.; Srivastava, S.K. Molecular targets of isothiocyanates in cancer: Recent advances. Mol. Nutr. Food Res. 2014, 58, 1685–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Wright, S.E.; Kim, S.H.; Srivastava, S.K. Phenethyl isothiocyanate: A comprehensive review of anti-cancer mechanisms. Biochim. Biophys. Acta 2014, 1846, 405–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Adkins, C.; Lockman, P.; Srivastava, S.K. Metastasis of Breast Tumor Cells to Brain Is Suppressed by Phenethyl Isothiocyanate in a Novel In Vivo Metastasis Model. PLoS ONE 2013, 8, e67278. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Srivastava, S.K. Antitumor activity of phenethyl isothiocyanate in HER2-positive breast cancer models. BMC Med. 2012, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Gorgani, L.; Mohammadi, M.; Najafpour, G.D.; Nikzad, M. Piperine—The Bioactive Compound of Black Pepper: From Isolation to Medicinal Formulations. Compr. Rev. Food Sci. Food 2017, 16, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, H.; Chikara, S.; Reindl, K.M. Piperlongumine induces pancreatic cancer cell death by enhancing reactive oxygen species and DNA damage. Toxicol. Rep. 2014, 1, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fofaria, N.M.; Qhattal, H.S.; Liu, X.; Srivastava, S.K. Nanoemulsion formulations for anti-cancer agent piplartine—Characterization, toxicological, pharmacokinetics and efficacy studies. Int. J. Pharm. 2016, 498, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Fofaria, N.M.; Kim, S.H.; Srivastava, S.K. Piperine causes G1 phase cell cycle arrest and apoptosis in melanoma cells through checkpoint kinase-1 activation. PLoS ONE 2014, 9, e94298. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Sahu, R.P.; Batra, S.; Srivastava, S.K. Activation of ATM/Chk1 by curcumin causes cell cycle arrest and apoptosis in human pancreatic cancer cells. Br. J. Cancer 2009, 100, 1425–1433. [Google Scholar] [CrossRef] [Green Version]
- Khaket, T.P.; Singh, M.P.; Khan, I.; Kang, S.C. In vitro and in vivo studies on potentiation of curcumin-induced lysosomal-dependent apoptosis upon silencing of cathepsin C in colorectal cancer cells. Pharm. Res. 2020, 161, 105156. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, W.; Sabbe, L.; Kaileh, M.; Haegeman, G.; Heyninck, K. Molecular insight in the multifunctional activities of Withaferin, A. Biochem. Pharm. 2012, 84, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Hassannia, B.; Wiernicki, B.; Ingold, I.; Qu, F.; Van Herck, S.; Tyurina, Y.Y.; Bayir, H.; Abhari, B.A.; Angeli, J.P.F.; Choi, S.M.; et al. Nano-targeted induction of dual ferroptotic mechanisms eradicates high-risk neuroblastoma. J. Clin. Investig. 2018, 128, 3341–3355. [Google Scholar] [CrossRef] [PubMed]
- Pavan, R.; Jain, S.; Shraddha; Kumar, A. Properties and therapeutic application of bromelain: A review. Biotechnol. Res. Int. 2012, 2012, 976203. [Google Scholar] [CrossRef] [Green Version]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Masullo, M.; Pizza, C.; Piacente, S. Ruscus Genus: A Rich Source of Bioactive Steroidal Saponins. Planta Med. 2016, 82, 1513–1524. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Xiang, X.; Li, J.; Deng, J.; Fang, Z.; Zhang, L.; Xiong, J. Ruscogenin induces ferroptosis in pancreatic cancer cells. Oncol. Rep. 2020, 43, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Owona, B.A.; Schluesener, H.J. Molecular Insight in the Multifunctional Effects of Oridonin. Drugs R D 2015, 15, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, N.; Zhou, Y.; Wang, K.; Sun, Y.; Yan, H.; Han, W.; Wang, X.; Wei, B.; Ke, Y.; et al. Oridonin induces ferroptosis by inhibiting gamma-glutamyl cycle in TE1 cells. Phytother. Res. 2020. [Google Scholar] [CrossRef]
- Fujii, J.; Homma, T.; Kobayashi, S. Ferroptosis caused by cysteine insufficiency and oxidative insult. Free Radic. Res. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (Review). Int. J. Oncol. 2018, 52, 19–37. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Park, K.A.; Byun, H.S.; Won, M.; Jeon, J.; Lee, Y.; Seok, J.H.; Choi, S.W.; Lee, S.H.; et al. Cucurbitacin induces autophagy through mitochondrial ROS production which counteracts to limit caspase-dependent apoptosis. Autophagy 2012, 8, 559–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.; Sha, T.; Guo, J.; Li, W.; Lu, J.; Chen, X. Cucurbitacin B induces DNA damage and autophagy mediated by reactive oxygen species (ROS) in MCF-7 breast cancer cells. J. Nat. Med. 2015, 69, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.J.; Li, Q.S.; Xia, M.Y.; Tashiro, S.; Onodera, S.; Ikejima, T. Silibinin activated p53 and induced autophagic death in human fibrosarcoma HT1080 cells via reactive oxygen species-p38 and c-Jun N-terminal kinase pathways. Biol. Pharm. Bull. 2011, 34, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Wang, W.; Jin, X.; Wang, Z.; Ji, Z.; Meng, G. Silibinin, a natural flavonoid, induces autophagy via ROS-dependent mitochondrial dysfunction and loss of ATP involving BNIP3 in human MCF7 breast cancer cells. Oncol Rep 2015, 33, 2711–2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, L.D.; Hunsaker, S.M. Allicin Bioavailability and Bioequivalence from Garlic Supplements and Garlic Foods. Nutrients 2018, 10, 812. [Google Scholar] [CrossRef] [Green Version]
- Pandey, N.; Tyagi, G.; Kaur, P.; Pradhan, S.; Rajam, M.V.; Srivastava, T. Allicin Overcomes Hypoxia Mediated Cisplatin Resistance in Lung Cancer Cells through ROS Mediated Cell Death Pathway and by Suppressing Hypoxia Inducible Factors. Cell Physiol. Biochem. 2020, 54, 748–766. [Google Scholar] [CrossRef]
- Chu, Y.L.; Ho, C.T.; Chung, J.G.; Raghu, R.; Lo, Y.C.; Sheen, L.Y. Allicin induces anti-human liver cancer cells through the p53 gene modulating apoptosis and autophagy. J. Agric. Food Chem. 2013, 61, 9839–9848. [Google Scholar] [CrossRef]
- Johnson, J.J. Carnosol: A promising anti-cancer and anti-inflammatory agent. Cancer Lett. 2011, 305, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.M.; Kim, D.H.; Chun, K.S. Carnosol induces apoptotic cell death through ROS-dependent inactivation of STAT3 in human melanoma G361 cells. Appl. Biol. Chem. 2019, 62, 1–11. [Google Scholar] [CrossRef]
- Al Dhaheri, Y.; Attoub, S.; Ramadan, G.; Arafat, K.; Bajbouj, K.; Karuvantevida, N.; AbuQamar, S.; Eid, A.; Iratni, R. Carnosol induces ROS-mediated beclin1-independent autophagy and apoptosis in triple negative breast cancer. PLoS ONE 2014, 9, e109630. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, T.; Cano, C.E.; Grasso, D.; Garcia, M.N.; Sandi, M.J.; Calvo, E.L.; Dagorn, J.C.; Lomberk, G.; Urrutia, R.; Goruppi, S.; et al. Nupr1-aurora kinase A pathway provides protection against metabolic stress-mediated autophagic-associated cell death. Clin. Cancer Res. 2012, 18, 5234–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Zeng, W.; Ouyang, W.; Xu, Q.; Chen, J.; Wang, B.; Zhang, X. Quercetin induced NUPR1-dependent autophagic cell death by disturbing reactive oxygen species homeostasis in osteosarcoma cells. J. Clin. Biochem. Nutr. 2020, 67, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neag, M.A.; Mocan, A.; Echeverria, J.; Pop, R.M.; Bocsan, C.I.; Crisan, G.; Buzoianu, A.D. Berberine: Botanical Occurrence, Traditional Uses, Extraction Methods, and Relevance in Cardiovascular, Metabolic, Hepatic, and Renal Disorders. Front. Pharmacol. 2018, 9, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, T.Z.; de Moraes, F.R.; Tedesco, A.C.; Arni, R.K.; Rahal, P.; Calmon, M.F. Berberine associated photodynamic therapy promotes autophagy and apoptosis via ROS generation in renal carcinoma cells. Biomed. Pharmacother. 2020, 123, 109794. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, L.; Shi, W.; Liu, H.; Yang, J.; Yuan, X.; Wu, L. The Multifunctional Effects of Nobiletin and Its Metabolites In Vivo and In Vitro. Evid Based Complement. Altern. Med. 2016, 2016, 2918796. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Chen, J.; Mao, L.; Guo, Y.; Hao, Y.; Deng, Y.; Han, X.; Li, Q.; Liao, W.; Yuan, M. Nobiletin Triggers Reactive Oxygen Species-Mediated Pyroptosis through Regulating Autophagy in Ovarian Cancer Cells. J. Agric. Food Chem. 2020, 68, 1326–1336. [Google Scholar] [CrossRef]
- He, W.T.; Wan, H.; Hu, L.; Chen, P.; Wang, X.; Huang, Z.; Yang, Z.H.; Zhong, C.Q.; Han, J. Gasdermin D is an executor of pyroptosis and required for interleukin-1beta secretion. Cell Res. 2015, 25, 1285–1298. [Google Scholar] [CrossRef]
- Huang, Q.; Lu, G.; Shen, H.M.; Chung, M.C.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef]
- Suresh, D.; Srinivasan, K. Tissue distribution & elimination of capsaicin, piperine & curcumin following oral intake in rats. Indian J. Med. Res. 2010, 131, 682–691. [Google Scholar]
- Chaiyasit, K.; Khovidhunkit, W.; Wittayalertpanya, S. Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. J. Med. Assoc. Thail. 2009, 92, 108–113. [Google Scholar]
- Sestili, P.; Fimognari, C. Cytotoxic and Antitumor Activity of Sulforaphane: The Role of Reactive Oxygen Species. Biomed. Res. Int. 2015, 2015, 402386. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.D.; Hsu, A.; Yu, Z.; Dashwood, R.H.; Ho, E. Differential effects of sulforaphane on histone deacetylases, cell cycle arrest and apoptosis in normal prostate cells versus hyperplastic and cancerous prostate cells. Mol. Nutr. Food Res. 2011, 55, 999–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, K.; Rizwanullah, M.; Kohli, K. Development and optimization of sulforaphane-loaded nanostructured lipid carriers by the Box-Behnken design for improved oral efficacy against cancer: In vitro, ex vivo and in vivo assessments. Artif. Cells Nanomed. Biotechnol. 2018, 46, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.Y.; Liu, H.L.; Stern, J.S.; Yu, P.Z.; Liu, S.L. Alpha-lipoic acid induces apoptosis in hepatoma cells via the PTEN/Akt pathway. FEBS Lett 2008, 582, 1667–1671. [Google Scholar] [CrossRef] [Green Version]
- Teichert, J.; Hermann, R.; Ruus, P.; Preiss, R. Plasma kinetics, metabolism, and urinary excretion of alpha-lipoic acid following oral administration in healthy volunteers. J. Clin. Pharmacol. 2003, 43, 1257–1267. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Ranjan, A.; Kim, S.H.; Srivastava, S.K. Mechanisms of the Anticancer Effects of Isothiocyanates. Enzymes 2015, 37, 111–137. [Google Scholar] [CrossRef]
- Xiao, D.; Vogel, V.; Singh, S.V. Benzyl isothiocyanate-induced apoptosis in human breast cancer cells is initiated by reactive oxygen species and regulated by Bax and Bak. Mol. Cancer Ther. 2006, 5, 2931–2945. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.K.; Singh, S.V. Cell cycle arrest, apoptosis induction and inhibition of nuclear factor kappa B activation in anti-proliferative activity of benzyl isothiocyanate against human pancreatic cancer cells. Carcinogenesis 2004, 25, 1701–1709. [Google Scholar] [CrossRef]
- Trachootham, D.; Zhou, Y.; Zhang, H.; Demizu, Y.; Chen, Z.; Pelicano, H.; Chiao, P.J.; Achanta, G.; Arlinghaus, R.B.; Liu, J.; et al. Selective killing of oncogenically transformed cells through a ROS-mediated mechanism by beta-phenylethyl isothiocyanate. Cancer Cell 2006, 10, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Kuo, Y.; Morris, M.E. Pharmacokinetics of dietary phenethyl isothiocyanate in rats. Pharm. Res. 2005, 22, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, J.; Liao, H.; Li, L.; Pan, L. Piperine induces apoptosis of lung cancer A549 cells via p53-dependent mitochondrial signaling pathway. Tumour Biol. 2014, 35, 3305–3310. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.M.; Yu, Z.X.; Ferrans, V.J.; Lowenstein, R.A.; Finkel, T. Reactive oxygen species are downstream mediators of p53-dependent apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 11848–11852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumpa-ngern, P. Pharmacokinetics of piperine following single dose administration of benjakul formulation in healthy Thai subjects. Afr. J. Pharm. Pharmacol. 2013, 7, 560–566. [Google Scholar] [CrossRef]
- Kim, B.; Kim, H.S.; Jung, E.J.; Lee, J.Y.; Tsang, B.K.; Lim, J.M.; Song, Y.S. Curcumin induces ER stress-mediated apoptosis through selective generation of reactive oxygen species in cervical cancer cells. Mol. Carcinog. 2016, 55, 918–928. [Google Scholar] [CrossRef]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [Green Version]
- Thaiparambil, J.T.; Bender, L.; Ganesh, T.; Kline, E.; Patel, P.; Liu, Y.; Tighiouart, M.; Vertino, P.M.; Harvey, R.D.; Garcia, A.; et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Int. J. Cancer 2011, 129, 2744–2755. [Google Scholar] [CrossRef]
- Committee for Medicinal Products for Veterinary Use. European Public MRL Assessment Report (EPMAR) Bromelain; European Medicines Agency: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Ji, P.Y.; Li, Z.W.; Yang, Q.; Wu, R. Rapid determination of ruscogenin in rat plasma with application to pharmacokinetic study. J. Chromatogr. B 2015, 985, 71–74. [Google Scholar] [CrossRef]
- Song, M.; Liu, X.; Liu, K.; Zhao, R.; Huang, H.; Shi, Y.; Zhang, M.; Zhou, S.; Xie, H.; Chen, H.; et al. Targeting AKT with Oridonin Inhibits Growth of Esophageal Squamous Cell Carcinoma In Vitro and Patient-Derived Xenografts In Vivo. Mol. Cancer Ther. 2018, 17, 1540–1553. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, N.; Li, N.; Fan, X.; Li, Y. Influence of verapamil on the pharmacokinetics of oridonin in rats. Pharm. Biol. 2019, 57, 787–791. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Islam, M.S.; Tian, J.; Lui, V.W.; Xiao, D. Inactivation of ATP citrate lyase by Cucurbitacin B: A bioactive compound from cucumber, inhibits prostate cancer growth. Cancer Lett. 2014, 349, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Hunsakunachai, N.; Nuengchamnong, N.; Jiratchariyakul, W.; Kummalue, T.; Khemawoot, P. Pharmacokinetics of cucurbitacin B from Trichosanthes cucumerina L. in rats. BMC Complement. Altern. Med. 2019, 19, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoh, C.; Boocock, D.; Marczylo, T.; Singh, R.; Berry, D.P.; Dennison, A.R.; Hemingway, D.; Miller, A.; West, K.; Euden, S.; et al. Pilot study of oral silibinin, a putative chemopreventive agent, in colorectal cancer patients: Silibinin levels in plasma, colorectum, and liver and their pharmacodynamic consequences. Clin. Cancer Res. 2006, 12, 2944–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.S.; Ma, J.Y.; Feng, R.; Ma, C.; Chen, W.J.; Sun, Y.P.; Fu, J.; Huang, M.; He, C.Y.; Shou, J.W.; et al. Tissue distribution of berberine and its metabolites after oral administration in rats. PLoS ONE 2013, 8, e77969. [Google Scholar] [CrossRef]
- Chen, J.; Chen, A.Y.; Huang, H.; Ye, X.; Rollyson, W.D.; Perry, H.E.; Brown, K.C.; Rojanasakul, Y.; Rankin, G.O.; Dasgupta, P.; et al. The flavonoid nobiletin inhibits tumor growth and angiogenesis of ovarian cancers via the Akt pathway. Int. J. Oncol. 2015, 46, 2629–2638. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Wahajuddin; Tewari, D.; Patel, K.; Jain, G.K. Permeability determination and pharmacokinetic study of nobiletin in rat plasma and brain by validated high-performance liquid chromatography method. Fitoterapia 2011, 82, 1206–1214. [Google Scholar] [CrossRef]
- Dumit, V.I.; Zerbes, R.M.; Kaeser-Pebernard, S.; Rackiewicz, M.; Wall, M.T.; Gretzmeier, C.; Kuttner, V.; van der Laan, M.; Braun, R.J.; Dengjel, J. Respiratory status determines the effect of emodin on cell viability. Oncotarget 2017, 8, 37478–37490. [Google Scholar] [CrossRef]
- Wu, W.; Yan, R.; Yao, M.; Zhan, Y.; Wang, Y. Pharmacokinetics of anthraquinones in rat plasma after oral administration of a rhubarb extract. Biomed. Chromatogr. 2014, 28, 564–572. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Phytochemical | PCD Type | Effective Anti-Cancer Concentration (In Vitro) | Dose Administered (In Vivo) | Pharmacokinetic Data | TttCell Lines/Subjects | Outcome/Comments | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Capsaicin | Apoptosis | 150 µM | 2.5 mg/kg | Not stated | Animal subject—Mice In vitro study Pancreatic cancer cells | Apoptosis was observed in AsPC-1 and BXPC3 cell lines at 150 µM concentration, while no effect were observed in normal (HPDE-6) cells. In an animal study, an oral dose 2.5 mg/kg was effective in suppressing tumor growth. Human equivalent dose—0.202 mg/kg. | [67,68] |
| 2 | Capsaicin (Pharmacokinetic study) | - | - | 5 g | Cmax: 2.47 ± 0.13 ng/mL | Human subjects | The oral bioavailability of capsaicin is effective for anti-diabetic effect, but plasma level is less for its anti-cancer activity. | [123,124] |
| 3 | Sulforaphane | Apoptosis | 5–10 µM | - | - | - | Various studies report anticancer activity in a range of 5–10 µM which is not attainable through dietary intake. A study in normal vs. cancerous prostate cells revealed selective cytotoxicity of sulforaphane in cancerous cells. No effect was observed in normal cells at a dose range of 0–15 µM. | [125,126] |
| 4 | Sulforaphane (Pharmacokinetic study) | - | - | 30 mg/kg | Cmax: 772.8 ± 54.36 ng/mL | Animal subject—Rats | Nano structured lipid formulation of sulforaphane increased its bioavailability in rat plasma wherein a dose close to effective anticancer level was achieved. | [127] |
| 5 | α-lipoic acid | Apoptosis | 500 µM | - | - | In vitro study hepatoma cell line | ROS-mediated apoptosis was observed at 500 µM. However, in another study, no effect was observed in normal L02 liver cells at 5 mM concentration. | [72,128] |
| 6 | α-lipoic acid (Pharmacokinetic study) | - | - | 600 mg per day | Cmax: 8–52 nM | Human subjects | Plasma levels achieved in plasma following oral administration of racemic α-lipoic acid is much less than required for its anti-cancer activity. | [129] |
| 7 | BITC | Apoptosis | - | - | - | In vitro study Pancreatic cancer cells | BITC caused ROS generation in a concentration dependent manner starting from 2.5 to 20 µM in MIAPaCa-2 and Capan-2 cell lines | [79,130] |
| 8 | BITC | Apoptosis | <2.5–5 µM | - | - | In vitro study Breast cancer cells and normal mammary epithelial cell line | BITC induced ROS-mediated apoptosis in MDA-MB-231 and MCF-7 at IC50 value of <2.5 µm and 5 µM respectively at 24-h time point. However, the IC50 was 20 µM in normal mammary epithelial cell line MCF-10A. | [131] |
| 9 | BITC | Apoptosis | 8 µM | - | - | In vitro study Pancreatic cancer cells | An IC50 of 8 µM was observed when BXPC3 cells were treated with BITC. | [132] |
| 10 | BITC (Pharmacokinetic study) | - | - | 12 µM/day | Cmax: 7.5 µM in tumor | Animal subject—Mice | BXPC3 cells were subcutaneously implanted and a 43% tumor inhibition was observed with a concentration near IC50 value achieved in plasma. | [74] |
| 11 | PEITC | Apoptosis | <5 µM | - | - | In vitro study Breast cancer cells | PEITC caused ROS-mediated apoptosis in MDA-MB-231 and MCF-7 cell lines. | [86] |
| 12 | PEITC | Apoptosis | 5–10 µM | - | - | In vitro study normal ovarian epithelial cell line | Ovarian epithelial cell line T72 was transfected to over express Ras oncogene to form oncogenic T72Ras cell line. Following PEITC treatment, the transformed cells showed higher sensitivity to ROS as compared to normal cells. Thus, showing selective activity of PEITC. | [133] |
| 13 | PEITC (Pharmacokinetic study) | - | - | 10–100 µmol/kg | Cmax: 9–42 µM | Animal subjects—Rats | A considerably high level of PEITC was achieved after oral administration of PEITC than required for its anti-cancer activity. | [134] |
| 14 | Piperine | Apoptosis | 136–137 µM | - | - | In vitro study Melanoma cell lines | Piperine induced RPS mediated apoptosis in SKMEL-28 and B16-F0 cells at 136 µM and 137 µM respectively at 72-h time point. | [90] |
| 15 | Piperine | Apoptosis | 132 µg/mL | - | - | In vitro study Lung cancer cells and normal lung fibroblasts | Piperine induced apoptotic cell death via p53 dependent mitochondrial pathway in A549 cancer cell line while no significant cytotoxicity was observed in normal WI38 human lung fibroblasts. ROS are considered to be downstream effectors of p53 mediated apoptosis. | [135,136] |
| 16 | Piperine (Pharmacokinetic study) | - | - | 200 mg tablets (Benjakul formulation) | Cmax: 1.078 µg/mL | Human subjects | Piperine is a major component of a traditional Thai medication called Benjakul. A dose of 200mg resulted in a Cmax level of 1.078 µg/mL which is significantly lower than that required for anticancer effect. | [137] |
| 17 | Curcumin | Apoptosis | 25 µM | 5 mg/kg | - | In vitro study Colorectal cancer cells Animal subjects—Mice | Curcumin induced ROS-mediated apoptotic cell death in HCT-116 cell line at 25 µM concentration within 48hh. Curcumin showed considerable tumor inhibition in in vivo xenograft model following administration of 5 mg/kg I.P dose. | [93] |
| 18 | Curcumin | Apoptosis | 20 µM | - | - | In vitro study Cervical cancer cells and normal cervical cells | Curcumin induced ROS-mediated apoptosis in cervical cancer cell lines C33A, CaSki, HeLa, and ME180 cells at approximately 20 µM in 48-h time period, while it did not induce significant toxicity in normal counterparts until 40 µM concentration. | [138] |
| 19 | Curcumin (Pharmacokinetic study) | - | - | Various doses (2–10 g) | Cmax: 1–3200 ng/mL | Healthy human volunteers and patients (cancer, Alzheimer’s disease etc.) | The serum levels of curcumin achieved by oral administration of crude curcumin are much lower than required for anti-cancer activity. However, a 10–15-fold increase in plasma levels was observed when curcumin was formulated as nanoparticle or combined with piperine, lecithin, etc. | [139] |
| 20 | Withaferin A | Ferroptosis | 5–10 µM | 4 mg/kg | Not stated | In vitro study Neuroblastoma cell lines Animal subjects— Mice | Crude Withaferin A and its nano particle formulation for tumor targeting showed ROS-mediated ferroptosis at 10 µM in neuroblastoma cell lines IMR-32, SK-N-SH, Kelly, NB69, and CHP-134 within 4–8 h. Tumor regression was mediated through lipid peroxidation. | [95] |
| 21 | Withaferin A | Apoptotic | 1–50 µM | 4 mg/kg | Cmax: 1.8 µM | In vitro study Breast cancer cell lines Animal subject—Mice | Withaferin A displayed cytotoxicity in breast carcinoma cell lines MDA-MB 231, H1299, T47D, MCF-7, LN686, as well as normal fibroblast cell line COS-7 in a wide range of 1–50 µM at 72-h time point. The IC50 was lesser than normal fibroblasts in majority of the cancer cell lines. | [140] |
| 22 | Bromelain | Ferroptosis | - | 3 mg/kg | Not stated | In vitro study Colon carcinoma cell lines and normal colon cell line. Animal subjects—Mice | Bromelain inhibited proliferation of in Kras mutant colon cancer cell lines HCT-116 and DLD-1 at 50 µg/mL, while it induced significant ferroptosis in cancer cell lines at a concentration of 5 µm when combined with Erastin as compared to normal colon cells CCD1co. Bromelain increased survival rate in treatment group as compared to vehicle group. | [81] |
| 23 | Bromelain (Pharmacokinetic study) | - | - | 143 mg/kg | Cmax: Very low (specific value not stated) | Human subjects | Bromelain showed very low (ng/mL) plasma levels following oral administration at 143 mg/kg body weight. | [141] |
| 24 | Ruscogenin | Ferroptosis | 7.3–28.19 µM | 5 or 10 mg/kg | Not stated | In vitro study Pancreatic cancer cells Animal subjects—Mice | Ruscogenin induced significant ferroptotic cell death in pancreatic cancer cell lines BxPC-3, SW1990, PANC-1, and ASPC-1 cells, as compared to normal pancreatic cell line HPDE-6-C7 wherein not IC50 was detected at 72-h time point in HPDE6-C7 cells. Additionally, it inhibited pancreatic cancer growth in vivo. | [99] |
| 25 | Ruscogenin (Pharmacokinetic study) | - | - | 8 mg/kg | Cmax: 504.50 ± 63.47 ng/mL (Mean value) | Animal subjects—Rats | Following IV administration of Ruscogenin in rats, a plasma level between 2–1000 ng/mL was observed which roughly translates to 2.35 µM, a level much lower than required for anticancer effect. | [142] |
| 26 | Oridonin | Ferroptosis | 26.93 µM | - | - | In vitro study Esophageal cancer cells | Oridonin induced ferroptotic cell death in esophageal cancer cell line TE1 at 27 µM within 24-h time point. | [101] |
| 27 | Oridonin | Apoptosis | 2.5–10 µM | - | - | In vitro study Esophageal cancer cell lines | Oridonin induced cellular death in human esophageal cancer cell lines (KYSE70, KYSE410, and KYSE450) at 2.5–10 µM concentration, while it showed 40% cell death in normal esophageal cell line SHEE at 40 µM, a significantly higher concentration. | [143] |
| 28 | Oridonin (Pharmacokinetic study) | - | - | 20 mg/kg | Cmax: 146.9 ± 10.17 ng/mL | Animal subjects—Rats | A concentration of approximately 150 ng/mL translating to 412 nM was achieved. Combination with verapamil increased the Cmax level of oridonin to 194 ± 10 ng/mL, still a lower level for anti-cancer efficacy. | [144] |
| 29 | Cucurbitacin B | Autophagy | Approximately 1 µM | - | - | In vitro study Cervical cancer cell line and breast cancer cell lines | Cucurbitacin B induced caspase independent autophagic cell death in HeLa cells at 1 µM. Additionally, it induced autophagy and increased ROS levels in MCF-7 at 200 nM. Autophagic cell death in normal counterparts was not studied. | [104] |
| 30 | Cucurbitacin B | Apoptosis | 0.2 µM | - | - | In vitro study Prostate cancer cell lines | Cucurbitacin B induced cell death and ROS production in human prostate cancer cell lines LNCaP and PC-3 at 0.2 µM while no significant cell death or ROS production was observed in normal prostate cell line PrEC. | [145] |
| 31 | Cucurbitacin B (Pharmacokinetic study) | - | - | 2–4 mg/kg | Cmax: 9–31 µg/L | Animal subjects—Rats | Following an oral administration of 2–4 mg/kg, a significantly low plasma level was achieved in rat plasma than required for anti-cancer activity. | [146] |
| 32 | Silibinin | Autophagy | 40 µM | - | - | In vitro study Fibrosarcoma cells | Silibinin induced autophagy in human fibrosarcoma cell lines HT1080 at 40 µM within a period of 4 h. Cellular death was concluded to be an autophagy mediated apoptosis process. Effect on normal cells was not studied. | [106] |
| 33 | Silibinin (Pharmacokinetic study) | - | - | 360, 720 and 1440 mg | Cmax: 0.4, 1.4 and 4 ± 5.3 µM respectively | Human Subjects—Colorectal cancer patients | Silibinin formulated capsules showed considerably high levels required to exert its anti-tumor effect. | [147] |
| 34 | Allicin | Autophagy and apoptosis | 15–30 µg/mL | - | - | In vitro study Non-small cell lung cancer cell lines | Allicin induced both autophagy and apoptosis in human lung cancer cell lines A549 and NCI-H460. | [109] |
| 35 | Carnosol | Autophagy | <25 µM | - | - | In vitro study Breast cancer cell lines | Carnosol induced ROS-mediated autophagy in triple negative breast cancer cell line MDA-MB-231. Effect on normal cell counterparts was not studied. | [113] |
| 36 | Quercetin | Autophagy | 200 µM | 100 mg/kg | - | In vitro study Osteosarcoma cell line Animal subjects—Mice | Quercetin induced ROS-mediated autophagic cell death in human osteosarcoma cell line MG-63 and also inhibited tumor growth in mice model. | [116] |
| 37 | Berberine | Autophagy | - | - | - | In vitro study Renal carcinoma cell lines and normal kidney cell line. | Berberine induced autophagic cell death in 786-O, ACHN cell lines via its photosensitizer activity when combined with laser. Additionally, it induced cell death in normal kidney cell line HK-2 at when combined with laser. Treatment with Berberine alone failed to induce cell death >20% in cancer cells, while it induced cell death >20% in HK-2 normal cells at 40 µM within 48 h. | [118] |
| 38 | Berberine (Pharmacokinetic study) | - | - | 200 mg/kg | Cmax: 25.85 ± 7.34 µg/L | Animal subject— Rats | A plasma level of 25.85 ± 7.34 µg/L equivalent to 76 µM was achieved. | [148] |
| 39 | Nobiletin | Pyroptosis | 34.85–35.31 µM | - | - | In vitro study Ovarian cancer cell lines | Nobiletin induced cytotoxicity at concentration of 35 µM but no data regarding its effect on normal cell lines were shown. | [120] |
| 40 | Nobiletin | Apoptosis | 40–80 µM | 100 mg/kg | - | In vitro study Ovarian cancer cell lines OVCAR-3 and A2780 Animal subject— Mice | Nobiletin exhibited cytotoxicity in ovarian cancer cell lines at 40–80 µM while the IC50 for normal ovarian cell line was around 160 µM. Additionally, significant tumor growth inhibition was observed in athymic nude mice model at a dose of 100 mg/kg. | [149] |
| 41 | Nobiletin (Pharmacokinetic study) | - | - | 50 mg/kg | Cmax: 1.78 µg/mL in plasma | Animal subject—Rats | The plasma level achieved after oral administration is 1.78 µg/mL which correlates to approximately 4–5 µM. | [150] |
| 42 | Emodin | Anoikis | 10 µM | - | - | In vitro study Gastric cancer cell lines | Significant difference observed in RhoA expression between cancer and normal cell lines. | [61] |
| 43 | Emodin | Apoptosis | 70 µM | - | - | In vitro study Cancer cell lines isolated from breast, lung, colon, and cervix carcinomas, normal human fibroblasts, and normal human keratinocytes | No effect observed on normal cell lines after 48-h treatment while cytotoxicity was observed in cancer cell lines. | [151] |
| 44 | Emodin (Pharmacokinetic study) | - | - | 4.5 mg/kg | Cmax: 0.2 ± 0.1 µM | Animal subject—Rats | The concentration of free emodin achieved in rat plasma after oral administration of rhubarb extract was found to be much lower than required for its anti-cancer activity. | [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaikwad, S.; Srivastava, S.K. Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants 2021, 10, 83. https://doi.org/10.3390/antiox10010083
Gaikwad S, Srivastava SK. Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants. 2021; 10(1):83. https://doi.org/10.3390/antiox10010083
Chicago/Turabian StyleGaikwad, Shreyas, and Sanjay K. Srivastava. 2021. "Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer" Antioxidants 10, no. 1: 83. https://doi.org/10.3390/antiox10010083
APA StyleGaikwad, S., & Srivastava, S. K. (2021). Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants, 10(1), 83. https://doi.org/10.3390/antiox10010083