
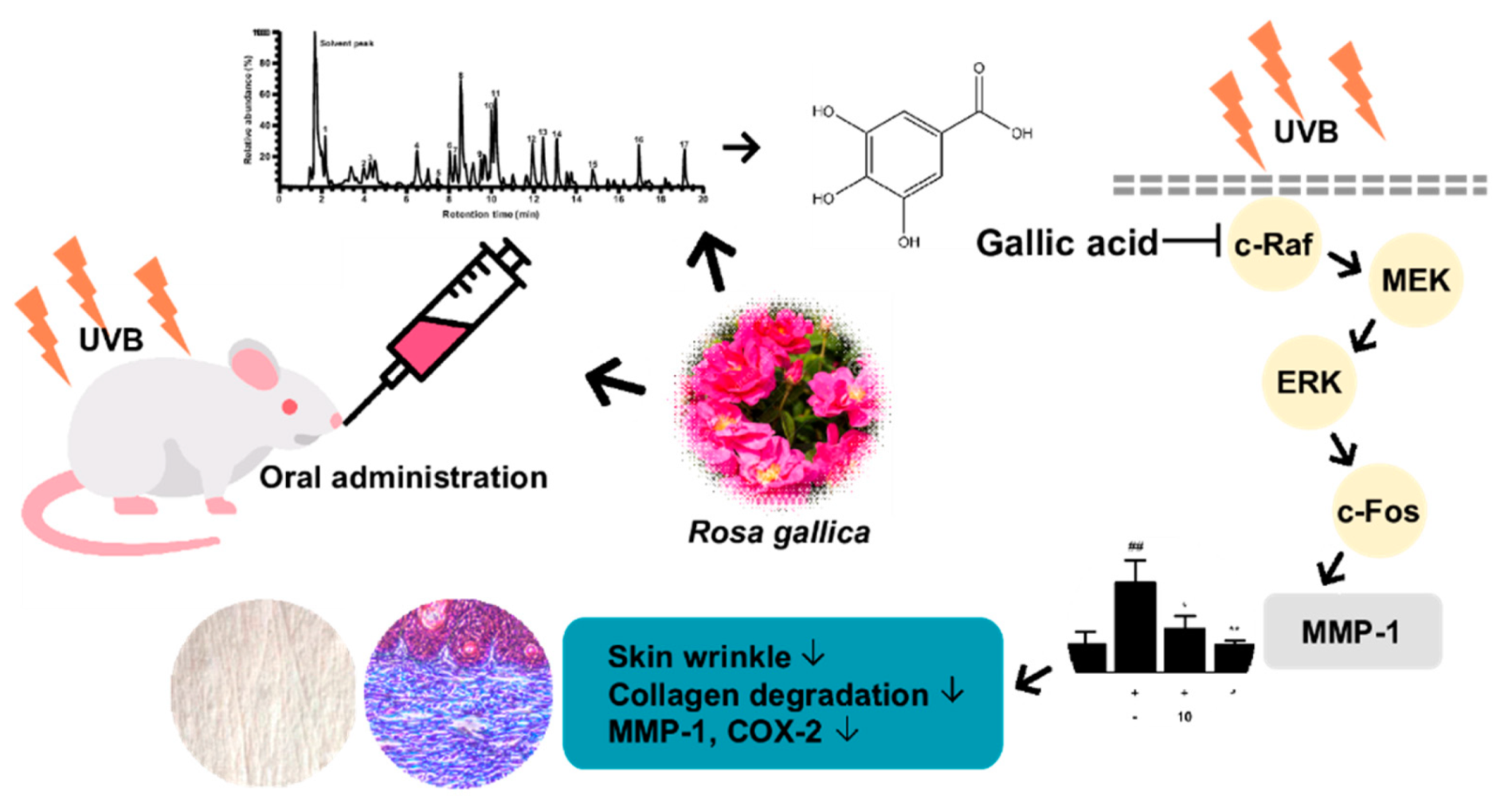
Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Rosa gallica Extract Preparation
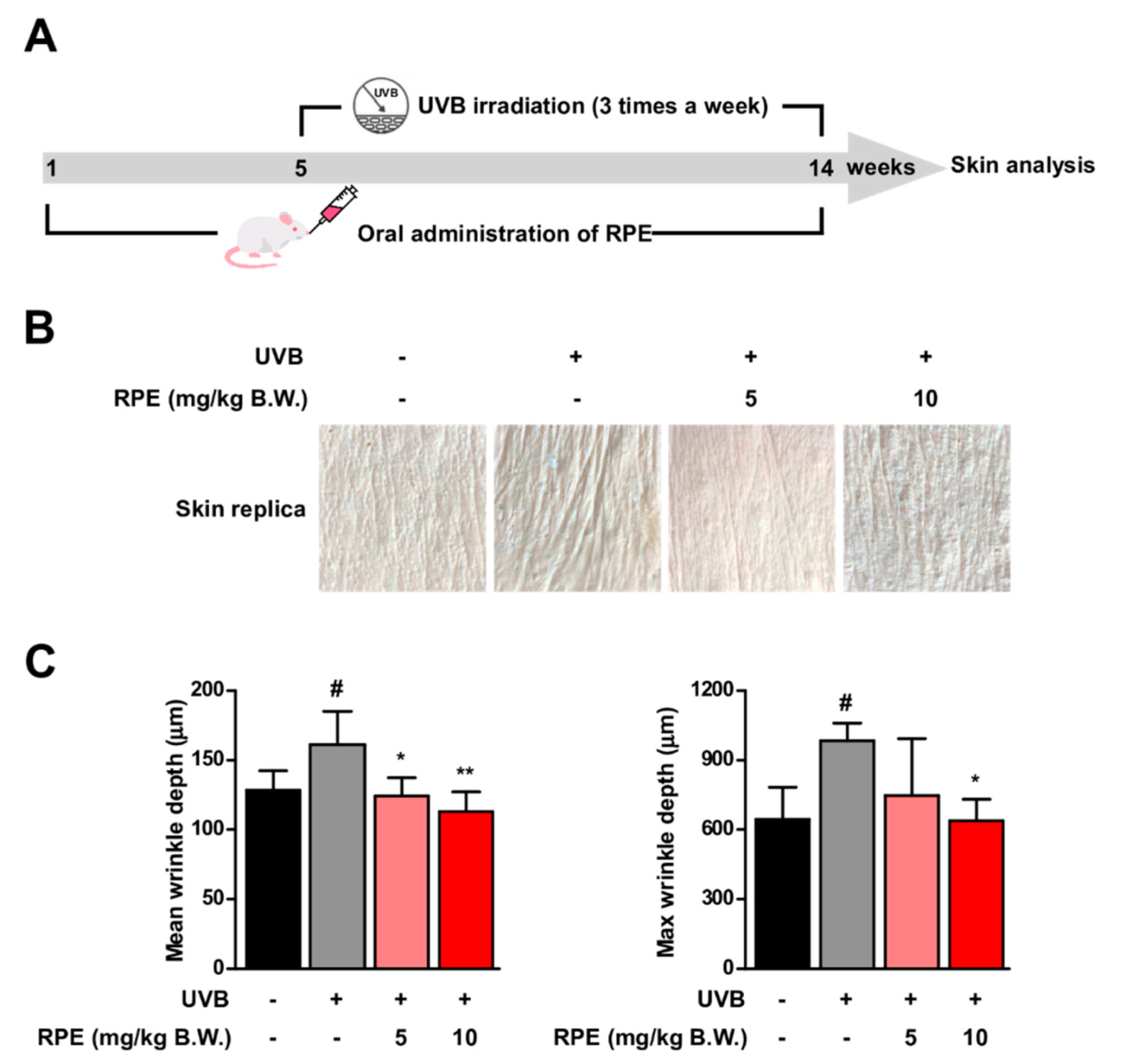
2.3. Experimental Animals and Treatments
2.4. Wrinkle Measurement
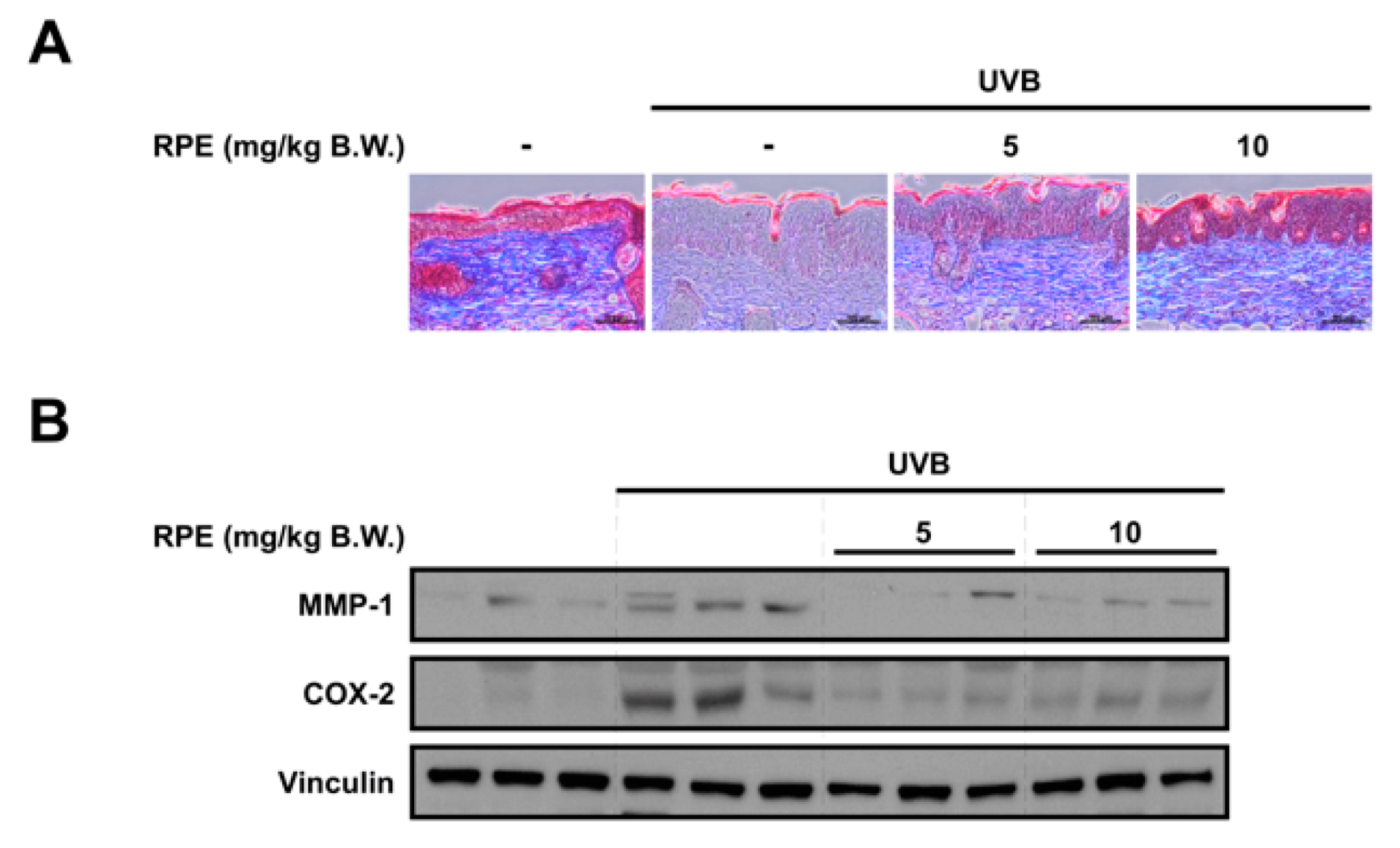
2.5. Masson’s Trichrome Staining
2.6. Immunoblot
2.7. UHPLC–LTQ–Orbitrap/MS/MS Conditions
2.8. Cell Culture and UVB Irradiation
2.9. SRB Staining
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Reconstructed 3D Human Skin Model
2.12. Collagen Staining in Reconstructed 3D Human Skin Model
2.13. Statistical Analysis
3. Results
3.1. Oral Consumption of Rosa gallica Petal Extract (RPE) Suppresses UVB−Mediated Skin Wrinkle In Vivo
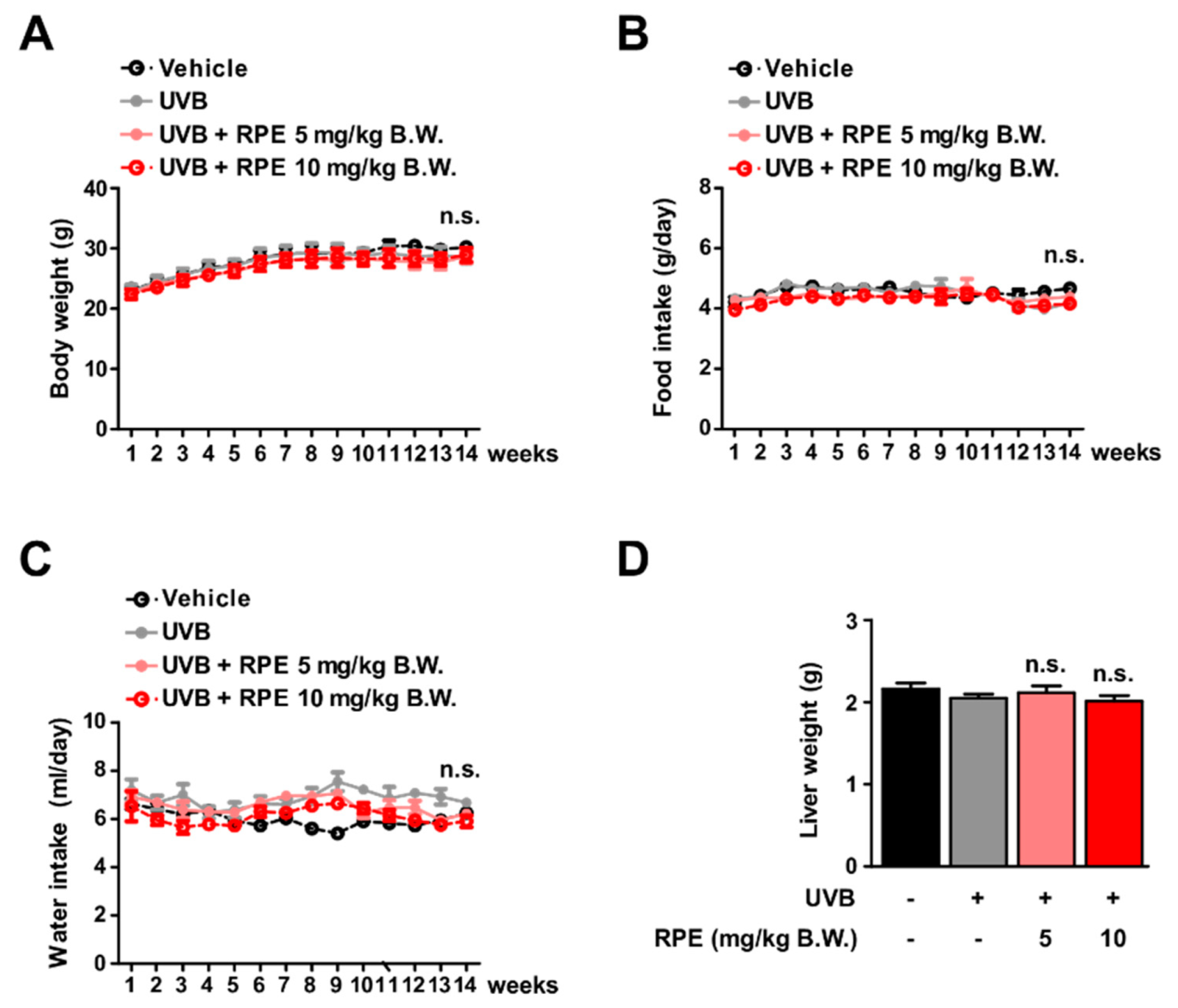
3.2. RPE Does Not Show Side Effects In Vivo
3.3. RPE Inhibits Collagen Degradation and Blocks UVB−Mediated Biomarker of Skin Aging
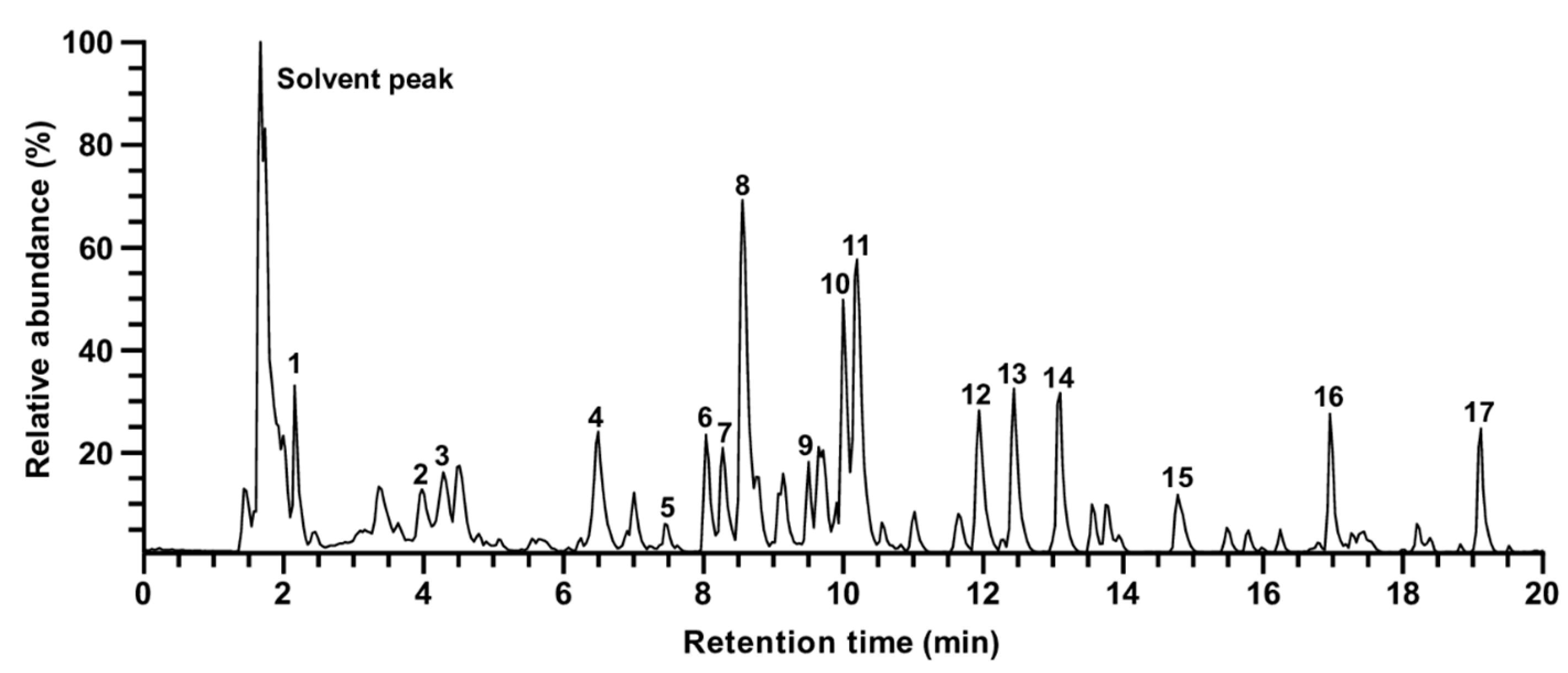
3.4. Identification of Compounds in Rosa gallica
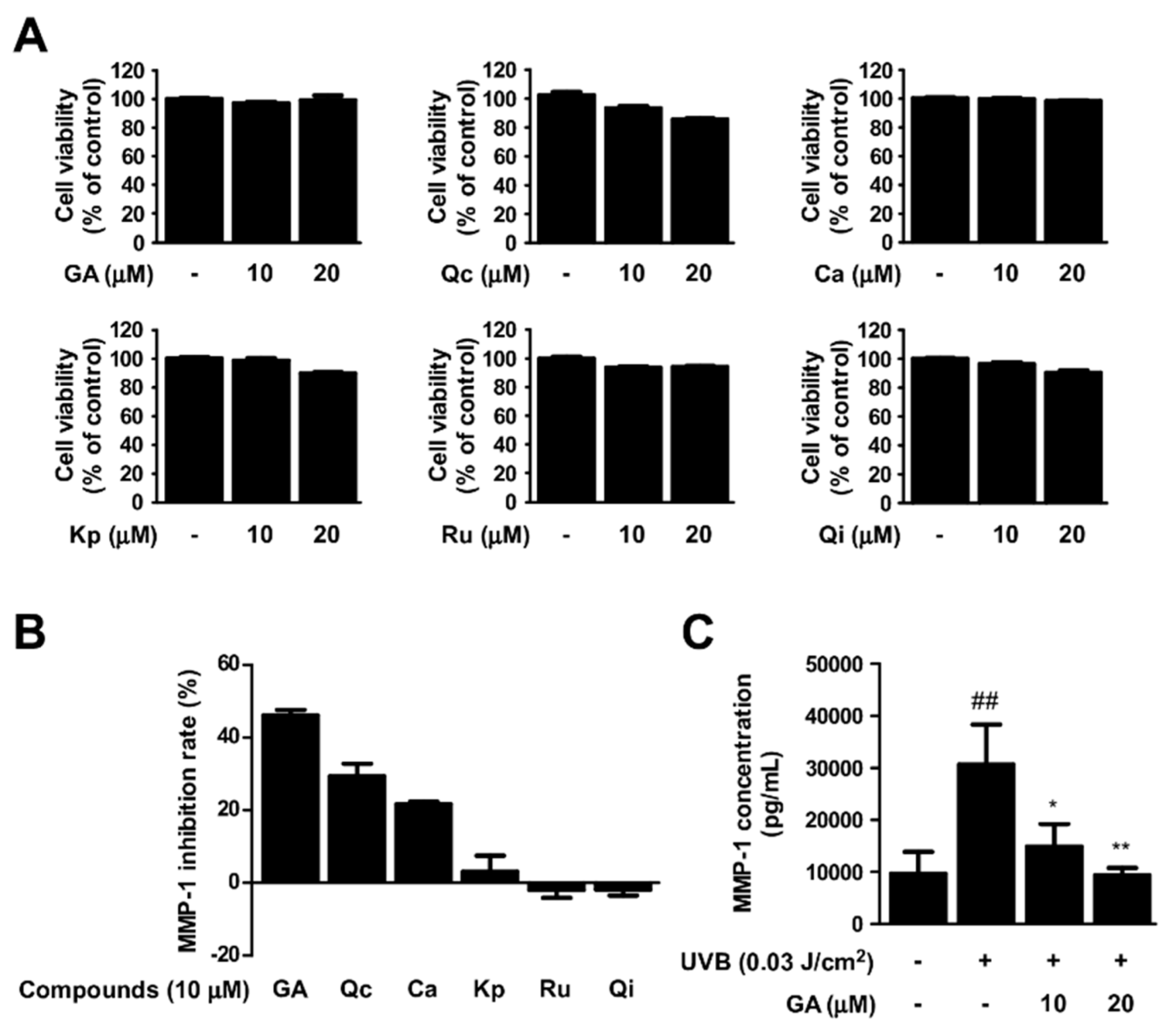
3.5. Gallic Acid Is a Major Active Compound of Rosa gallica in Preventing Skin Aging
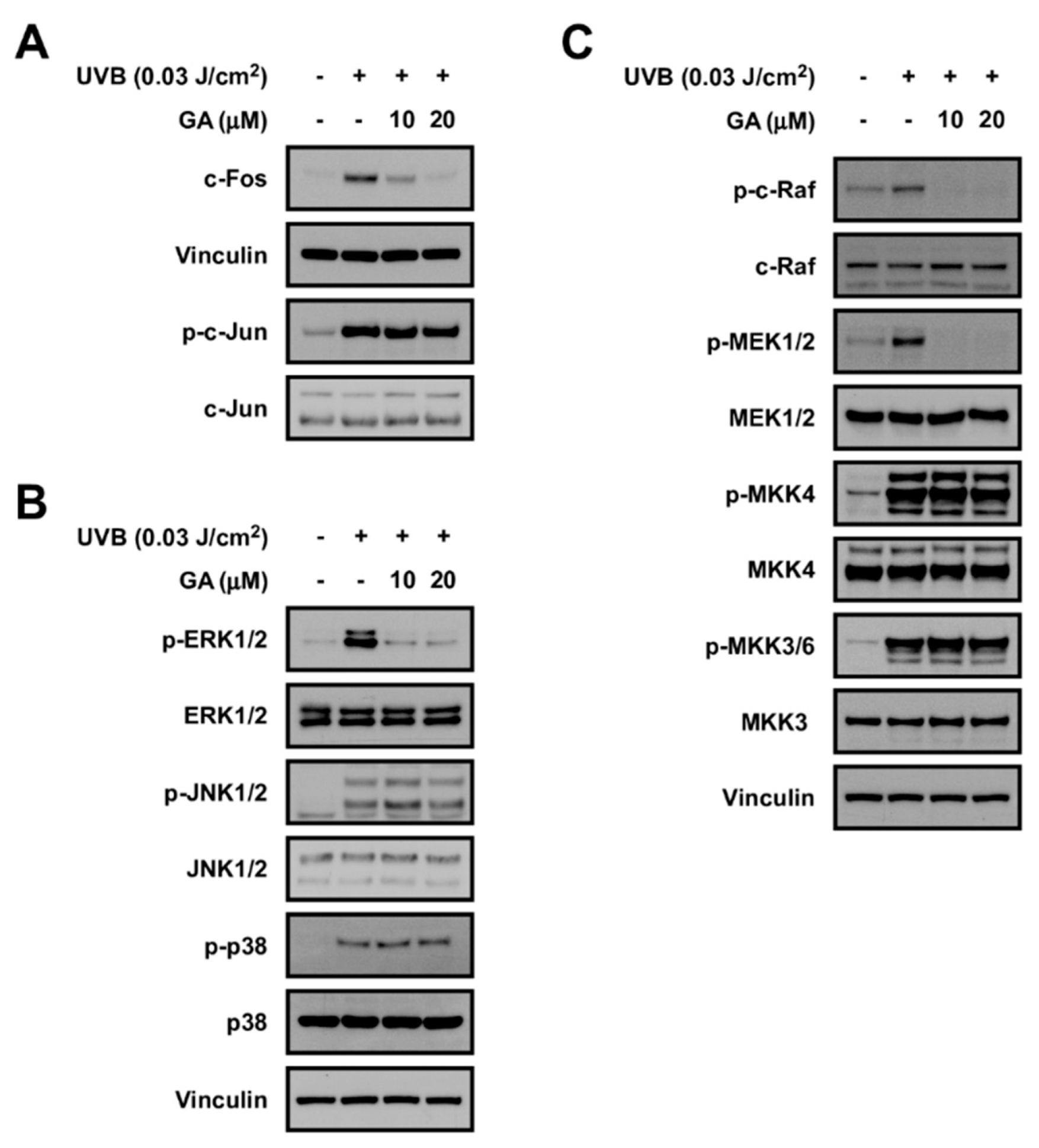
3.6. Gallic Acid Targets the c−Raf Signaling Pathway
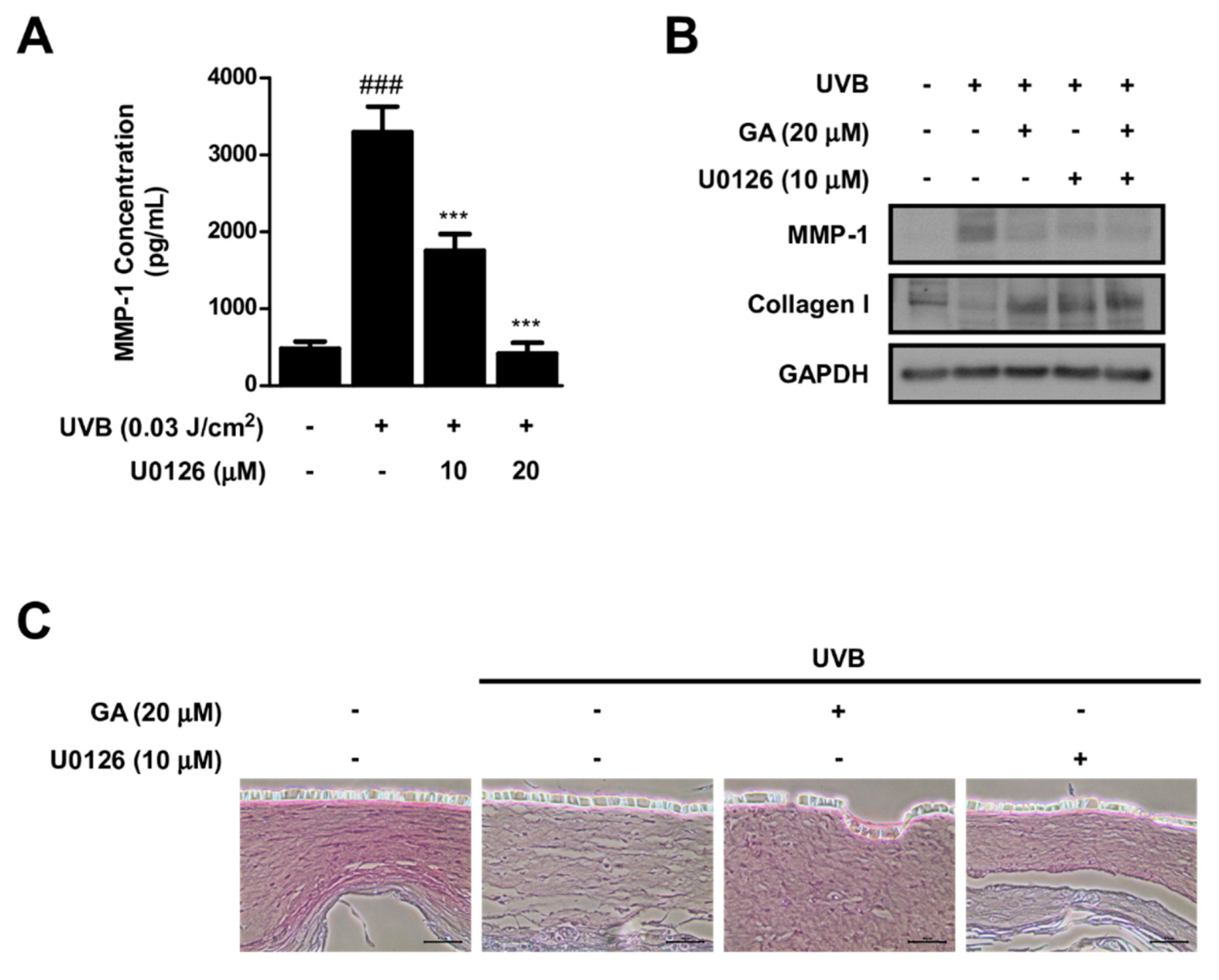
3.7. MEK Inhibition and Gallic Acid Blocks UVB−Induced MMP−1 Expression and Collagen Reduction in Reconstructed 3D Human Skin Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, prevention and therapy. Br. J. Derm. 2007, 157, 874–887. [Google Scholar] [CrossRef]
- Perez-Sanchez, A.; Barrajon-Catalan, E.; Herranz-Lopez, M.; Micol, V. Nutraceuticals for Skin Care: A Comprehensive Review of Human Clinical Studies. Nutrients 2018, 10, 403. [Google Scholar] [CrossRef] [Green Version]
- Rittie, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Derm. 2002, 138, 1462–1470. [Google Scholar] [CrossRef]
- Surowiak, P.; Gansukh, T.; Donizy, P.; Halon, A.; Rybak, Z. Increase in cyclooxygenase-2 (COX-2) expression in keratinocytes and dermal fibroblasts in photoaged skin. J. Cosmet Derm. 2014, 13, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.; Auler, S.; Hugo, R.; Gonzalez, S. Beneficial regulation of matrix metalloproteinases for skin health. Enzym. Res. 2011, 2011, 427285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, M.A.; Salem, S.A.; Hakim, S.A.; Shalan, Y.A. Comparative immunohistochemical assessment of cutaneous cyclooxygenase-2 enzyme expression in chronological aging and photoaging. Photodermatol. Photoimmunol. Photomed. 2014, 30, 43–51. [Google Scholar] [CrossRef]
- Rittie, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Angel, P.; Szabowski, A.; Schorpp-Kistner, M. Function and regulation of AP-1 subunits in skin physiology and pathology. Oncogene 2001, 20, 2413–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Alhendi, A.M.N.; Yeh, M.C.; Elahy, M.; Santiago, F.S.; Deshpande, N.P.; Wu, B.; Chan, E.; Inam, S.; Prado-Lourenco, L.; et al. Thermostable small-molecule inhibitor of angiogenesis and vascular permeability that suppresses a pERK-FosB/DeltaFosB-VCAM-1 axis. Sci Adv. 2020, 6, eaaz7815. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.A.; Cruzalegui, F.H.; Treisman, R. The p38 and ERK MAP kinase pathways cooperate to activate Ternary Complex Factors and c-fos transcription in response to UV light. EMBO J. 1996, 15, 6552–6563. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, R.; Lobo, M.; Trindade, K.; Silva, D.F.; Pereira, N. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging. Ski. Pharm. Physiol 2019, 32, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Radler-Pohl, A.; Sachsenmaier, C.; Gebel, S.; Auer, H.P.; Bruder, J.T.; Rapp, U.; Angel, P.; Rahmsdorf, H.J.; Herrlich, P. UV-induced activation of AP-1 involves obligatory extranuclear steps including Raf-1 kinase. EMBO J. 1993, 12, 1005–1012. [Google Scholar] [CrossRef]
- Bode, A.M.; Dong, Z. Mitogen-activated protein kinase activation in UV-induced signal transduction. Sci. STKE 2003, 2003, RE2. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Camarillo, C.; Ocampo, E.A.; Casamichana, M.L.; Perez-Plasencia, C.; Alvarez-Sanchez, E.; Marchat, L.A. Protein kinases and transcription factors activation in response to UV-radiation of skin: Implications for carcinogenesis. Int. J. Mol. Sci. 2012, 13, 142–172. [Google Scholar] [CrossRef]
- Fisher, G.J.; Voorhees, J.J. Molecular Mechanisms of Photoaging and its Prevention by Retinoic Acid: Ultraviolet Irradiation Induces MAP Kinase Signal Transduction Cascades that Induce Ap-1-Regulated Matrix Metalloproteinases that Degrade Human Skin In Vivo. J. Investig. Dermatol. Symp. Proc. 1998, 3, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrubasik, C.; Roufogalis, B.D.; Muller-Ladner, U.; Chrubasik, S. A systematic review on the Rosa canina effect and efficacy profiles. Phytother. Res. 2008, 22, 725–733. [Google Scholar] [CrossRef]
- Boskabady, M.H.; Shafei, M.N.; Saberi, Z.; Amini, S. Pharmacological effects of Rosa Damascena. Iran. J. Basic Med. Sci. 2011, 14, 295–307. [Google Scholar] [CrossRef]
- Fujii, T.; Ikeda, K.; Saito, M. Inhibitory effect of rose hip (Rosa canina L.) on melanogenesis in mouse melanoma cells and on pigmentation in brown guinea pigs. Biosci. Biotechnol. Biochem. 2011, 75, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.; Kim, D.H.; Nho, Y.H.; Park, J.E.; Kim, S.N.; Choi, E.H. A Mixture of Extracts of Kochia scoparia and Rosa multiflora with PPAR α/γ Dual Agonistic Effects Prevents Photoaging in Hairless Mice. Int. J. Mol. Sci. 2016, 17, 1919. [Google Scholar] [CrossRef] [Green Version]
- Pires, T.C.S.P.; Dias, M.I.; Barros, L.; Calhelha, R.C.; Alves, M.J.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Edible flowers as sources of phenolic compounds with bioactive potential. Food Res. Int. 2018, 105, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Koczka, N.; Stefanovits-Banyai, E.; Ombodi, A. Total Polyphenol Content and Antioxidant Capacity of Rosehips of Some Rosa Species. Medicines 2018, 5, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.J.; Han, A.R.; Lee, M.H.; Song, Y.R.; Lee, K.M.; Nam, T.G.; Lee, P.; Lee, S.Y.; Lim, T.G. Extraction conditions for Rosa gallica petal extracts with anti-skin aging activities. Food Sci. Biotechnol. 2019, 28, 1439–1446. [Google Scholar] [CrossRef]
- Shin, S.H.; Lee, J.S.; Zhang, J.M.; Choi, S.; Boskovic, Z.V.; Zhao, R.; Song, M.; Wang, R.; Tian, J.; Lee, M.H.; et al. Synthetic lethality by targeting the RUVBL1/2-TTT complex in mTORC1-hyperactive cancer cells. Sci. Adv. 2020, 6, eaay9131. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Kume, T.; Fujimura, T.; Kawada, H.; Moriwaki, S.; Takema, Y. The formation of wrinkles caused by transition of keratin intermediate filaments after repetitive UVB exposure. Arch. Derm. Res. 2005, 296, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Yamaba, H.; Haba, M.; Kunita, M.; Sakaida, T.; Tanaka, H.; Yashiro, Y.; Nakata, S. Morphological change of skin fibroblasts induced by UV Irradiation is involved in photoaging. Exp. Derm. 2016, 25 (Suppl. S3), 45–51. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.Z.; Xing, J.; Sun, M.; Zhan, Z.Q.; Corke, H. Phenolic Antioxidants (Hydrolyzable Tannins, Flavonols, and Anthocyanins) Identified by LC-ESI-MS and MALDI-QIT-TOF MS from Rosa chinensis flowers. J. Agric. Food Chem. 2005, 53, 9940–9948. [Google Scholar] [CrossRef]
- Barros, L.; Alves, C.T.; Duenas, M.; Silva, S.; Oliveira, R.; Carvalho, A.M.; Henriques, M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Characterization of phenolic compounds in wild medicinal flowers from Portugal by HPLC-DAD-ESI/MS and evaluation of antifungal properties. Ind. Crop. Prod. 2013, 44, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Bhandari, P.; Singh, B.; Bari, S.S. Antioxidant activity and ultra-performance LC-electrospray ionization-quadrupole time-of-flight mass spectrometry for phenolics-based fingerprinting of Rose species: Rosa damascena, Rosa bourboniana and Rosa brunonii. Food Chem. Toxicol. 2009, 47, 361–367. [Google Scholar] [CrossRef]
- Ochir, S.; Nishizawa, M.; Park, B.J.; Ishii, K.; Kanazawa, T.; Funaki, M.; Yamagishi, T. Inhibitory effects of Rosa gallica on the digestive enzymes. J. Nat. Med. 2010, 64, 275–280. [Google Scholar] [CrossRef]
- Qing, L.S.; Xue, Y.; Zhang, J.G.; Zhang, Z.F.; Liang, J.; Jiang, Y.; Liu, Y.M.; Liao, X. Identification of flavonoid glycosides in Rosa chinensis flowers by liquid chromatography-tandem mass spectrometry in combination with 13C nuclear magnetic resonance. J. Chromatogr. A 2012, 1249, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozga, J.A.; Saeed, A.; Wismer, W.; Reinecke, D.M. Characterization of cyanidin- and quercetin-derived flavonoids and other phenolics in mature saskatoon fruits (Amelanchier alnifolia Nutt.). J. Agric. Food Chem. 2007, 55, 10414–10424. [Google Scholar] [CrossRef] [PubMed]
- Elejalde-Palmett, C.; de Bernonville, T.D.; Glevarec, G.; Pichon, O.; Papon, N.; Courdavault, V.; St-Pierre, B.; Giglioli-Guivarc’h, N.; Lanoue, A.; Besseau, S. Characterization of a spermidine hydroxycinnamoyltransferase in Malus domestica highlights the evolutionary conservation of trihydroxycinnamoyl spermidines in pollen coat of core Eudicotyledons. J. Exp. Bot. 2015, 66, 7271–7285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef]
- Hoyos, B.; Imam, A.; Korichneva, I.; Levi, E.; Chua, R.; Hammerling, U. Activation of c-Raf kinase by ultraviolet light. Regulation by retinoids. J. Biol. Chem. 2002, 277, 23949–23957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.H.; Nam, T.G.; Lee, I.; Shin, E.J.; Han, A.R.; Lee, P.; Lee, S.Y.; Lim, T.G. Skin anti-inflammatory activity of rose petal extract (Rosa gallica) through reduction of MAPK signaling pathway. Food Sci. Nutr. 2018, 6, 2560–2567. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.-J.; Kim, S.-N.; Kim, J.-K.; Lee, B.-G.; Chang, I.-S. Effect of Green Tea Catechins on the Expression and Activity of MMPs and Type I Procollagen Synthesis in Human Dermal Fibroblasts. J. Soc. Cosmet. Sci. Korea 2006, 32, 117–121. [Google Scholar]
- Shin, E.J.; Lee, J.S.; Hong, S.; Lim, T.G.; Byun, S. Quercetin Directly Targets JAK2 and PKCδ and Prevents UV-Induced Photoaging in Human Skin. Int. J. Mol. Sci. 2019, 20, 5262. [Google Scholar] [CrossRef] [Green Version]
- Sudhakaran, M.; Parra, M.R.; Stoub, H.; Gallo, K.A.; Doseff, A.I. Apigenin by targeting hnRNPA2 sensitizes triple-negative breast cancer spheroids to doxorubicin-induced apoptosis and regulates expression of ABCC4 and ABCG2 drug efflux transporters. Biochem. Pharm. 2020, 182, 114259. [Google Scholar] [CrossRef]
- Lin, C.F.; Leu, Y.L.; Al-Suwayeh, S.A.; Ku, M.C.; Hwang, T.L.; Fang, J.Y. Anti-inflammatory activity and percutaneous absorption of quercetin and its polymethoxylated compound and glycosides: The relationships to chemical structures. Eur. J. Pharm. Sci. 2012, 47, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraj, P.; Nakagawa, H.; Yamaki, K. Cyanidin and cyanidin-3-glucoside derived from Vigna unguiculata act as noncompetitive inhibitors of pancreatic lipase. J. Food Biochem. 2019, 43, e12774. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Zhou, B.; Yang, L.; Liu, Z.L. Inhibition of human low density lipoprotein oxidation by flavonols and their glycosides. Chem. Phys. Lipids 2004, 129, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Alam, M.B.; Lee, S.H. Protection of UVB-Induced Photoaging by Fuzhuan-Brick Tea Aqueous Extract via MAPKs/Nrf2-Mediated Down-Regulation of MMP-1. Nutrients 2018, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, E.; Park, S.Y.; Lee, H.J.; Lee, T.Y.; Sun, Z.W.; Yi, T.H. Gallic acid regulates skin photoaging in UVB-exposed fibroblast and hairless mice. Phytother. Res. 2014, 28, 1778–1788. [Google Scholar] [CrossRef]
- Green, C.L.; Khavari, P.A. Targets for molecular therapy of skin cancer. Semin. Cancer Biol. 2004, 14, 63–69. [Google Scholar] [CrossRef]
- Khazak, V.; Astsaturov, I.; Serebriiskii, I.G.; Golemis, E.A. Selective Raf inhibition in cancer therapy. Expert Opin. Ther. Targets 2007, 11, 1587–1609. [Google Scholar] [CrossRef]
- Assi, M.; Achouri, Y.; Loriot, A.; Dauguet, N.; Dahou, H.; Baldan, J.; Libert, M.; Fain, J.S.; Guerra, C.; Bouwens, L.; et al. Dynamic Regulation of Expression of KRAS and Its Effectors Determines the Ability to Initiate Tumorigenesis in Pancreatic Acinar Cells. Cancer Res. 2021, 81, 2679–2689. [Google Scholar] [CrossRef]
- Farrand, L.; Byun, S. Induction of Synthetic Lethality by Natural Compounds Targeting Cancer Signaling. Curr. Pharm. Des. 2017, 23, 4311–4320. [Google Scholar] [CrossRef]
- Brunetti, G.; Di Rosa, G.; Scuto, M.; Leri, M.; Stefani, M.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Maintenance and Prevention of Parkinson’s-like Phenotypes with Hydroxytyrosol and Oleuropein Aglycone in C. elegans. Int. J. Mol. Sci. 2020, 21, 2588. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, V.; Cornelius, C.; Dinkova-Kostova, A.T.; Calabrese, E.J.; Mattson, M.P. Cellular stress responses, the hormesis paradigm, and vitagenes: Novel targets for therapeutic intervention in neurodegenerative disorders. Antioxid. Redox Signal. 2010, 13, 1763–1811. [Google Scholar] [CrossRef]
- Miquel, S.; Champ, C.; Day, J.; Aarts, E.; Bahr, B.A.; Bakker, M.; Banati, D.; Calabrese, V.; Cederholm, T.; Cryan, J.; et al. Poor cognitive ageing: Vulnerabilities, mechanisms and the impact of nutritional interventions. Ageing Res. Rev. 2018, 42, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Du, J.; Hwang, E.; Yi, T.H. Paeonol extracted from Paeonia suffruticosa Andr. ameliorated UVB-induced skin photoaging via DLD/Nrf2/ARE and MAPK/AP-1 pathway. Phytother. Res. 2018, 32, 1741–1749. [Google Scholar] [CrossRef]
- Feng, R.B.; Wang, Y.; He, C.; Yang, Y.; Wan, J.B. Gallic acid, a natural polyphenol, protects against tert-butyl hydroperoxide- induced hepatotoxicity by activating ERK-Nrf2-Keap1-mediated antioxidative response. Food Chem. Toxicol. 2018, 119, 479–488. [Google Scholar] [CrossRef]
- Jimenez, S.; Gascon, S.; Luquin, A.; Laguna, M.; Ancin-Azpilicueta, C.; Rodriguez-Yoldi, M.J. Rosa canina Extracts Have Antiproliferative and Antioxidant Effects on Caco-2 Human Colon Cancer. PLoS ONE 2016, 11, e0159136. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | tR (min) | Molecular Weight | Ion Mode | Precursor Ion (m/z) | Fragment Ions (m/z) | Tentative Identification | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 2.16 | 170.1195 | [M − H]− | 169.0000 | N.D. | Gallic acid | [30] |
| 2 | 4.04 | 290.2681 | [M + H]+ | 291.0864 | N.D. | Catechin | [29] |
| 3 | 4.28 | 954.7038 | [M + H]+ | 955.1054 | N.D. | Rugosin B | [28] |
| 4 | 6.50 | 936.6454 | [M + H]+ | 937.0934 | N.D. | Casuarictin | [31] |
| 5 | 7.45 | 616.4806 | [M + H]+ | 617.1137 | 303.0499 | Quercetin-3-O-gallate-glucoside | [32] |
| 6 | 8.00 | 610.5175 | [M + H]+ | 611.1607 | 465.1029, 303.0500 | Rutin | [30] |
| 7 | 8.28 | 464.3763 | [M + H]+ | 465.1028 | 303.0499 | Quercetin-3-O-galactoside | [28,33] |
| 8 | 8.56 | 464.3763 | [M + H]+ | 465.1026 | 303.0498 | Quercetin-3-O-glucoside | [30,33] |
| 9 | 9.51 | 448.3769 | [M + H]+ | 449.1078 | 287.0549 | Kaempferol-3-O-galactoside | [28,33] |
| 10 | 10.00 | 448.3769 | [M + H]+ | 449.1080 | 287.0550 | Kaempferol-3-O-glucoside | [30,33] |
| 11 | 10.20 | 448.3769 | [M + H]+ | 449.1078 | 303.0498 | Quercitrin | [30] |
| 12 | 11.94 | 438.5128 | [M − H]− | 437.1445 | N.D. | unknown | |
| 13 | 12.44 | 432.3775 | [M + H]+ | 433.1130 | 287.0550 | Kaempferol-3-O-rhamnoside | [30] |
| 14 | 13.10 | 652.5542 | [M + H]+ | 653.1712 | 303.0500 | Quercetin diglycoside | [30] |
| 15 | 14.78 | 636.5548 | [M + H]+ | 637.1764 | 303.0500 | Quercetin diglycoside | [30] |
| 16 | 16.96 | 583.6741 | [M + H]+ | 584.2754 | 438.2387 | Tricoumaroyl spermidine | [34] |
| 17 | 19.12 | 288.2522 | [M + H]+ | 289.2373 | 271.2268 | Dihydrokaempferol | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, S.; Jung, Y.-S.; Cho, Y.-R.; Seo, J.-W.; Lim, W.-C.; Nam, T.-G.; Lim, T.-G.; Byun, S. Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis. Antioxidants 2021, 10, 1663. https://doi.org/10.3390/antiox10111663
Jo S, Jung Y-S, Cho Y-R, Seo J-W, Lim W-C, Nam T-G, Lim T-G, Byun S. Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis. Antioxidants. 2021; 10(11):1663. https://doi.org/10.3390/antiox10111663
Chicago/Turabian StyleJo, Seongin, Young-Sung Jung, Ye-Ryeong Cho, Ji-Won Seo, Won-Chul Lim, Tae-Gyu Nam, Tae-Gyu Lim, and Sanguine Byun. 2021. "Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis" Antioxidants 10, no. 11: 1663. https://doi.org/10.3390/antiox10111663
APA StyleJo, S., Jung, Y. -S., Cho, Y. -R., Seo, J. -W., Lim, W. -C., Nam, T. -G., Lim, T. -G., & Byun, S. (2021). Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis. Antioxidants, 10(11), 1663. https://doi.org/10.3390/antiox10111663