Embryo and Its Mitochondria

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
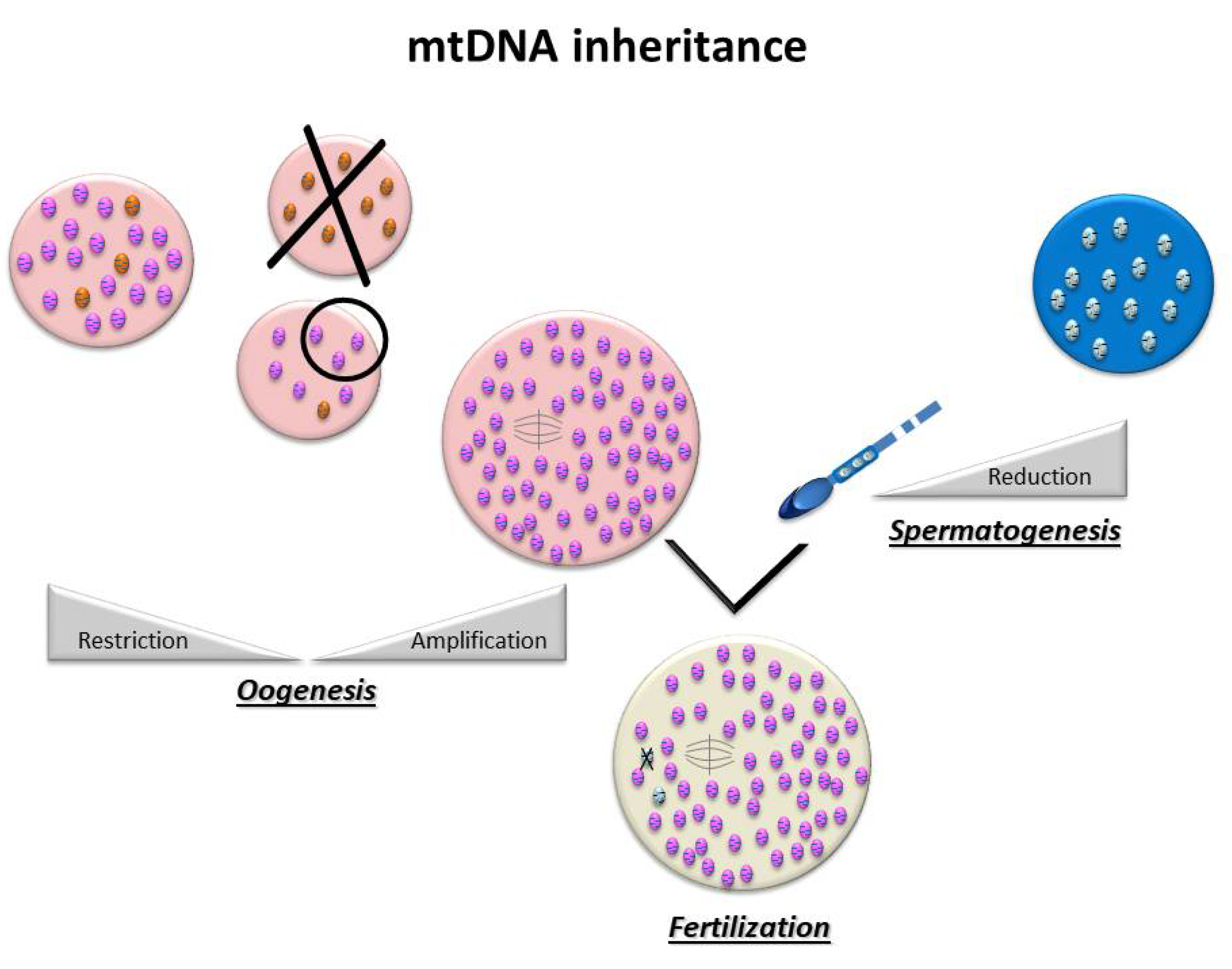
2. Mitochondria, Gametogenesis, and mtDNA Transmission
3. Mitochondria and the Embryo: Pathophysiological Aspects
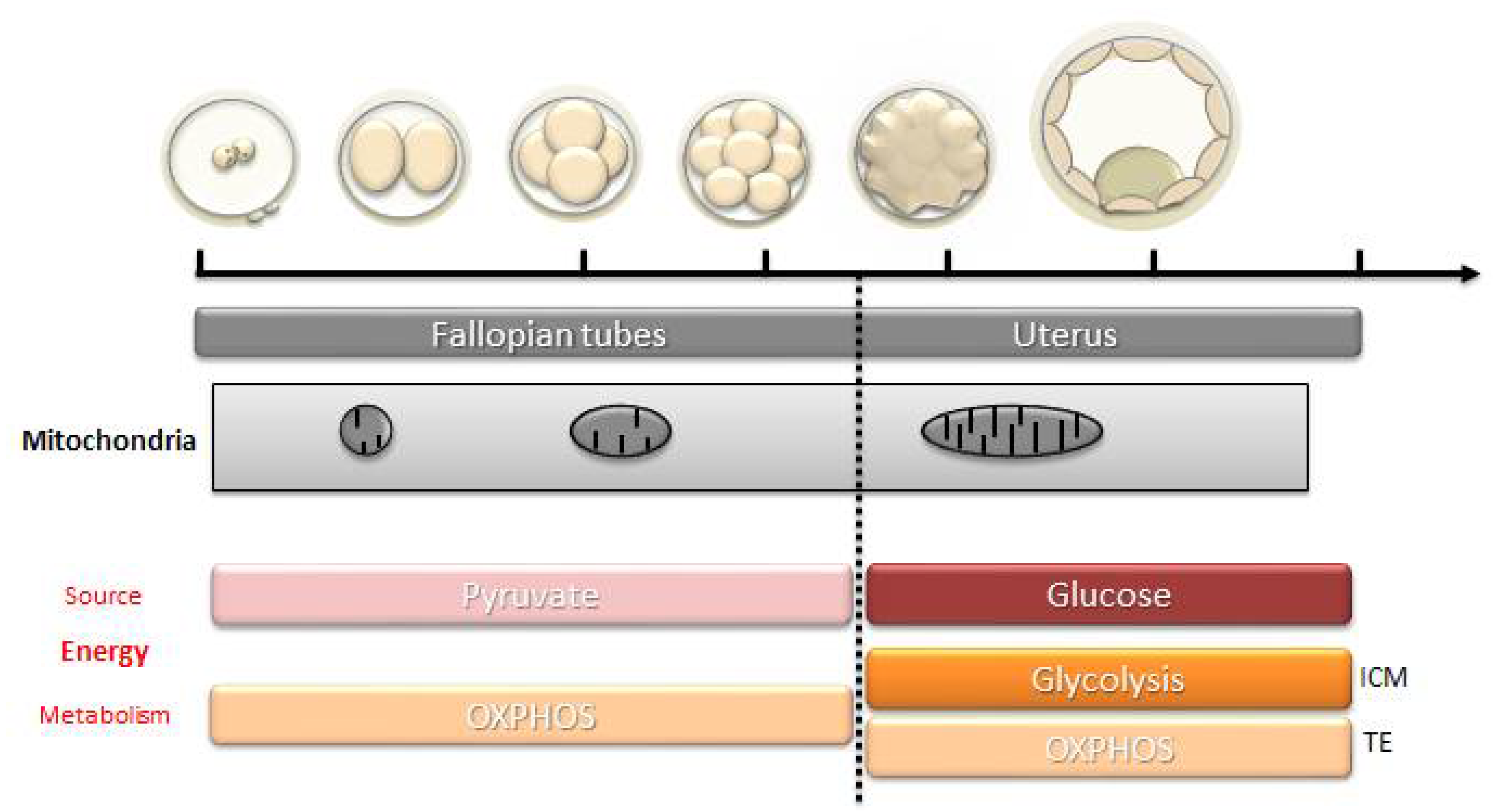
3.1. The Importance of the Mitochondrial Pool for the Initiation of Embryo Development
3.2. Ensure a Sufficient Energy Production
3.3. The Paradox of the Cleavage-Stage Embryo: Oxidative Metabolism and Low Mitochondrial Activity
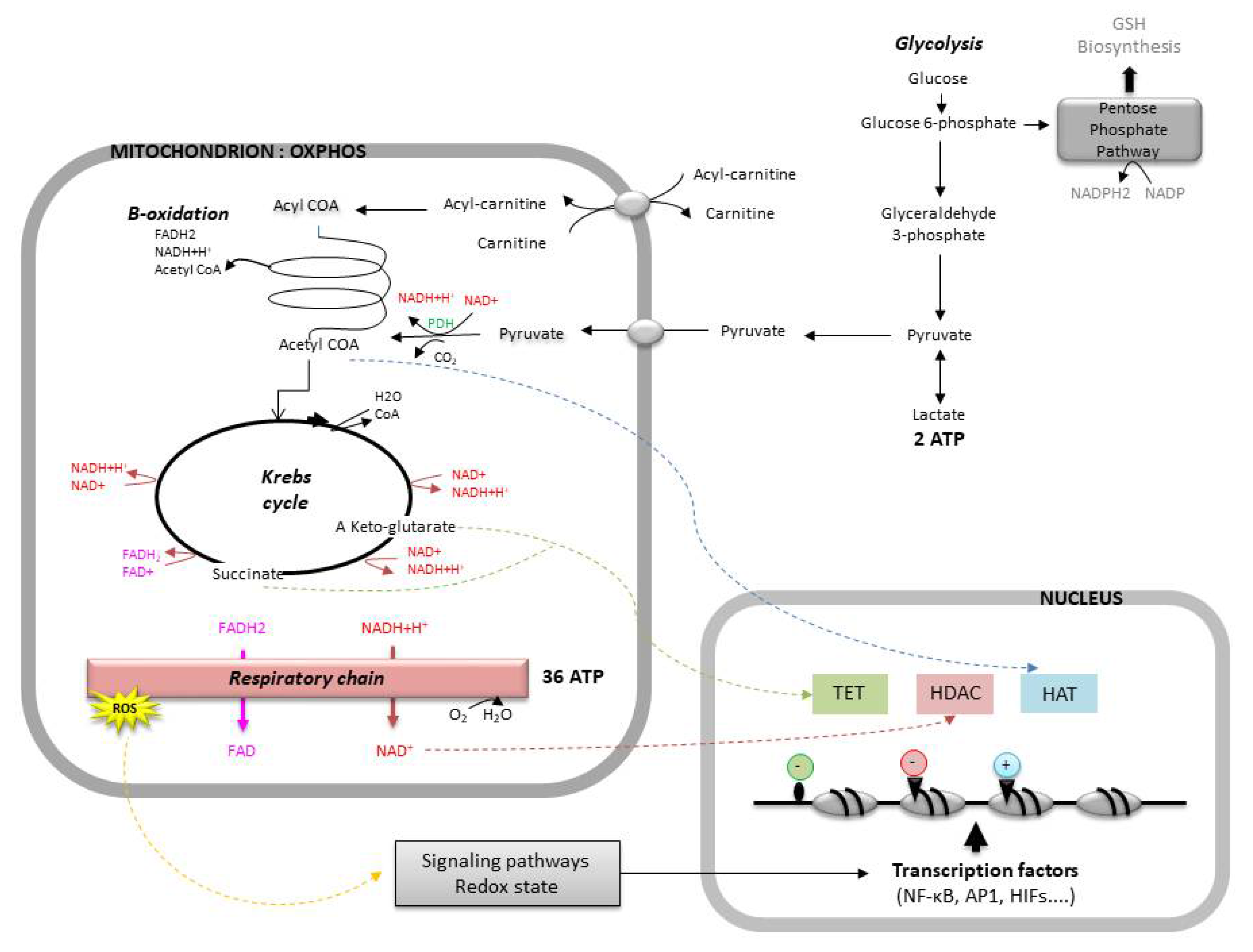
3.3.1. Limiting Oxidative Stress
3.3.2. Maintaining Redox Homeostasis
3.3.3. Transcriptional Regulation and Embryo Genome Activation
3.4. Resumption of Mitochondrial Biogenesis: From Morula to Blastocyst
3.4.1. The Set-Point Theory
3.4.2. Abnormal Resumption of Mitochondrial Biogenesis
4. Mitochondria and Biomarkers of Embryo Development
4.1. Mitochondria of the Cumulus Cells
4.2. Free mtDNA
4.3. MtDNA and Trophectoderm Biopsy
5. Mitochondria as Targets to Improve Fertility
5.1. Improving the Mitochondrial Function
5.2. Cytoplasmic and Mitochondrial Transfer
5.2.1. Mitochondrial Replacement Therapy
5.2.2. Cytoplasmic or Mitochondrial Transfer for the Treatment of Infertility
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gray, M.W. Mitochondrial Evolution. Science 1999, 283, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Gut, P.; Verdin, E. The Nexus of Chromatin Regulation and Intermediary Metabolism. Nature 2013, 502, 489–498. [Google Scholar] [CrossRef]
- Shadel, G.S.; Horvath, T.L. Mitochondrial ROS Signaling in Organismal Homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, N.S. Mitochondria as Signaling Organelles. BMC Biol. 2014, 12, 34. [Google Scholar] [CrossRef]
- Archer, S.L. Mitochondrial Dynamics—Mitochondrial Fission and Fusion in Human Diseases. N. Engl. J. Med. 2013, 369, 2236–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and Organization of the Human Mitochondrial Genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Popov, L. Mitochondrial Biogenesis: An Update. J. Cell. Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, C.M.; Falkenberg, M.; Larsson, N.-G. Maintenance and Expression of Mammalian Mitochondrial DNA. Annu. Rev. Biochem. 2016, 85, 133–160. [Google Scholar] [CrossRef]
- Giles, R.E.; Blanc, H.; Cann, H.M.; Wallace, D.C. Maternal Inheritance of Human Mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 6715–6719. [Google Scholar] [CrossRef] [Green Version]
- Carelli, V. Keeping in Shape the Dogma of Mitochondrial DNA Maternal Inheritance. PLoS Genet. 2015, 11, e1005179. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Sato, K. Maternal Inheritance of Mitochondrial DNA by Diverse Mechanisms to Eliminate Paternal Mitochondrial DNA. Biochim. Biophys. Acta BBA Mol. Cell Res. 2013, 1833, 1979–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otten, A.B.C.; Smeets, H.J.M. Evolutionary Defined Role of the Mitochondrial DNA in Fertility, Disease and Ageing. Hum. Reprod. Update 2015, 21, 671–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May-Panloup, P.; Chrétien, M.F.; Jacques, C.; Vasseur, C.; Malthièry, Y.; Reynier, P. Low Oocyte Mitochondrial DNA Content in Ovarian Insufficiency. Hum. Reprod. 2005, 20, 593–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauswirth, W.W.; Laipis, P.J. Mitochondrial DNA Polymorphism in a Maternal Lineage of Holstein Cows. Proc. Natl. Acad. Sci. USA 1982, 79, 4686–4690. [Google Scholar] [CrossRef] [Green Version]
- Olivo, P.D.; Van de Walle, M.J.; Laipis, P.J.; Hauswirth, W.W. Nucleotide Sequence Evidence for Rapid Genotypic Shifts in the Bovine Mitochondrial DNA D-Loop. Nature 1983, 306, 400–402. [Google Scholar] [CrossRef]
- Marchington, D.R.; Hartshorne, G.M.; Barlow, D.; Poulton, J. Homopolymeric Tract Heteroplasmy in MtDNA from Tissues and Single Oocytes: Support for a Genetic Bottleneck. Am. J. Hum. Genet. 1997, 60, 408–416. [Google Scholar]
- Poulton, J.; Macaulay, V.; Marchington, D.R. Is the Bottleneck Cracked? Am. J. Hum. Genet. 1998, 62, 752–757. [Google Scholar] [CrossRef] [Green Version]
- Howell, N.; Halvorson, S.; Kubacka, I.; McCullough, D.A.; Bindoff, L.A.; Turnbull, D.M. Mitochondrial Gene Segregation in Mammals: Is the Bottleneck Always Narrow? Hum. Genet. 1992, 90. [Google Scholar] [CrossRef]
- Hauswirth, W.; Laipis, P.J. Transmission genetics of mammalian mitochondria: A molecular model and experimental evidence. In Achievement and Perspectives of Mitochondrial Research; Elsevier: Amsterdam, The Netherlands, 1985; Volume 2, pp. 49–59. [Google Scholar]
- Cree, L.M.; Samuels, D.C.; de Sousa Lopes, S.C.; Rajasimha, H.K.; Wonnapinij, P.; Mann, J.R.; Dahl, H.-H.M.; Chinnery, P.F. A Reduction of Mitochondrial DNA Molecules during Embryogenesis Explains the Rapid Segregation of Genotypes. Nat. Genet. 2008, 40, 249–254. [Google Scholar] [CrossRef]
- Wai, T.; Teoli, D.; Shoubridge, E.A. The Mitochondrial DNA Genetic Bottleneck Results from Replication of a Subpopulation of Genomes. Nat. Genet. 2008, 40, 1484–1488. [Google Scholar] [CrossRef]
- Cree, L.M.; Samuels, D.C.; Chinnery, P.F. The Inheritance of Pathogenic Mitochondrial DNA Mutations. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2009, 1792, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.G.; Burgstaller, J.P.; Havlicek, V.; Kolbe, T.; Rülicke, T.; Brem, G.; Poulton, J.; Jones, N.S. Stochastic Modelling, Bayesian Inference, and New in vivo Measurements Elucidate the Debated MtDNA Bottleneck Mechanism. eLife 2015, 4, e07464. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Shitara, H.; Horii, T.; Nagao, Y.; Imai, H.; Abe, K.; Hara, T.; Hayashi, J.-I.; Yonekawa, H. The Mitochondrial Bottleneck Occurs without Reduction of MtDNA Content in Female Mouse Germ Cells. Nat. Genet. 2007, 39, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Khrapko, K.; Coller, H.A.; Andre, P.C.; Li, X.-C.; Hanekamp, J.S.; Thilly, W.G. Mitochondrial Mutational Spectra in Human Cells and Tissues. Proc. Natl. Acad. Sci. USA 1997, 94, 13798–13803. [Google Scholar] [CrossRef] [Green Version]
- Bratic, A.; Larsson, N.-G. The Role of Mitochondria in Aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Lin, C.S.; Potluri, P.; Procaccio, V.; Wallace, D.C. MtDNA Lineage Analysis of Mouse L-Cell Lines Reveals the Accumulation of Multiple MtDNA Mutants and Intermolecular Recombination. Genes Dev. 2012, 26, 384–394. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.C.; Chalkia, D. Mitochondrial DNA Genetics and the Heteroplasmy Conundrum in Evolution and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a021220. [Google Scholar] [CrossRef]
- Stewart, J.B.; Freyer, C.; Elson, J.L.; Wredenberg, A.; Cansu, Z.; Trifunovic, A.; Larsson, N.-G. Strong Purifying Selection in Transmission of Mammalian Mitochondrial DNA. PLoS Biol. 2008, 6, e10. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.B.; Chinnery, P.F. The Dynamics of Mitochondrial DNA Heteroplasmy: Implications for Human Health and Disease. Nat. Rev. Genet. 2015, 16, 530–542. [Google Scholar] [CrossRef]
- Wei, W.; Tuna, S.; Keogh, M.J.; Smith, K.R.; Aitman, T.J.; Beales, P.L.; Bennett, D.L.; Gale, D.P.; Bitner-Glindzicz, M.A.K.; Black, G.C.; et al. Germline Selection Shapes Human Mitochondrial DNA Diversity. Science 2019, 364, eaau6520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffann, J.; Monnot, S.; Bonnefont, J.-P. MtDNA Mutations Variously Impact MtDNA Maintenance throughout the Human Embryofetal Development. Clin. Genet. 2015, 88, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.J.; Carling, P.J.; Alston, C.L.; Floros, V.I.; Pyle, A.; Hudson, G.; Sallevelt, S.C.E.H.; Lamperti, C.; Carelli, V.; Bindoff, L.A.; et al. Mitochondrial DNA Sequence Characteristics Modulate the Size of the Genetic Bottleneck. Hum. Mol. Genet. 2016, 25, 1031–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumollard, R.; Carroll, J.; Duchen, M.R.; Campbell, K.; Swann, K. Mitochondrial Function and Redox State in Mammalian Embryos. Semin. Cell Dev. Biol. 2009, 20, 346–353. [Google Scholar] [CrossRef]
- Reynier, P.; May-Panloup, P.; Chretien, M.-F.; Morgan, C.J.; Jean, M.; Savagner, F.; Barriere, P.; Malthiery, Y. Mitochondrial DNA Content Affects the Fertilizability of Human Oocytes. Mol. Hum. Reprod. 2001, 7, 425–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, T.A.; El Shourbagy, S.; St. John, J.C. Mitochondrial Content Reflects Oocyte Variability and Fertilization Outcome. Fertil. Steril. 2006, 85, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakoshi, Y.; Sueoka, K.; Takahashi, K.; Sato, S.; Sakurai, T.; Tajima, H.; Yoshimura, Y. Embryo Developmental Capability and Pregnancy Outcome Are Related to the Mitochondrial DNA Copy Number and Ooplasmic Volume. J. Assist. Reprod. Genet. 2013, 30, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Tollner, T.L.; Hu, Z.; Dai, M.; Li, X.; Guan, H.; Shan, D.; Zhang, X.; Lv, J.; Huang, C.; et al. The Importance of Mitochondrial Metabolic Activity and Mitochondrial DNA Replication during Oocyte Maturation in vitro on Oocyte Quality and Subsequent Embryo Developmental Competence. Mol. Reprod. Dev. 2012, 79, 392–401. [Google Scholar] [CrossRef] [PubMed]
- El Shourbagy, S.H.; Spikings, E.C.; Freitas, M.; St John, J.C. Mitochondria Directly Influence Fertilisation Outcome in the Pig. Reproduction 2006, 131, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Pikó, L.; Taylor, K.D. Amounts of Mitochondrial DNA and Abundance of Some Mitochondrial Gene Transcripts in Early Mouse Embryos. Dev. Biol. 1987, 123, 364–374. [Google Scholar] [CrossRef]
- May-Panloup, P.; Vignon, X.; Chrétien, M.-F.; Heyman, Y.; Tamassia, M.; Malthièry, Y.; Reynier, P. Increase of Mitochondrial DNA Content and Transcripts in Early Bovine Embryogenesis Associated with Upregulation of MtTFA and NRF1 Transcription Factors. Reprod. Biol. Endocrinol. 2005, 3, 65. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, Y.; Filion, F.; Yoo, J.G.; Smith, L.C. Characterization of Mitochondrial Replication and Transcription Control during Rat Early Development in vivo and in vitro. Reproduction 2007, 133, 423–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spikings, E.C.; Alderson, J.; St. John, J.C. Regulated Mitochondrial DNA Replication During Oocyte Maturation Is Essential for Successful Porcine Embryonic Development. Biol. Reprod. 2007, 76, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.; Morimoto, N.; Yamanaka, M.; Matsumoto, H.; Yamochi, T.; Goto, H.; Inoue, M.; Nakaoka, Y.; Shibahara, H.; Morimoto, Y. Quantitative and Qualitative Changes of Mitochondria in Human Preimplantation Embryos. J. Assist. Reprod. Genet. 2017, 34, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConnell, J.M.; Petrie, L. Mitochondrial DNA Turnover Occurs during Preimplantation Development and Can Be Modulated by Environmental Factors. Reprod. Biomed. Online 2004, 9, 418–424. [Google Scholar] [CrossRef]
- Cagnone, G.L.M.; Tsai, T.-S.; Makanji, Y.; Matthews, P.; Gould, J.; Bonkowski, M.S.; Elgass, K.D.; Wong, A.S.A.; Wu, L.E.; McKenzie, M.; et al. Restoration of Normal Embryogenesis by Mitochondrial Supplementation in Pig Oocytes Exhibiting Mitochondrial DNA Deficiency. Sci. Rep. 2016, 6, 23229. [Google Scholar] [CrossRef]
- Gardner, D.K.; Wale, P.L. Analysis of Metabolism to Select Viable Human Embryos for Transfer. Fertil. Steril. 2013, 99, 1062–1072. [Google Scholar] [CrossRef]
- Collado-Fernandez, E.; Picton, H.M.; Dumollard, R. Metabolism throughout Follicle and Oocyte Development in Mammals. Int. J. Dev. Biol. 2012, 56, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.; Swann, K. Mitochondria and Lipid Metabolism in Mammalian Oocytes and Early Embryos. Int. J. Dev. Biol. 2019, 63, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Wilding, M.; Dale, B.; Marino, M.; di Matteo, L.; Alviggi, C.; Pisaturo, M.L.; Lombardi, L.; De Placido, G. Mitochondrial Aggregation Patterns and Activity in Human Oocytes and Preimplantation Embryos. Hum. Reprod. 2001, 16, 909–917. [Google Scholar] [CrossRef]
- Au, H.-K.; Yeh, T.-S.; Kao, S.-H.; Tzeng, C.-R.; Hsieh, R.-H. Abnormal Mitochondrial Structure in Human Unfertilized Oocytes and Arrested Embryos. Ann. N. Y. Acad. Sci. 2005, 1042, 177–185. [Google Scholar] [CrossRef]
- Van Blerkom, J.; Davis, P.W.; Lee, J. Fertilization and Early Embryolgoy: ATP Content of Human Oocytes and Developmental Potential and Outcome after in-Vitro Fertilization and Embryo Transfer. Hum. Reprod. 1995, 10, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J.; Davis, P.; Alexander, S. Differential Mitochondrial Distribution in Human Pronuclear Embryos Leads to Disproportionate Inheritance between Blastomeres: Relationship to Microtubular Organization, ATP Content and Competence. Hum. Reprod. 2000, 15, 2621–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motta, P.M.; Nottola, S.A.; Makabe, S.; Heyn, R. Mitochondrial Morphology in Human Fetal and Adult Female Germ Cells. Hum. Reprod. 2000, 15, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J. Mitochondria in Human Oogenesis and Preimplantation Embryogenesis: Engines of Metabolism, Ionic Regulation and Developmental Competence. Reproduction 2004, 128, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho-Santos, J.; Varum, S.; Amaral, S.; Mota, P.C.; Sousa, A.P.; Amaral, A. Mitochondrial Functionality in Reproduction: From Gonads and Gametes to Embryos and Embryonic Stem Cells. Hum. Reprod. Update 2009, 15, 553–572. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J. Mitochondrial Function in the Human Oocyte and Embryo and Their Role in Developmental Competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondria in Early Mammalian Development. Semin. Cell Dev. Biol. 2009, 20, 354–364. [Google Scholar] [CrossRef]
- Dumollard, R.; Ward, Z.; Carroll, J.; Duchen, M.R. Regulation of Redox Metabolism in the Mouse Oocyte and Embryo. Development 2006, 134, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Patergnani, S.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. Calcium Signaling around Mitochondria Associated Membranes (MAMs). Cell Commun. Signal. 2011, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Dumollard, R.; Duchen, M.; Sardet, C. Calcium Signals and Mitochondria at Fertilisation. Semin. Cell Dev. Biol. 2006, 17, 314–323. [Google Scholar] [CrossRef]
- Scantland, S.; Tessaro, I.; Macabelli, C.H.; Macaulay, A.D.; Cagnone, G.; Fournier, É.; Luciano, A.M.; Robert, C. The Adenosine Salvage Pathway as an Alternative to Mitochondrial Production of ATP in Maturing Mammalian Oocytes1. Biol. Reprod. 2014, 91. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.J. Mitochondria in Early Development: Linking the Microenvironment, Metabolism and the Epigenome. Reproduction 2019, 157, R159–R179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lees, J.G.; Gardner, D.K.; Harvey, A.J. Pluripotent Stem Cell Metabolism and Mitochondria: Beyond ATP. Stem Cells Int. 2017, 2017, 2874283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P. How Mitochondria Produce Reactive Oxygen Species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, C.L.; Perevoschikova, I.V.; Goncalves, R.L.S.; Hey-Mogensen, M.; Brand, M.D. The Determination and Analysis of Site-Specific Rates of Mitochondrial Reactive Oxygen Species Production. Methods Enzymol. 2013, 526, 189–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, F.L.; Liu, Y.; Van Remmen, H. Complex III Releases Superoxide to Both Sides of the Inner Mitochondrial Membrane. J. Biol. Chem. 2004, 279, 49064–49073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridovich, I. Superoxide Anion Radical (O2-), Superoxide Dismutases, and Related Matters. J. Biol. Chem. 1997, 272, 18515–18517. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Canali, R.; Rettori, D.; Kaplowitz, N. Effect of Glutathione Depletion on Sites and Topology of Superoxide and Hydrogen Peroxide Production in Mitochondria. Mol. Pharmacol. 2003, 64, 1136–1144. [Google Scholar] [CrossRef]
- Cross, C.E.; Halliwell, B.; Borish, E.; Pryor, W.; Ames, B.; Saul, R.; McCord, J.; Harman, D. Oxygen Radicals and Human Disease. Ann. Intern. Med. 1987, 107, 526. [Google Scholar] [CrossRef]
- Guerin, P. Oxidative Stress and Protection against Reactive Oxygen Species in the Pre-Implantation Embryo and Its Surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Grindler, N.M.; Moley, K.H. Maternal Obesity, Infertility and Mitochondrial Dysfunction: Potential Mechanisms Emerging from Mouse Model Systems. Mol. Hum. Reprod. 2013, 19, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.; Li, Z.; Zhang, Y.; Huang, Y.; Wu, Q.; Ru, G.; Chen, M.; Wang, B. Response of Mouse Zygotes Treated with Mild Hydrogen Peroxide as a Model to Reveal Novel Mechanisms of Oxidative Stress-Induced Injury in Early Embryos. Oxidative Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez-Pardo, C.; Morales, A.T.; Rio, M.J.D.; Olivera-Angel, M. Endogenously Generated Hydrogen Peroxide Induces Apoptosis via Mitochondrial Damage Independent of NF-ΚB and P53 Activation in Bovine Embryos. Theriogenology 2007, 67, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Bain, N.T.; Madan, P.; Betts, D.H. The Early Embryo Response to Intracellular Reactive Oxygen Species Is Developmentally Regulated. Reprod. Fertil. Dev. 2011, 23, 561. [Google Scholar] [CrossRef]
- Yang, H.W.; Hwang, K.J.; Kwon, H.C.; Kim, H.S.; Choi, K.W.; Oh, K.S. Detection of Reactive Oxygen Species (ROS) and Apoptosis in Human Fragmented Embryos. Hum. Reprod. 1998, 13, 998–1002. [Google Scholar] [CrossRef]
- Noda, Y.; Goto, Y.; Umaoka, Y.; Shiotani, M.; Nakayama, T.; Mori, T. Culture of Human Embryos in Alpha Modification of Eagle’s Medium under Low Oxygen Tension and Low Illumination. Fertil. Steril. 1994, 62, 1022–1027. [Google Scholar] [CrossRef]
- Ciray, H.N.; Aksoy, T.; Yaramanci, K.; Karayaka, I.; Bahceci, M. In vitro Culture under Physiologic Oxygen Concentration Improves Blastocyst Yield and Quality: A Prospective Randomized Survey on Sibling Oocytes. Fertil. Steril. 2009, 91, 1459–1461. [Google Scholar] [CrossRef]
- Dumoulin, J.C.; Meijers, C.J.; Bras, M.; Coonen, E.; Geraedts, J.P.; Evers, J.L. Effect of Oxygen Concentration on Human In-Vitro Fertilization and Embryo Culture. Hum. Reprod. 1999, 14, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Kovačič, B.; Vlaisavljević, V. Influence of Atmospheric versus Reduced Oxygen Concentration on Development of Human Blastocysts in vitro: A Prospective Study on Sibling Oocytes. Reprod. Biomed. Online 2008, 17, 229–236. [Google Scholar] [CrossRef]
- Belli, M.; Zhang, L.; Liu, X.; Donjacour, A.; Ruggeri, E.; Palmerini, M.G.; Nottola, S.A.; Macchiarelli, G.; Rinaudo, P. Oxygen Concentration Alters Mitochondrial Structure and Function in in vitro Fertilized Preimplantation Mouse Embryos. Hum. Reprod. 2019, 34, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M. Oxidative Stress and Redox Regulation on in vitro Development of Mammalian Embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Vidal, E.; Lomelí, H.; Castro-Obregón, S.; Cuervo, R.; Escalante-Alcalde, D.; Covarrubias, L. Reactive Oxygen Species Participate in the Control of Mouse Embryonic Cell Death. Exp. Cell Res. 1998, 238, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Ufer, C.; Wang, C.C.; Borchert, A.; Heydeck, D.; Kuhn, H. Redox Control in Mammalian Embryo Development. Antioxid. Redox Signal. 2010, 13, 833–875. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.S.; Lane, M.; Thompson, J.G. Oxygen Consumption and ROS Production Are Increased at the Time of Fertilization and Cell Cleavage in Bovine Zygotes. Hum. Reprod. 2010, 25, 2762–2773. [Google Scholar] [CrossRef] [Green Version]
- Morales, H.; Tilquin, P.; Rees, J.F.; Massip, A.; Dessy, F.; Van Langendonckt, A. Pyruvate Prevents Peroxide-Induced Injury of in vitro Preimplantation Bovine Embryos. Mol. Reprod. Dev. 1999, 52, 149–157. [Google Scholar] [CrossRef]
- Rhee, S.G. Cell Signaling. H2O2, a Necessary Evil for Cell Signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Orr, A.L.; Vargas, L.; Turk, C.N.; Baaten, J.E.; Matzen, J.T.; Dardov, V.J.; Attle, S.J.; Li, J.; Quackenbush, D.C.; Goncalves, R.L.S.; et al. Suppressors of Superoxide Production from Mitochondrial Complex III. Nat. Chem. Biol. 2015, 11, 834–836. [Google Scholar] [CrossRef] [Green Version]
- D’Autréaux, B.; Toledano, M.B. ROS as Signalling Molecules: Mechanisms That Generate Specificity in ROS Homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Janssen-Heininger, Y.M.W.; Mossman, B.T.; Heintz, N.H.; Forman, H.J.; Kalyanaraman, B.; Finkel, T.; Stamler, J.S.; Rhee, S.G.; van der Vliet, A. Redox-Based Regulation of Signal Transduction: Principles, Pitfalls, and Promises. Free Radic. Biol. Med. 2008, 45, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tonks, N.K. Redox Redux: Revisiting PTPs and the Control of Cell Signaling. Cell 2005, 121, 667–670. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-R.; Yang, K.-S.; Kwon, J.; Lee, C.; Jeong, W.; Rhee, S.G. Reversible Inactivation of the Tumor Suppressor PTEN by H2O2. J. Biol. Chem. 2002, 277, 20336–20342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, K.M.; Subbaram, S.; Regan, K.J.; Nelson, K.K.; Mazurkiewicz, J.E.; Bartholomew, P.J.; Aplin, A.E.; Tai, Y.-T.; Aguirre-Ghiso, J.; Flores, S.C.; et al. Mitochondrial H2O2 Regulates the Angiogenic Phenotype via PTEN Oxidation. J. Biol. Chem. 2005, 280, 16916–16924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.M.; Lee, K.S.; Woo, H.A.; Kang, D.; Rhee, S.G. Control of the Pericentrosomal H2O2 Level by Peroxiredoxin I Is Critical for Mitotic Progression. J. Cell Biol. 2015, 210, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarugi, P.; Pani, G.; Giannoni, E.; Taddei, L.; Colavitti, R.; Raugei, G.; Symons, M.; Borrello, S.; Galeotti, T.; Ramponi, G. Reactive Oxygen Species as Essential Mediators of Cell Adhesion: The Oxidative Inhibition of a FAK Tyrosine Phosphatase Is Required for Cell Adhesion. J. Cell Biol. 2003, 161, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Patterson, H.C.; Gerbeth, C.; Thiru, P.; Vögtle, N.F.; Knoll, M.; Shahsafaei, A.; Samocha, K.E.; Huang, C.X.; Harden, M.M.; Song, R.; et al. A Respiratory Chain Controlled Signal Transduction Cascade in the Mitochondrial Intermembrane Space Mediates Hydrogen Peroxide Signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E5679–E5688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xanthoudakis, S.; Curran, T. Identification and Characterization of Ref-1, a Nuclear Protein That Facilitates AP-1 DNA-Binding Activity. EMBO J. 1992, 11, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.R.; Wakasugi, N.; Virelizier, J.-L.; Yodoi, J.; Hay, R.T. Thiordoxin Regulates the DNA Binding Activity of NF-ΧB by Reduction of a Disulphid Bond Involving Cysteine 62. Nucl. Acids Res. 1992, 20, 3821–3830. [Google Scholar] [CrossRef] [PubMed]
- Mitomo, K.; Nakayama, K.; Fujimoto, K.; Sun, X.; Seki, S.; Yamamoto, K. Two Different Cellular Redox Systems Regulate the DNA-Binding Activity of the P50 Subunit of NF-ΚB in vitro. Gene 1994, 145, 197–203. [Google Scholar] [CrossRef]
- Shaulian, E.; Karin, M. AP-1 as a Regulator of Cell Life and Death. Nat. Cell Biol. 2002, 4, E131–E136. [Google Scholar] [CrossRef]
- Tsunoda, S.; Kibe, N.; Kurahashi, T.; Fujii, J. Differential Responses of SOD1-Deficient Mouse Embryonic Fibroblasts to Oxygen Concentrations. Arch. Biochem. Biophys. 2013, 537, 5–11. [Google Scholar] [CrossRef]
- Cagnone, G.; Sirard, M.-A. The Embryonic Stress Response to in vitro Culture: Insight from Genomic Analysis. Reproduction 2016, 152, R247–R261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luberda, Z. The Role of Glutathione in Mammalian Gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar] [PubMed]
- Abeydeera, L.R.; Wang, W.H.; Cantley, T.C.; Prather, R.S.; Day, B.N. Glutathione Content and Embryo Development after in vitro Fertilisation of Pig Oocytes Matured in the Presence of a Thiol Compound and Various Concentrations of Cysteine. Zygote 1999, 7, 203–210. [Google Scholar] [CrossRef] [PubMed]
- De Matos, D.G.; Furnus, C.C. The Importance of Having High Glutathione (GSH) Level after Bovine in vitro Maturation on Embryo Development: Effect of β-Mercaptoethanol, Cysteine and Cystine. Theriogenology 2000, 53, 761–771. [Google Scholar] [CrossRef]
- Fischer, B.; Bavister, B.D. Oxygen Tension in the Oviduct and Uterus of Rhesus Monkeys, Hamsters and Rabbits. Reproduction 1993, 99, 673–679. [Google Scholar] [CrossRef]
- Waypa, G.B.; Schumacker, P.T. Hypoxia-Induced Changes in Pulmonary and Systemic Vascular Resistance: Where Is the O2 Sensor? Respir. Physiol. Neurobiol. 2010, 174, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Chandel, N.S.; McClintock, D.S.; Feliciano, C.E.; Wood, T.M.; Melendez, J.A.; Rodriguez, A.M.; Schumacker, P.T. Reactive Oxygen Species Generated at Mitochondrial Complex III Stabilize Hypoxia-Inducible Factor-1α during Hypoxia. J. Biol. Chem. 2000, 275, 25130–25138. [Google Scholar] [CrossRef] [Green Version]
- Brunelle, J.K.; Bell, E.L.; Quesada, N.M.; Vercauteren, K.; Tiranti, V.; Zeviani, M.; Scarpulla, R.C.; Chandel, N.S. Oxygen Sensing Requires Mitochondrial ROS but Not Oxidative Phosphorylation. Cell Metab. 2005, 1, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Guzy, R.D.; Hoyos, B.; Robin, E.; Chen, H.; Liu, L.; Mansfield, K.D.; Simon, M.C.; Hammerling, U.; Schumacker, P.T. Mitochondrial Complex III Is Required for Hypoxia-Induced ROS Production and Cellular Oxygen Sensing. Cell Metab. 2005, 1, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Bailey, P.; Nathan, J. Metabolic Regulation of Hypoxia-Inducible Transcription Factors: The Role of Small Molecule Metabolites and Iron. Biomedicines 2018, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, D.C.; Brüne, B. Mitochondrial Composition and Function under the Control of Hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef]
- Wheaton, W.W.; Chandel, N.S. Hypoxia. 2. Hypoxia Regulates Cellular Metabolism. Am. J. Physiol. Cell Physiol. 2011, 300, C385–C393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almansa-Ordonez, A.; Bellido, R.; Vassena, R.; Barragan, M.; Zambelli, F. Oxidative Stress in Reproduction: A Mitochondrial Perspective. Biology 2020, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Mantikou, E.; Jonker, M.J.; Wong, K.M.; van Montfoort, A.P.A.; de Jong, M.; Breit, T.M.; Repping, S.; Mastenbroek, S. Factors Affecting the Gene Expression of in vitro Cultured Human Preimplantation Embryos. Hum. Reprod. 2015, 298–311. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, R.; Sharpley, M.S.; Chi, F.; Braas, D.; Zhou, Y.; Kim, R.; Clark, A.T.; Banerjee, U. Nuclear Localization of Mitochondrial TCA Cycle Enzymes as a Critical Step in Mammalian Zygotic Genome Activation. Cell 2017, 168, 210–223.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guantes, R.; Díaz-Colunga, J.; Iborra, F.J. Mitochondria and the Non-Genetic Origins of Cell-to-Cell Variability: More Is Different. BioEssays 2016, 38, 64–76. [Google Scholar] [CrossRef]
- Leese, H.J.; Guerif, F.; Allgar, V.; Brison, D.R.; Lundin, K.; Sturmey, R.G. Biological Optimization, the Goldilocks Principle, and How Much Is Lagom in the Preimplantation Embryo. Mol. Reprod. Dev. 2016, 83, 748–754. [Google Scholar] [CrossRef]
- Houghton, F.D.; Leese, H.J. Metabolism and Developmental Competence of the Preimplantation Embryo. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115, S92–S96. [Google Scholar] [CrossRef]
- St. John, J. The Control of MtDNA Replication during Differentiation and Development. Biochim. Biophys. Acta BBA Gen. Subj. 2014, 1840, 1345–1354. [Google Scholar] [CrossRef]
- Houghton, F.D. Energy Metabolism of the Inner Cell Mass and Trophectoderm of the Mouse Blastocyst. Differentiation 2006, 74, 11–18. [Google Scholar] [CrossRef]
- Kumar, R.P.; Ray, S.; Home, P.; Saha, B.; Bhattacharya, B.; Wilkins, H.M.; Chavan, H.; Ganguly, A.; Milano-Foster, J.; Paul, A.; et al. Regulation of Energy Metabolism during Early Mammalian Development: TEAD4 Controls Mitochondrial Transcription. Development 2018, 145, dev162644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, K.J. Metabolism of Preimplantation Embryo Development. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 120, pp. 259–310. ISBN 9780128014288. [Google Scholar]
- Sun, X.; St. John, J.C. The Role of the MtDNA Set Point in Differentiation, Development and Tumorigenesis. Biochem. J. 2016, 473, 2955–2971. [Google Scholar] [CrossRef] [PubMed]
- Diez-Juan, A.; Rubio, C.; Marin, C.; Martinez, S.; Al-Asmar, N.; Riboldi, M.; Díaz-Gimeno, P.; Valbuena, D.; Simón, C. Mitochondrial DNA Content as a Viability Score in Human Euploid Embryos: Less Is Better. Fertil. Steril. 2015, 104, 534–541.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May-Panloup, P.; Brochard, V.; Hamel, J.F.; Desquiret-Dumas, V.; Chupin, S.; Reynier, P.; Duranthon, V. Maternal Ageing Impairs Mitochondrial DNA Kinetics during Early Embryogenesis in Mice. Hum. Reprod. 2019, 34, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-Secreted Factors: Regulators of Cumulus Cell Function and Oocyte Quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef] [Green Version]
- Richani, D.; Dunning, K.R.; Thompson, J.G.; Gilchrist, R.B. Metabolic Co-Dependence of the Oocyte and Cumulus Cells: Essential Role in Determining Oocyte Developmental Competence. Hum. Reprod. Update 2021, 27, 27–47. [Google Scholar] [CrossRef]
- Seli, E.; Babayev, E.; Collins, S.C.; Nemeth, G.; Horvath, T.L. Minireview: Metabolism of Female Reproduction: Regulatory Mechanisms and Clinical Implications. Mol. Endocrinol. 2014, 28, 790–804. [Google Scholar] [CrossRef] [Green Version]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte Environment: Follicular Fluid and Cumulus Cells Are Critical for Oocyte Health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Guedikian, A.A.; Madrigal, V.K.; Phan, J.D.; Hill, D.L.; Alvarez, J.P.; Chazenbalk, G.D. Cumulus Cell Mitochondrial Resistance to Stress in vitro Predicts Oocyte Development During Assisted Reproduction. J. Clin. Endocrinol. Metab. 2016, 101, 2235–2245. [Google Scholar] [CrossRef]
- Sutton-McDowall, M.L.; Gilchrist, R.B.; Thompson, J.G. The Pivotal Role of Glucose Metabolism in Determining Oocyte Developmental Competence. Reproduction 2010, 139, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, K.; Pendola, F.L.; Eppig, J.J. Oocyte Control of Metabolic Cooperativity between Oocytes and Companion Granulosa Cells: Energy Metabolism. Dev. Biol. 2005, 279, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, K.; Su, Y.-Q.; Diaz, F.J.; Pangas, S.A.; Sharma, S.; Wigglesworth, K.; O’Brien, M.J.; Matzuk, M.M.; Shimasaki, S.; Eppig, J.J. Oocyte-Derived BMP15 and FGFs Cooperate to Promote Glycolysis in Cumulus Cells. Development 2007, 134, 2593–2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.-Q.; Sugiura, K.; Eppig, J.J. Mouse Oocyte Control of Granulosa Cell Development and Function: Paracrine Regulation of Cumulus Cell Metabolism. Semin. Reprod. Med. 2009, 27, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downs, S.M.; Mosey, J.L.; Klinger, J. Fatty Acid Oxidation and Meiotic Resumption in Mouse Oocytes. Mol. Reprod. Dev. 2009, 76, 844–853. [Google Scholar] [CrossRef] [Green Version]
- Dunning, K.R.; Cashman, K.; Russell, D.L.; Thompson, J.G.; Norman, R.J.; Robker, R.L. Beta-Oxidation Is Essential for Mouse Oocyte Developmental Competence and Early Embryo Development1. Biol. Reprod. 2010, 83, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Paczkowski, M.; Silva, E.; Schoolcraft, W.B.; Krisher, R.L. Comparative Importance of Fatty Acid Beta-Oxidation to Nuclear Maturation, Gene Expression, and Glucose Metabolism in Mouse, Bovine, and Porcine Cumulus Oocyte Complexes. Biol. Reprod. 2013, 88, 111. [Google Scholar] [CrossRef]
- Colonna, R.; Mangia, F. Mechanisms of Amino Acid Uptake in Cumulus-Enclosed Mouse Oocytes. Biol. Reprod. 1983, 28, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Zhang, S.; Gong, F.; Lu, C.; Lin, G.; Hu, L. The Mitochondrial DNA Copy Number of Cumulus Granulosa Cells May Be Related to the Maturity of Oocyte Cytoplasm. Hum. Reprod. 2020, 35, 1120–1129. [Google Scholar] [CrossRef]
- Boucret, L.; Chao de la Barca, J.M.; Moriniere, C.; Desquiret, V.; Ferre-L’Hotellier, V.; Descamps, P.; Marcaillou, C.; Reynier, P.; Procaccio, V.; May-Panloup, P. Relationship between Diminished Ovarian Reserve and Mitochondrial Biogenesis in Cumulus Cells. Hum. Reprod. 2015, 30, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Ogino, M.; Tsubamoto, H.; Sakata, K.; Oohama, N.; Hayakawa, H.; Kojima, T.; Shigeta, M.; Shibahara, H. Mitochondrial DNA Copy Number in Cumulus Cells Is a Strong Predictor of Obtaining Good-Quality Embryos after IVF. J. Assist. Reprod. Genet. 2016, 33, 367–371. [Google Scholar] [CrossRef]
- Desquiret-Dumas, V.; Clément, A.; Seegers, V.; Boucret, L.; Ferré-L’Hotellier, V.; Bouet, P.E.; Descamps, P.; Procaccio, V.; Reynier, P.; May-Panloup, P. The Mitochondrial DNA Content of Cumulus Granulosa Cells Is Linked to Embryo Quality. Hum. Reprod. 2017, 32. [Google Scholar] [CrossRef] [PubMed]
- Taugourdeau, A.; Desquiret-Dumas, V.; Hamel, J.F.; Chupin, S.; Boucret, L.; Ferré-L’Hotellier, V.; Bouet, P.E.; Descamps, P.; Procaccio, V.; Reynier, P.; et al. The Mitochondrial DNA Content of Cumulus Cells May Help Predict Embryo Implantation. J. Assist. Reprod. Genet. 2019, 36, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Stigliani, S.; Persico, L.; Lagazio, C.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA in Day 3 Embryo Culture Medium Is a Novel, Non-Invasive Biomarker of Blastocyst Potential and Implantation Outcome. MHR Basic Sci. Reprod. Med. 2014, 20, 1238–1246. [Google Scholar] [CrossRef]
- Kobayashi, M.; Kobayashi, J.; Shirasuna, K.; Iwata, H. Abundance of Cell-free Mitochondrial DNA in Spent Culture Medium Associated with Morphokinetics and Blastocyst Collapse of Expanded Blastocysts. Reprod. Med. Biol. 2020, 19, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.R.; McGillivray, B.C.; Wicker, S.M.; Peek, J.C.; Shelling, A.N.; Stone, P.; Chamley, L.W.; Cree, L.M. Characterizing Nuclear and Mitochondrial DNA in Spent Embryo Culture Media: Genetic Contamination Identified. Fertil. Steril. 2017, 107, 220–228.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D. Comparison of Mitochondrial DNA Contents in Human Embryos with Good or Poor Morphology at the 8-Cell Stage. Fertil. Steril. 2004, 81, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Spath, K.; Alfarawati, S.; Kaper, F.; Craig, A.; Michel, C.-E.; Kokocinski, F.; Cohen, J.; Munne, S.; Wells, D. Altered Levels of Mitochondrial DNA Are Associated with Female Age, Aneuploidy, and Provide an Independent Measure of Embryonic Implantation Potential. PLoS Genet. 2015, 11, e1005241. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, M.; Alfarawati, S.; Hurd, D.; Paolucci, M.; Shovelton, J.; Fragouli, E.; Wells, D. Simultaneous Assessment of Aneuploidy, Polymorphisms, and Mitochondrial DNA Content in Human Polar Bodies and Embryos with the Use of a Novel Microarray Platform. Fertil. Steril. 2014, 102, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, K.; McCaffrey, C.; Grifo, J.; Morales, A.; Perloe, M.; Munne, S.; Wells, D.; Fragouli, E. Mitochondrial DNA Quantification as a Tool for Embryo Viability Assessment: Retrospective Analysis of Data from Single Euploid Blastocyst Transfers. Hum. Reprod. 2017, 32, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Fragouli, E.; McCaffrey, C.; Ravichandran, K.; Spath, K.; Grifo, J.A.; Munné, S.; Wells, D. Clinical Implications of Mitochondrial DNA Quantification on Pregnancy Outcomes: A Blinded Prospective Non-Selection Study. Hum. Reprod. 2017, 32, 2340–2347. [Google Scholar] [CrossRef] [Green Version]
- Monnot, S.; Samuels, D.C.; Hesters, L.; Frydman, N.; Gigarel, N.; Burlet, P.; Kerbrat, V.; Lamazou, F.; Frydman, R.; Benachi, A.; et al. Mutation Dependance of the Mitochondrial DNA Copy Number in the First Stages of Human Embryogenesis. Hum. Mol. Genet. 2013, 22, 1867–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, A.R.; Brake, A.J.; Tyndall, J.C.; Griffin, D.K.; Zouves, C.G.; Barnes, F.L.; Viotti, M. Accurate Quantitation of Mitochondrial DNA Reveals Uniform Levels in Human Blastocysts Irrespective of Ploidy, Age, or Implantation Potential. Fertil. Steril. 2017, 107, 34–42.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Munck, N.; Liñán, A.; Elkhatib, I.; Bayram, A.; Arnanz, A.; Rubio, C.; Garrido, N.; Lawrenz, B.; Fatemi, H.M. MtDNA Dynamics between Cleavage-Stage Embryos and Blastocysts. J. Assist. Reprod. Genet. 2019, 36, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.T.; Sun, L.; Zhan, Y.; Marin, D.; Tao, X.; Seli, E. Mitochondrial DNA Content Is Not Predictive of Reproductive Competence in Euploid Blastocysts. Reprod. Biomed. Online 2020, 41, 183–190. [Google Scholar] [CrossRef]
- Treff, N.R.; Zhan, Y.; Tao, X.; Olcha, M.; Han, M.; Rajchel, J.; Morrison, L.; Morin, S.J.; Scott, R.T. Levels of Trophectoderm Mitochondrial DNA Do Not Predict the Reproductive Potential of Sibling Embryos. Hum. Reprod. 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.R.; Arrach, N.; Rhodes-Long, K.; Salem, W.; McGinnis, L.K.; Chung, K.; Bendikson, K.A.; Paulson, R.J.; Ahmady, A. Blastulation Timing Is Associated with Differential Mitochondrial Content in Euploid Embryos. J. Assist. Reprod. Genet. 2018, 35, 711–720. [Google Scholar] [CrossRef]
- Klimczak, A.M.; Pacheco, L.E.; Lewis, K.E.; Massahi, N.; Richards, J.P.; Kearns, W.G.; Saad, A.F.; Crochet, J.R. Embryonal Mitochondrial DNA: Relationship to Embryo Quality and Transfer Outcomes. J. Assist. Reprod. Genet. 2018, 35, 871–877. [Google Scholar] [CrossRef]
- De los Santos, M.J.; Diez Juan, A.; Mifsud, A.; Mercader, A.; Meseguer, M.; Rubio, C.; Pellicer, A. Variables Associated with Mitochondrial Copy Number in Human Blastocysts: What Can We Learn from Trophectoderm Biopsies? Fertil. Steril. 2018, 109, 110–117. [Google Scholar] [CrossRef]
- Cecchino, G.N.; Seli, E.; Alves da Motta, E.L.; García-Velasco, J.A. The Role of Mitochondrial Activity in Female Fertility and Assisted Reproductive Technologies: Overview and Current Insights. Reprod. Biomed. Online 2018, 36, 686–697. [Google Scholar] [CrossRef] [Green Version]
- Showell, M.G.; Mackenzie-Proctor, R.; Jordan, V.; Hart, R.J. Antioxidants for Female Subfertility. Cochrane Database Syst. Rev. 2020. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Luo, S.; Chavez-Badiola, A.; Liu, Z.; Yang, M.; Munne, S.; Konstantinidis, M.; Wells, D.; Huang, T. First Live Birth Using Human Oocytes Reconstituted by Spindle Nuclear Transfer for Mitochondrial DNA Mutation Causing Leigh Syndrome. Fertil. Steril. 2016, 106, e375–e376. [Google Scholar] [CrossRef]
- Zhang, J.; Zhuang, G.; Zeng, Y.; Grifo, J.; Acosta, C.; Shu, Y.; Liu, H. Pregnancy Derived from Human Zygote Pronuclear Transfer in a Patient Who Had Arrested Embryos after IVF. Reprod. Biomed. Online 2016, 33, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyre-Walker, A. Mitochondrial Replacement Therapy: Are Mito-Nuclear Interactions Likely to Be a Problem? Genetics 2017, 205, 1365–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobler, R.; Dowling, D.K.; Morrow, E.H.; Reinhardt, K. A Systematic Review and Meta-Analysis Reveals Pervasive Effects of Germline Mitochondrial Replacement on Components of Health. Hum. Reprod. Update 2018, 24, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Eyre-Walker, A. Mitochondrial Replacement and Its Effects on Human Health. Hum. Reprodu. Update 2019, 25, 392–394. [Google Scholar] [CrossRef]
- Cohen, J.; Scott, R.; Schimmel, T.; Levron, J.; Willadsen, S. Birth of Infant after Transfer of Anucleate Donor Oocyte Cytoplasm into Recipient Eggs. Lancet 1997, 350, 186–187. [Google Scholar] [CrossRef]
- Takeda, K.; Tasai, M.; Iwamoto, M.; Onishi, A.; Tagami, T.; Nirasawa, K.; Hanada, H.; Pinkert, C.A. Microinjection of Cytoplasm or Mitochondria Derived from Somatic Cells Affects Parthenogenetic Development of Murine Oocytes1. Biol. Reprod. 2005, 72, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Oktay, K.; Baltaci, V.; Sonmezer, M.; Turan, V.; Unsal, E.; Baltaci, A.; Aktuna, S.; Moy, F. Oogonial Precursor Cell-Derived Autologous Mitochondria Injection to Improve Outcomes in Women with Multiple IVF Failures Due to Low Oocyte Quality: A Clinical Translation. Reprod. Sci. 2015, 22, 1612–1617. [Google Scholar] [CrossRef]
- Labarta, E.; de los Santos, M.J.; Herraiz, S.; Escribá, M.J.; Marzal, A.; Buigues, A.; Pellicer, A. Autologous Mitochondrial Transfer as a Complementary Technique to Intracytoplasmic Sperm Injection to Improve Embryo Quality in Patients Undergoing in vitro Fertilization—A Randomized Pilot Study. Fertil. Steril. 2019, 111, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.F.; Soares, M.; Almeida Reis, S.; Ramalho-Santos, J.; Sousa, A.P.; Almeida-Santos, T. Does Supplementation with Mitochondria Improve Oocyte Competence? A Systematic Review. Reproduction 2020. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
May-Panloup, P.; Boguenet, M.; El Hachem, H.; Bouet, P.-E.; Reynier, P. Embryo and Its Mitochondria. Antioxidants 2021, 10, 139. https://doi.org/10.3390/antiox10020139
May-Panloup P, Boguenet M, El Hachem H, Bouet P-E, Reynier P. Embryo and Its Mitochondria. Antioxidants. 2021; 10(2):139. https://doi.org/10.3390/antiox10020139
Chicago/Turabian StyleMay-Panloup, Pascale, Magalie Boguenet, Hady El Hachem, Pierre-Emmanuel Bouet, and Pascal Reynier. 2021. "Embryo and Its Mitochondria" Antioxidants 10, no. 2: 139. https://doi.org/10.3390/antiox10020139
APA StyleMay-Panloup, P., Boguenet, M., El Hachem, H., Bouet, P. -E., & Reynier, P. (2021). Embryo and Its Mitochondria. Antioxidants, 10(2), 139. https://doi.org/10.3390/antiox10020139