
A Comparative Study of the Antioxidative Effects of Helichrysum italicum and Helichrysum arenarium Infusions

 ,
, 
Abstract
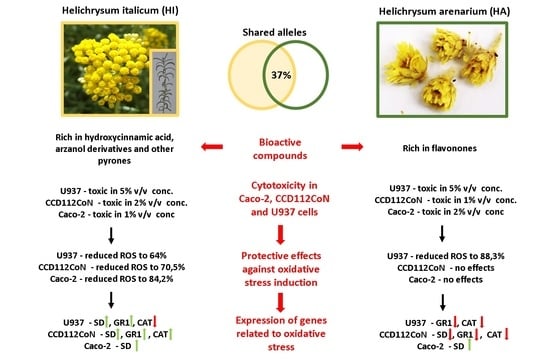
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and Microsatellite Analysis
2.3. Infusion Preparation
2.4. Chemical Analysis
2.5. Radical Scavenging Activity Assay
2.6. Cell Cultures
2.7. Cell ViabilityAassay
2.8. Intracellular ROS Level
2.9. Gene Expression Analysis
2.10. Statistical Analysis
3. Results
3.1. Plant Identification and Characterization
3.2. Radical Scavenging Activity
3.3. Cytotoxicity and the Protective Effect from Oxidative Stress Induction
3.4. The Expression of Genes Related to Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Committee on Herbal Medicinal Products (HMPC). European Union Herbal Monograph on Helichrysum arenarium (L.); Flos: Merano, Italy, 2016. [Google Scholar]
- Committee on Herbal Medicinal Products (HMPC). Assessment Report on Helichrysum arenarium (L.); Flos: Moench, Italy, 2016. [Google Scholar]
- Pljevljakušić, D.; Bigović, D.; Janković, T.; Jelačić, S.; Šavikin, K. Sandy Everlasting (Helichrysum arenarium (L.) Moench): Botanical, Chemical and Biological Properties. Front. Plant Sci. 2018, 9, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninčević, T.; Grdiša, M.; Šatović, Z.; Jug-Dujaković, M. Helichrysum italicum (Roth) G. Don: Taxonomy, Biological Activity, Biochemical and Genetic Diversity. Ind. Crop. Prod. 2019, 138, 111487. [Google Scholar] [CrossRef]
- Sala, A.; Recio, M.; Giner, R.M.; Máñez, S.; Tournier, H.; Schinella, G.; Ríos, J.-L. Anti-Inflammatory and Antioxidant Properties of Helichrysum italicum. J. Pharm. Pharmacol. 2002, 54, 365–371. [Google Scholar] [CrossRef]
- Taglialatela-Scafati, O.; Pollastro, F.; Chianese, G.; Minassi, A.; Gibbons, S.; Arunotayanun, W.; Mabebie, B.; Ballero, M.; Appendino, G. Antimicrobial Phenolics and Unusual Glycerides from Helichrysum italicum Subsp. Microphyllum. J. Nat. Prod. 2013, 76, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Garzoli, S.; Sabatino, M.; Tadić, V.; Costantini, S.; Ragno, R.; Božović, M. Chemical Composition and Antimicrobial Activity of Essential Oil of Helichrysum italicum (Roth) G. Don Fil. (Asteraceae) from Montenegro. Nat. Prod. Res. 2019, 34, 445–448. [Google Scholar] [CrossRef] [PubMed]
- D’Abrosca, B.; Buommino, E.; Caputo, P.; Scognamiglio, M.; Chambery, A.; Donnarumma, G.; Fiorentino, A. Phytochemical Study of Helichrysum italicum (Roth) G. Don: Spectroscopic Elucidation of Unusual Amino-Phlorogucinols and Antimicrobial Assessment of Secondary Metabolites from Medium-Polar Extract. Phytochemistry 2016, 132, 86–94. [Google Scholar] [CrossRef]
- Antunes Viegas, D.; Palmeira-de-Oliveira, A.; Salgueiro, L.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, R. Helichrysum italicum: From Traditional Use to Scientific Data. J. Ethnopharmacol. 2014, 151, 54–65. [Google Scholar] [CrossRef]
- Griffiths, K.; Aggarwal, B.B.; Singh, R.B.; Buttar, H.S.; Wilson, D.; De Meester, F. Food Antioxidants and Their Anti-Inflammatory Properties: A Potential Role in Cardiovascular Diseases and Cancer Prevention. Diseases 2016, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Ciulla, M.; Marinelli, L.; Cacciatore, I.; Stefano, A.D. Role of Dietary Supplements in the Management of Parkinson’s Disease. Biomolecules 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sõukand, R.; Quave, C.L.; Pieroni, A.; Pardo-de-Santayana, M.; Tardío, J.; Kalle, R.; Łuczaj, Ł.; Svanberg, I.; Kolosova, V.; Aceituno-Mata, L.; et al. Plants Used for Making Recreational Tea in Europe: A Review Based on Specific Research Sites. J. Ethnobiol. Ethnomed. 2013, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Kramberger, K.; Barlič-Maganja, D.; Bandelj, D.; Baruca Arbeiter, A.; Peeters, K.; Miklavčič Višnjevec, A.; Jenko Pražnikar, Z. HPLC-DAD-ESI-QTOF-MS Determination of Bioactive Compounds and Antioxidant Activity Comparison of the Hydroalcoholic and Water Extracts from Two Helichrysum italicum Species. Metabolites 2020, 10, 403. [Google Scholar] [CrossRef]
- Herrando Moraira, S.; Blanco Moreno, J.M.; Sáez, L.; Galbany Casals, M. Re-Evaluation of Helichrysum italicum Complex (Compositae: Gnaphalieae): A New Species from Majorca (Balearic Islands). Collect. Bot. 2016, 35, e009. [Google Scholar]
- Baruca Arbeiter, A.; Hladnik, M.; Jakše, J.; Bandelj, D. First Set of Microsatellite Markers for Immortelle (Helichrysum italicum (Roth) G. Don): A Step towards the Selection of the Most Promising Genotypes for Cultivation. Ind. Crop. Prod. 2021, 162, 113298. [Google Scholar] [CrossRef]
- Schuelke, M. An Economic Method for the Fluorescent Labeling of PCR Fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-F.; Wang, Y.-K.; Yih, K.-H. DPPH Free-radical Scavenging Ability, Total Phenolic Content, and Chemical Composition Analysis of Forty-five Kinds of Essential Oils. Int. J. Cosmet. Sci. 2009, 31, 475–476. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A Resource of Human and Mouse PCR Primer Pairs for Gene Expression Detection and Quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.G.; Barreira, L.; Bijttebier, S.; Pieters, L.; Neves, V.; Rodrigues, M.J.; Rivas, R.; Varela, J.; Custódio, L. Chemical Profiling of Infusions and Decoctions of Helichrysum italicum Subsp. Picardii by UHPLC-PDA-MS and in Vitro Biological Activities Comparatively with Green Tea (Camellia sinensis) and Rooibos Tisane (Aspalathus linearis). J. Pharm. Biomed. Anal. 2017, 145, 593–603. [Google Scholar] [CrossRef]
- Galbany-Casals, M.; Blanco-Moreno, J.M.; Garcia-Jacas, N.; Breitwieser, I.; Smissen, R.D. Genetic Variation in Mediterranean Helichrysum italicum (Asteraceae; Gnaphalieae): Do Disjunct Populations of Subsp. Microphyllum Have a Common Origin? Plant Biol. 2011, 13, 678–687. [Google Scholar] [CrossRef]
- Rosa, A.; Pollastro, F.; Atzeri, A.; Appendino, G.; Melis, M.P.; Deiana, M.; Incani, A.; Loru, D.; Dessì, M.A. Protective Role of Arzanol against Lipid Peroxidation in Biological Systems. Chem. Phys. Lipids 2011, 164, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Spagnol, C.M.; Assis, R.P.; Brunetti, I.L.; Isaac, V.L.B.; Salgado, H.R.N.; Corrêa, M.A. In Vitro Methods to Determine the Antioxidant Activity of Caffeic Acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 219, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Jeng, T.L.; Lai, C.C.; Liao, T.C.; Lin, S.Y.; Sung, J.M. Effects of Drying on Caffeoylquinic Acid Derivative Content and Antioxidant Capacity of Sweet Potato Leaves. J. Food Drug Anal. 2015, 23, 701–708. [Google Scholar] [CrossRef]
- D’Andrea, G. Quercetin: A Flavonol with Multifaceted Therapeutic Applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- Gradinaru, A.C.; Silion, M.; Trifan, A.; Miron, A.; Aprotosoaie, A.C. Helichrysum arenarium Subsp. Arenarium: Phenolic Composition and Antibacterial Activity against Lower Respiratory Tract Pathogens. Nat. Prod. Res. 2014, 28, 2076–2080. [Google Scholar] [CrossRef]
- Czinner, E.; Kéry, A.; Hagymási, K.; Blázovics, A.; Lugasi, A.; Szöke, E.; Lemberkovics, E. Biologically Active Compounds of Helichrysum arenarium (L.) Moench. Eur. J. Drug Metab. Pharmacokinet. 1999, 24, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Kuta, I.; Cisowski, W.; Dryś, A. Antiradical Activity of Hydrolyzed and Non-Hydrolyzed Extracts from Helichrysi Inflorescentia and Its Phenolic Contents. Z. Nat. C 2004, 59, 363–367. [Google Scholar] [CrossRef]
- Mari, A.; Napolitano, A.; Masullo, M.; Pizza, C.; Piacente, S. Identification and Quantitative Determination of the Polar Constituents in Helichrysum italicum Flowers and Derived Food Supplements. J. Pharm. Biomed. Anal. 2014, 96, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.; Ebrahim, W.; Özkaya, F.C.; Mándi, A.; Kurtán, T.; El-Neketi, M.; Liu, Z.; Proksch, P. Pyrone Derivatives from Helichrysum italicum. Fitoterapia 2019, 133, 80–84. [Google Scholar] [CrossRef]
- Maksimovic, S.; Tadic, V.; Skala, D.; Zizovic, I. Separation of Phytochemicals from Helichrysum italicum: An Analysis of Different Isolation Techniques and Biological Activity of Prepared Extracts. Phytochemistry 2017, 138, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; La Rocca, A.; Marsili, S.; Mariotti, M.G. Traditional Uses of Plants in the Eastern Riviera (Liguria, Italy). J. Ethnopharmacol. 2009, 125, 16–30. [Google Scholar] [CrossRef]
- Pieroni, A. Medicinal Plants and Food Medicines in the Folk Traditions of the Upper Lucca Province, Italy. J. Ethnopharmacol. 2000, 70, 235–273. [Google Scholar] [CrossRef]
- Bruni, A.; Ballero, M.; Poli, F. Quantitative Ethnopharmacological Study of the Campidano Valley and Urzulei District, Sardinia, Italy. J. Ethnopharmacol. 1997, 57, 97–124. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Ros, R.; Cayssials, V.; Jenab, M.; Rothwell, J.A.; Fedirko, V.; Aleksandrova, K.; Tjønneland, A.; Kyrø, C.; Overvad, K.; Boutron-Ruault, M.-C.; et al. Dietary Intake of Total Polyphenol and Polyphenol Classes and the Risk of Colorectal Cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) Cohort. Eur. J. Epidemiol. 2018, 33, 1063–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apaydin Yildirim, B.; Kordali, S.; Terim Kapakin, K.A.; Yildirim, F.; Aktas Senocak, E.; Altun, S. Effect of Helichrysum plicatum DC. Subsp. Plicatum Ethanol Extract on Gentamicin-Induced Nephrotoxicity in Rats. J. Zhejiang Univ. Sci. B 2017, 18, 501–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, M.P. Hormesis and Disease Resistance: Activation of Cellular Stress Response Pathways. Hum. Exp. Toxicol. 2008, 27, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Santoro, A.; Monti, D.; Crupi, R.; Di Paola, R.; Latteri, S.; Cuzzocrea, S.; Zappia, M.; Giordano, J.; Calabrese, E.J.; et al. Aging and Parkinson’s Disease: Inflammaging, Neuroinflammation and Biological Remodeling as Key Factors in Pathogenesis. Free Radic. Biol. Med. 2018, 115, 80–91. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Calabrese, V.; Tsatsakis, A.; Giordano, J.J. Hormesis and Ginkgo Biloba (GB): Numerous Biological Effects of GB Are Mediated via Hormesis. Ageing Res. Rev. 2020, 64, 101019. [Google Scholar] [CrossRef] [PubMed]
- Ping, Z.; Liu, W.; Kang, Z.; Cai, J.; Wang, Q.; Cheng, N.; Wang, S.; Wang, S.; Zhang, J.H.; Sun, X. Sulforaphane Protects Brains against Hypoxic–Ischemic Injury through Induction of Nrf2-Dependent Phase 2 Enzyme. Brain Res. 2010, 1343, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Scuto, M.; Fusco, R.; Trovato, A.; Ontario, M.L.; Crea, R.; Di Paola, R.; Cuzzocrea, S.; Calabrese, V. Anti-Inflammatory and Anti-Oxidant Activity of Hidrox® in Rotenone-Induced Parkinson’s Disease in Mice. Antioxidants 2020, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.; Ojcius, D.M.; Ko, Y.-F.; Ke, P.-Y.; Wu, C.-Y.; Peng, H.-H.; Young, J.D. Hormetic Effects of Phytochemicals on Health and Longevity. Trends Endocrinol. Metab. 2019, 30, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Duda, S.C. Changes in Major Bioactive Compounds with Antioxidant Activity of Agastache Foeniculum, Lavandula Angustifolia, Melissa Officinalis and Nepeta Cataria: Effect of Harvest Time and Plant Species. Ind. Crop. Prod. 2015, 77, 499–507. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Infusion | ||
|---|---|---|---|
| HIa | HIb | HA | |
| Total Identified Phenolic Compounds 1 | 2071068 | 974921 | 681606 |
| Total hydroxycinnamic acids | 1104310 | 472357 | 189173 |
| Caffeic acid and its derivatives | 174492 | 218381 | 31144 |
| Other hydroxycinnamic acids and their derivatives | 43618 | 44407 | 0 |
| Caffeoylquinic acids | 443858 | 197797 | 150534 |
| Other monoesters | 59450 | 0 | 4167 |
| Dicaffeoylquinic acids | 382892 | 11772 | 3328 |
| Total hydroxybenzoic acids | 89445 | 92466 | 64752 |
| Monohydroxybenzoic acids | 43026 | 72816 | 42956 |
| Dihydroxybenzoic acids | 46419 | 19650 | 21796 |
| Total flavonoids | 92004 | 24180 | 227018 |
| Quercetin derivatives | 25573 | 0 | 0 |
| Myricetin derivatives | 10056 | 5223 | 0 |
| Kaempferol derivatives | 47970 | 0 | 0 |
| Flavanones | 0 | 18957 | 227018 |
| Flavones | 8405 | 0 | 0 |
| Total arzanol derivatives and other pyrones | 392448 | 275723 | 7521 |
| Pyrones | 384837 | 256991 | 0 |
| Arzanol and its derivatives | 7611 | 18732 | 7521 |
| Total other phenolic compounds | 392861 | 110195 | 193142 |
| Cyclic polyols | 180368 | 54374 | 168071 |
| Isobenzofuranones | 156472 | 4638 | 22703 |
| Neolignans | 48243 | 15502 | 2368 |
| Acetophenones | 0 | 3750 | 0 |
| Tremetones Coumarins | 0 7778 | 14525 17406 | 0 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kramberger, K.; Jenko Pražnikar, Z.; Baruca Arbeiter, A.; Petelin, A.; Bandelj, D.; Kenig, S. A Comparative Study of the Antioxidative Effects of Helichrysum italicum and Helichrysum arenarium Infusions. Antioxidants 2021, 10, 380. https://doi.org/10.3390/antiox10030380
Kramberger K, Jenko Pražnikar Z, Baruca Arbeiter A, Petelin A, Bandelj D, Kenig S. A Comparative Study of the Antioxidative Effects of Helichrysum italicum and Helichrysum arenarium Infusions. Antioxidants. 2021; 10(3):380. https://doi.org/10.3390/antiox10030380
Chicago/Turabian StyleKramberger, Katja, Zala Jenko Pražnikar, Alenka Baruca Arbeiter, Ana Petelin, Dunja Bandelj, and Saša Kenig. 2021. "A Comparative Study of the Antioxidative Effects of Helichrysum italicum and Helichrysum arenarium Infusions" Antioxidants 10, no. 3: 380. https://doi.org/10.3390/antiox10030380
APA StyleKramberger, K., Jenko Pražnikar, Z., Baruca Arbeiter, A., Petelin, A., Bandelj, D., & Kenig, S. (2021). A Comparative Study of the Antioxidative Effects of Helichrysum italicum and Helichrysum arenarium Infusions. Antioxidants, 10(3), 380. https://doi.org/10.3390/antiox10030380