
Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Treatments
2.3. Detection of MG-H1 Protein Adducts
2.4. Protein Extraction
2.5. Glyoxalase 1 (Glo1) Enzyme Activity and Protein Assay
2.6. H2O2 and MDA Detection
2.7. GSH Detection
2.8. Cell Proliferation Assay
2.9. Apoptosis Detection
2.10. IL-1β, IL-6 and IL-8 Detection
2.11. Nrf2, HO-1, Hsp70, Trx and γ-GCS Detection
2.12. RNA Isolation, Reverse Transcription, and Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) Analyses
2.13. Preparation of DPSCs Conditioned Medium
2.14. In Vitro Tubule Like-Formation and Stabilization Assay
2.15. Statistical Analysis
3. Results
3.1. MG Induces Glycative Stress in DPSCs
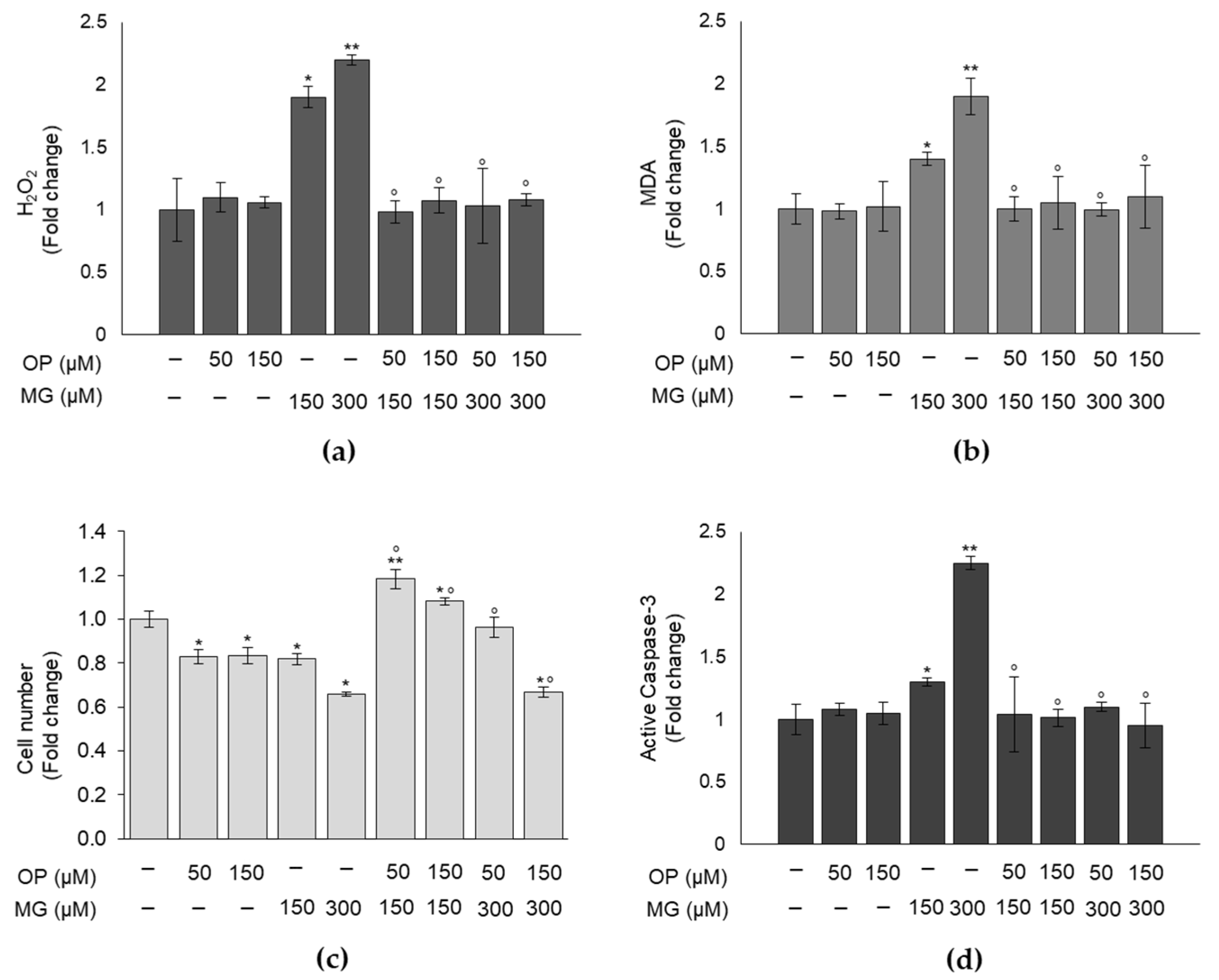
3.2. MG Induces Oxidative Stress in DPSCs and Impairs Cell Growth
3.3. OP Prevents MG-Dependent Dicarbonyl Stress in DPSCs
3.4. OP Prevents MG-Dependent Oxidative Stress and Cell Growth Impairment in DPSCs
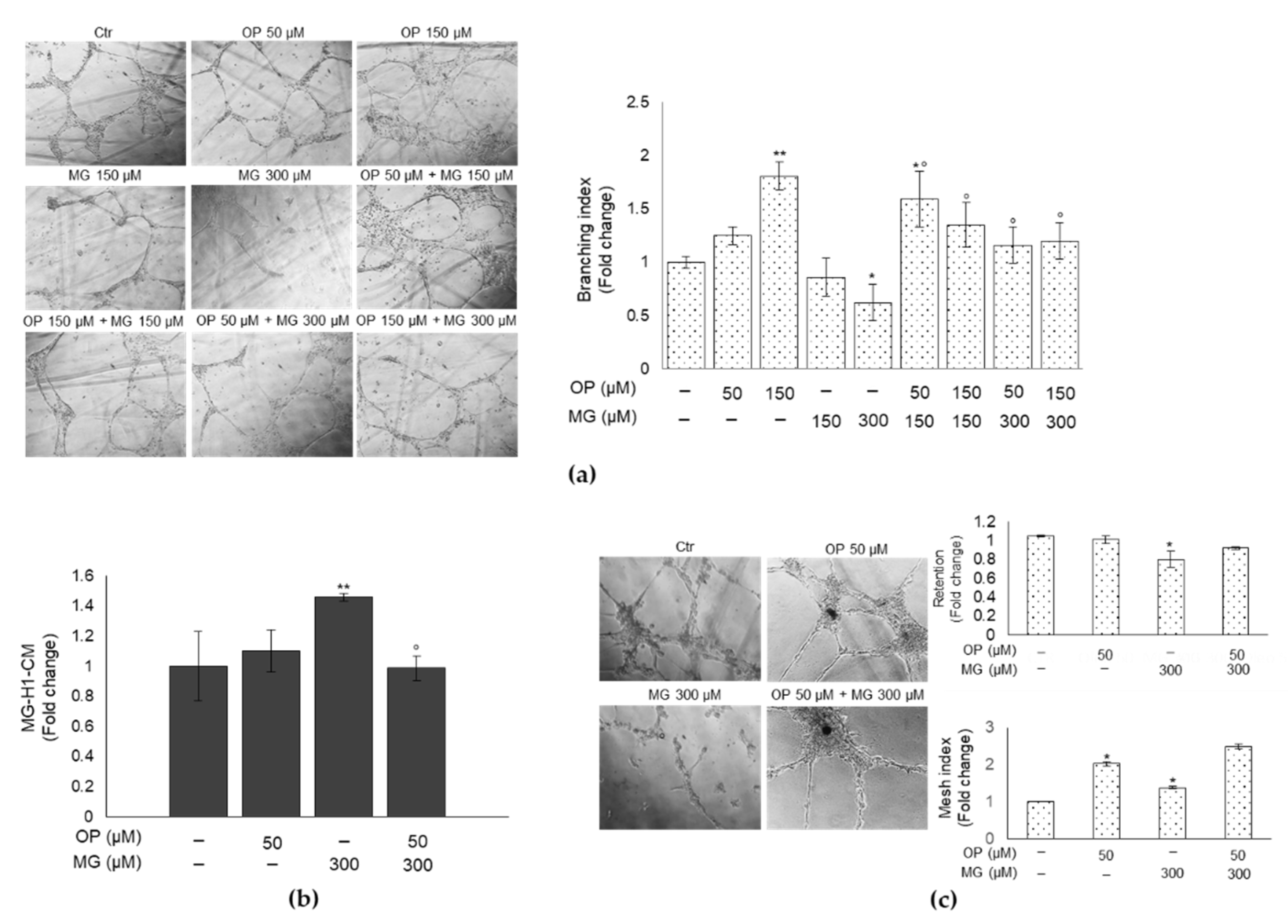
3.5. OP Prevents DPSCs Exposed to MG from Affecting Tube Formation of HUVECs
3.6. OP Partially Protects DPSCs from MG-Induced Inflammation
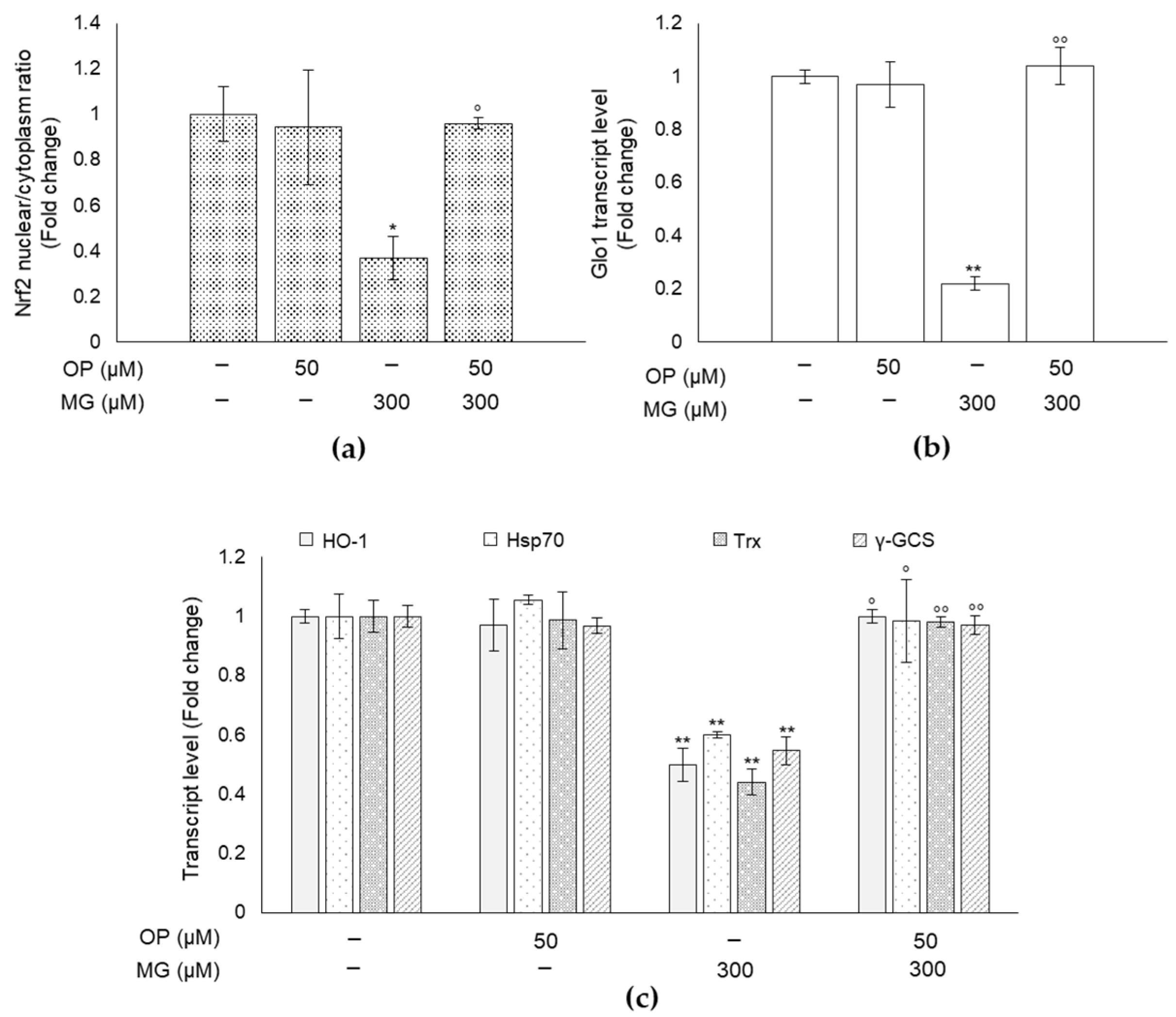
3.7. MG-Induced Responses and Their Prevention by OP Are Paralleled by Changes of the Master Redox-Sensitive Transcriptional Regulator Nrf2 in DPSCs
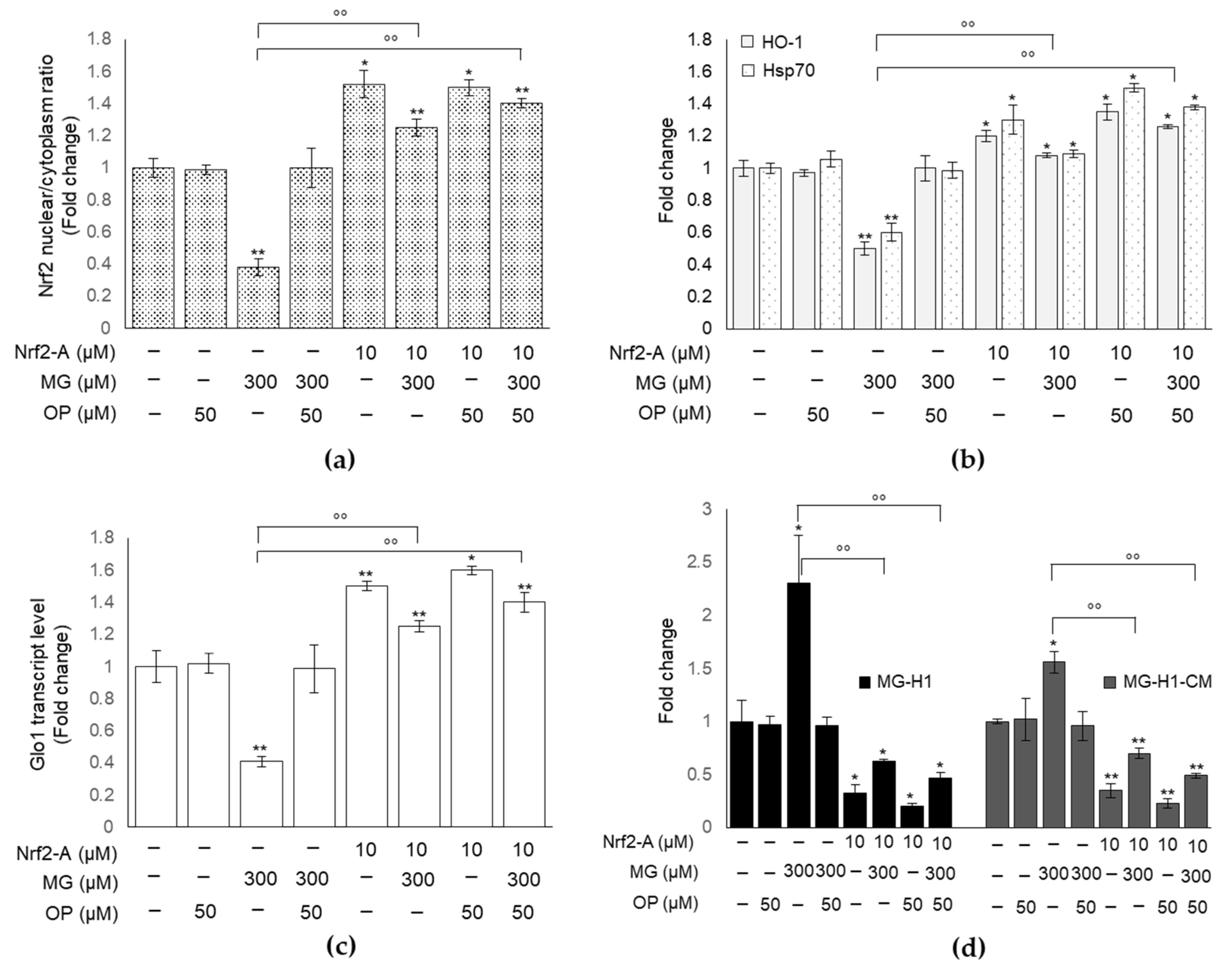
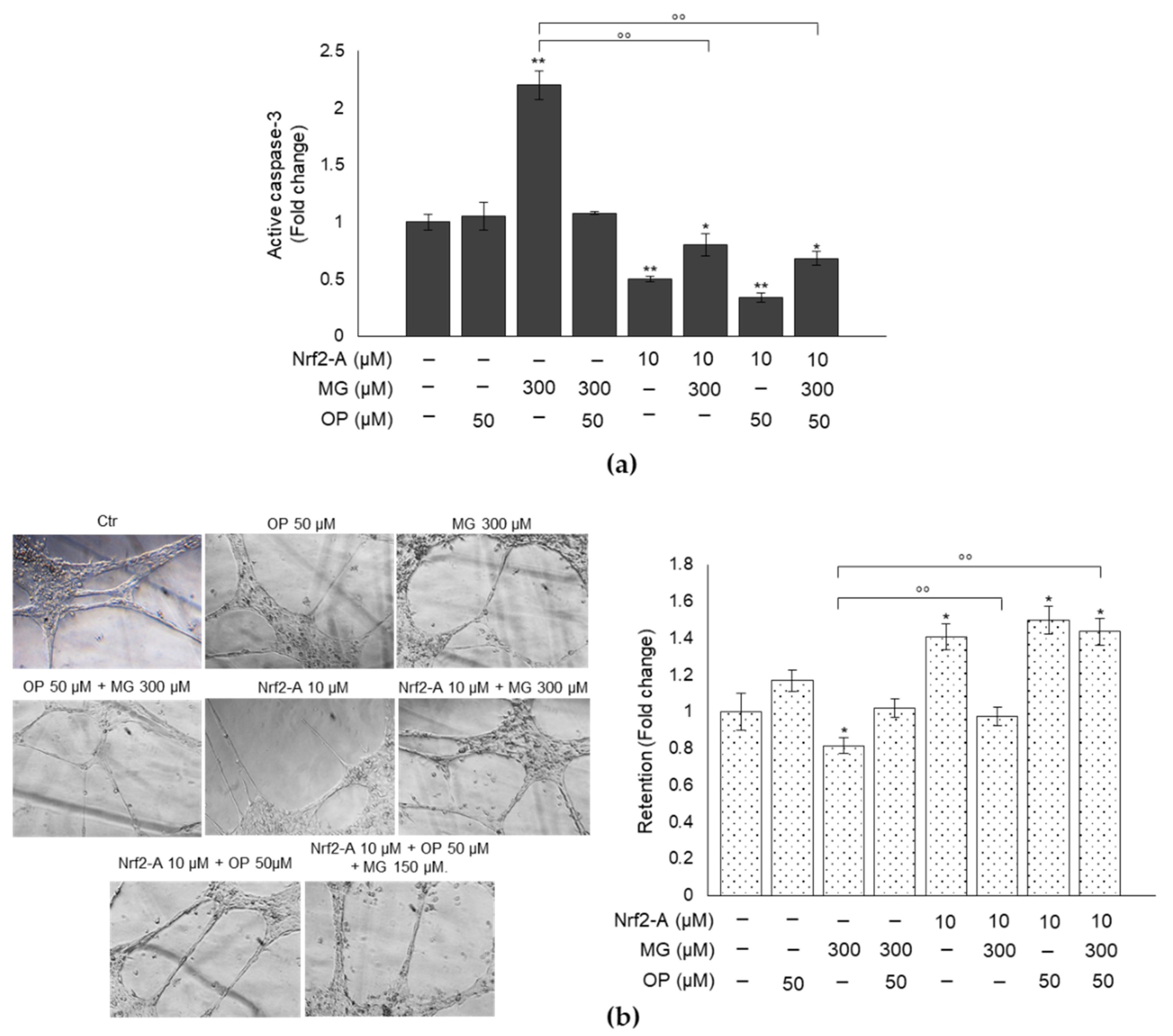
3.8. Pharmacological Activation of Nrf2 Confirms the Role of This Transcription Factor in MG-Induced Responses and Their Prevention by OP Through Nrf2/Glo1 Axis in DPSCs
3.9. MG-Induced Responses and Their Prevention by OP Are Paralleled by Changes of GSH in DPSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antognelli, C.; Talesa, V.N. Glyoxalases in Urological Malignancies. Int. J. Mol. Sci. 2018, 19, 415. [Google Scholar] [CrossRef] [Green Version]
- Marinucci, L.; Balloni, S.; Fettucciari, K.; Bodo, M.; Talesa, V.N.; Antognelli, C. Nicotine Induces Apoptosis in Human Osteoblasts via a Novel Mechanism Driven by H2O2 and Entailing Glyoxalase 1–Dependent MG-H1 Accumulation Leading to TG2–Mediated NF-kB Desensitization: Implication for Smokers-Related Osteoporosis. Free Radic. Biol. Med. 2018, 117, 6–17. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Nakamura, N.; Sotokawauchi, A.; Higashimoto, Y.; Yamagishi, S.I. Methylglyoxal-Derived Hydroimidazolone–1 Evokes Inflammatory Reactions in Endothelial Cells via an Interaction with Receptor for Advanced Glycation End Products. Diab. Vas. Dis. Res. 2017, 14, 450–453. [Google Scholar] [CrossRef]
- Antognelli, C.; Gambelunghe, A.; Muzi, G.; Talesa, V.N. Peroxynitrite-Mediated Glyoxalase I Epigenetic Inhibition Drives Apoptosis in Airway Epithelial Cells Exposed to Crystalline Silica via a Novel Mechanism Involving Argpyrimidine-Modified Hsp70, JNK, and NF-Kappab. Free Radic. Biol. Med. 2015, 84, 128–141. [Google Scholar] [CrossRef]
- Antognelli, C.; Mancuso, F.; Frosini, R.; Arato, I.; Calvitti, M.; Calafiore, R.; Talesa, V.N.; Luca, G. Testosterone and Follicle Stimulating Hormone-Dependent Glyoxalase 1 Up-Regulation Sustains the Viability of Porcine Sertoli Cells through the Control of Hydroimidazolone–and Argpyrimidine-Mediated NF-kappaB Pathway. Am. J. Pathol. 2018, 188, 2553–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antognelli, C.; Gambelunghe, A.; Muzi, G.; Talesa, V.N. Glyoxalase I Drives Epithelial-to-Mesenchymal Transition via Argpyrimidine-Modified Hsp70, Mir-21 and SMAD Signalling in Human Bronchial Cells BEAS-2B Chronically Exposed to Crystalline Silica Min-U-Sil 5: Transformation into a Neoplastic-Like Phenotype. Free Radic. Biol. Med. 2016, 92, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Cecchetti, R.; Riuzzi, F.; Peirce, M.J.; Talesa, V.N. Glyoxalase 1 Sustains the Metastatic Phenotype of Prostate Cancer Cells via EMT Control. J. Cell. Mol. Med. 2018, 22, 2865–2883. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Mezzasoma, L.; Mearini, E.; Talesa, V.N. Glyoxalase 1-419C>A Variant Is Associated with Oxidative Stress: Implications in Prostate Cancer Progression. PLoS ONE 2013, 8, e74014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antognelli, C.; Moretti, S.; Frosini, R.; Puxeddu, E.; Sidoni, A.; Talesa, V.N. Methylglyoxal Acts as a Tumor-Promoting Factor in Anaplastic Thyroid Cancer. Cells 2019, 8, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatone, C.; Carbone, M.C.; Campanella, G.; Festuccia, C.; Artini, P.G.; Talesa, V.; Focarelli, R.; Amicarelli, F. Female Reproductive Dysfunction During Ageing: Role of Methylglyoxal in the Formation of Advanced Glycation Endproducts in Ovaries of Reproductively-Aged Mice. J. Biol. Regul. Homeost. Agents 2010, 24, 63–72. [Google Scholar] [PubMed]
- Lee, K.M.; Lee, C.Y.; Zhang, G.; Lyu, A.; Yue, K.K.M. Methylglyoxal Activates Osteoclasts through JNK Pathway Leading to Osteoporosis. Chem. Biol. Interact. 2019, 308, 147–154. [Google Scholar] [CrossRef]
- De Felice, F.; Megiorni, F.; Pietrantoni, I.; Tini, P.; Lessiani, G.; Mastroiacovo, D.; Mattana, P.; Antinozzi, C.; Di Luigi, L.; Delle Monache, S.; et al. Sulodexide Counteracts Endothelial Dysfunction Induced by Metabolic or Non-Metabolic Stresses through Activation of the Autophagic Program. Eur. Rev. Med. Pharm. Sci. 2019, 23, 2669–2680. [Google Scholar] [CrossRef]
- Allaman, I.; Belanger, M.; Magistretti, P.J. Methylglyoxal, the Dark Side of Glycolysis. Front. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Jagt, D.L.V. Methylglyoxal, Diabetes Mellitus and Diabetic Complications. Drug Metab. Drug Interact. 2008, 23, 93–124. [Google Scholar] [CrossRef] [PubMed]
- Zunkel, K.; Simm, A.; Bartling, B. Long-Term Intake of the Reactive Metabolite Methylglyoxal Is Not Toxic in Mice. Food Chem. Toxicol. 2020, 141, 111333. [Google Scholar] [CrossRef]
- Pfeifer, Y.V.; Haase, P.T.; Kroh, L.W. Reactivity of Thermally Treated Alpha-Dicarbonyl Compounds. J. Agric. Food Chem. 2013, 61, 3090–3096. [Google Scholar] [CrossRef] [PubMed]
- Ashour, A.; Xue, M.; Al-Motawa, M.; Thornalley, P.J.; Rabbani, N. Glycolytic Overload-Driven Dysfunction of Periodontal Ligament Fibroblasts in High Glucose Concentration, Corrected by Glyoxalase 1 Inducer. BMJ Open Diabetes Res. Care 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Retamal, I.N.; Hernandez, R.; Gonzalez-Rivas, C.; Caceres, M.; Arancibia, R.; Romero, A.; Martinez, C.; Tobar, N.; Martinez, J.; Smith, P.C. Methylglyoxal and Methylglyoxal-Modified Collagen as Inducers of Cellular Injury in Gingival Connective Tissue Cells. J. Periodontal Res. 2016, 51, 812–821. [Google Scholar] [CrossRef]
- Kashket, S.; Maiden, M.F.; Haffajee, A.D.; Kashket, E.R. Accumulation of Methylglyoxal in the Gingival Crevicular Fluid of Chronic Periodontitis Patients. J. Clin. Periodontol. 2003, 30, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Settem, R.P.; Honma, K.; Shankar, M.; Li, M.; LaMonte, M.; Xu, D.; Genco, R.J.; Browne, R.W.; Sharma, A. Tannerella Forsythia-Produced Methylglyoxal Causes Accumulation of Advanced Glycation Endproducts to Trigger Cytokine Secretion in Human Monocytes. Mol. Oral Microbiol. 2018, 33, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Maasen, K.; Scheijen, J.L.; Opperhuizen, A.; Stehouwer, C.D.A.; Van Greevenbroek, M.M.; Schalkwijk, C.G. Quantification of Dicarbonyl Compounds in Commonly Consumed Foods and Drinks; Presentation of a Food Composition Database for Dicarbonyls. Food Chem. 2021, 339, 128063. [Google Scholar] [CrossRef]
- Kwak, S.; Choi, Y.S.; Na, H.G.; Bae, C.H.; Song, S.Y.; Kim, Y.D. Glyoxal and Methylglyoxal as E-cigarette Vapor Ingredients-Induced Pro-Inflammatory Cytokine and Mucins Expression in Human Nasal Epithelial Cells. Am. J. Rhinol. Allergy 2021, 35, 213–220. [Google Scholar] [CrossRef]
- Azimi, P.; Keshavarz, Z.; Luna, M.L.; Laurent, J.G.C.; Vallarino, J.; Christiani, D.C.; Allen, J.G. An Unrecognized Hazard in E-Cigarette Vapor: Preliminary Quantification of Methylglyoxal Formation from Propylene Glycol in E-Cigarettes. Int. J. Environ. Res. Public Health 2021, 18, 385. [Google Scholar] [CrossRef] [PubMed]
- SriChinthu, K.K.; Pavithra, V.; Kumar, G.S.; Prasad, H.; Prema, P.; Yoithapprabhunath, T.R.; Rangarajan, N. Evaluation of Gingival and Periodontal Status in Obese and Non-Obese Type II Diabetic Patients–A Cross Sectional Study. Med. Pharm. Rep. 2021, 94, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.T.M.; Akhter, R.; Garde, S.; Scott, C.; Twigg, S.M.; Colagiuri, S.; Ajwani, S.; Eberhard, J. The Association of Periodontal Disease with the Complications of Diabetes Mellitus. A Systematic Review. Diabetes Res. Clin. Pract. 2020, 165, 108244. [Google Scholar] [CrossRef] [PubMed]
- Delle Monache, S.; Martellucci, S.; Clementi, L.; Pulcini, F.; Santilli, F.; Mei, C.; Piccoli, L.; Angelucci, A.; Mattei, V. In Vitro Conditioning Determines the Capacity of Dental Pulp Stem Cells to Function as Pericyte-Like Cells. Stem Cells Dev. 2019, 28, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Tomokiyo, A.; Hasegawa, D.; Hamano, S.; Sugii, H.; Maeda, H. Insight into the Role of Dental Pulp Stem Cells in Regenerative Therapy. Biology 2020, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T. Pulp and Dentin Tissue Engineering and Regeneration: Current Progress. Regen. Med. 2009, 4, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, N. Characterisation of Dental Pulp Stem Cells: A New Horizon for Tissue Regeneration? Arch. Oral Biol. 2012, 57, 1439–1458. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Khan, A.S.; Khan, N.A.; Hichami, A. Antioxidant and Anti-Inflammatory Potential of Polyphenols Contained in Mediterranean Diet in Obesity: Molecular Mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Navarro, M.; Morales, F.J.; Ramos, S. Olive Leaf Extract Concentrated in Hydroxytyrosol Attenuates Protein Carbonylation and the Formation of Advanced Glycation End Products in a Hepatic Cell Line (Hepg2). Food Funct. 2017, 8, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Antognelli, C.; Gambelunghe, A.; Del Buono, C.; Murgia, N.; Talesa, V.N.; Muzi, G. Crystalline Silica Min-U-Sil 5 Induces Oxidative Stress in Human Bronchial Epithelial Cells BEAS-2B by Reducing the Efficiency of Antiglycation and Antioxidant Enzymatic Defenses. Chem. Biol. Interact. 2009, 182, 13–21. [Google Scholar] [CrossRef]
- Delle Monache, S.; Calgani, A.; Sanita, P.; Zazzeroni, F.; Warschauer, E.G.; Giuliani, A.; Amicucci, G.; Angelucci, A. Adipose-Derived Stem Cells Sustain Prolonged Angiogenesis Through Leptin Secretion. Growth Factors 2016, 34, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, H.; Aissa, I.; Slim, C.; Boujbiha, M.A.; Zaouali, M.A.; Bejaoui, M.; Wilke, V.; Ben Jannet, H.; Mosbah, H.; Ben Abdennebi, H. Effect of Oleuropein on Oxidative Stress, Inflammation and Apoptosis Induced by Ischemia-Reperfusion Injury in Rat Kidney. Life Sci. 2020, 255, 117833. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Wu, J.; Lu, W.; Xu, W.; Wu, B. Dental Pulp Stem Cells from Human Teeth with Deep Caries Displayed an Enhanced Angiogenesis Potential In Vitro. J. Dent. Sci. 2021, 16, 318–326. [Google Scholar] [CrossRef]
- Lee, J.H.; Parveen, A.; Do, M.H.; Kang, M.C.; Yumnam, S.; Kim, S.Y. Molecular Mechanisms of Methylglyoxal-Induced Aortic Endothelial Dysfunction in Human Vascular Endothelial Cells. Cell Death Dis. 2020, 11, 403. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) Signaling in Oxidative Stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambelunghe, A.; Giovagnoli, S.; Di Michele, A.; Boncompagni, S.; Dell’Omo, M.; Leopold, K.; Iavicoli, I.; Talesa, V.N.; Antognelli, C. Redox-Sensitive Glyoxalase 1 Up-Regulation Is Crucial for Protecting Human Lung Cells from Gold Nanoparticles Toxicity. Antioxidants 2020, 9, 697. [Google Scholar] [CrossRef]
- Antognelli, C.; Trapani, E.; Delle Monache, S.; Perrelli, A.; Daga, M.; Pizzimenti, S.; Barrera, G.; Cassoni, P.; Angelucci, A.; Trabalzini, L.; et al. KRIT1 Loss-of-Function Induces a Chronic Nrf2-Mediated Adaptive Homeostasis that Sensitizes Cells to Oxidative Stress: Implication for Cerebral Cavernous Malformation Disease. Free Radic. Biol. Med. 2018, 115, 202–218. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Kumar, S.; Hassan, M.; Wu, H.; Thimmulappa, R.K.; Kumar, A.; Sharma, S.K.; Parmar, V.S.; Biswal, S.; Malhotra, S.V. Novel Chalcone Derivatives as Potent Nrf2 Activators in Mice and Human Lung Epithelial Cells. J. Med. Chem. 2011, 54, 4147–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suantawee, T.; Thilavech, T.; Cheng, H.; Adisakwattana, S. Cyanidin Attenuates Methylglyoxal-Induced Oxidative Stress and Apoptosis in INS-1 Pancreatic β-Cells by Increasing Glyoxalase-1 Activity. Nutrients 2020, 12, 1319. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.S.; Chon, S.; Jung, W.W.; Choi, E.M. Crocin Attenuates Methylglyoxal-Induced Osteoclast Dysfunction by Regulating Glyoxalase, Oxidative Stress, and Mitochondrial Function. Food Chem. Toxicol. 2019, 124, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Liu, L.; Zhou, S.; Zhu, M.; Wang, B. N-acetylcysteine Inhibits Atherosclerosis by Correcting Glutathionedependent Methylglyoxal Elimination and Dicarbonyl/Oxidative Stress in the Aorta of Diabetic Mice. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef]
- Casagrande, L.; Cordeiro, M.M.; Nor, S.A.; Nor, J.E. Dental Pulp Stem Cells in Regenerative Dentistry. Odontology 2011, 99, 1–7. [Google Scholar] [CrossRef]
- Zhong, T.Y.; Zhang, Z.C.; Gao, Y.N.; Lu, Z.; Qiao, H.; Zhou, H.; Liu, Y. Loss of Wnt4 expression Inhibits the Odontogenic Potential of Dental Pulp Stem Cells through JNK Signaling in Pulpitis. Am. J. Transl. Res. 2019, 11, 1819–1826. [Google Scholar]
- Tatullo, M.; Marrelli, M.; Shakesheff, K.M.; White, L.J. Dental Pulp Stem Cells: Function, Isolation and Applications in Regenerative Medicine. J. Tissue Eng. Regen. Med. 2015, 9, 1205–1216. [Google Scholar] [CrossRef]
- Bakopoulou, A.; About, I. Stem Cells of Dental Origin: Current Research Trends and Key Milestones towards Clinical Application. Stem Cells Int. 2016, 2016, 4209891. [Google Scholar] [CrossRef] [Green Version]
- Hanssen, N.M.J.; Stehouwer, C.D.A.; Schalkwijk, C.G. Methylglyoxal Stress, the Glyoxalase System, and Diabetic Chronic Kidney Disease. Curr. Opin. Nephrol. Hypertens. 2019, 28, 26–33. [Google Scholar] [CrossRef]
- López-López, J.; Jané-Salas, E.; Estrugo-Devesa, A.; Velasco-Ortega, E.; Martín-González, J.; Segura-Egea, J.J. Periapical and Endodontic Status of Type 2 Diabetic Patients in Catalonia, Spain: A Cross-Sectional Study. J. Endod. 2011, 37, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Marotta, P.S.; Fontes, T.V.; Armada, L.; Lima, K.C.; Rocas, I.N.; Siqueira, J.F. Type 2 Diabetes Mellitus and the Prevalence of Apical Periodontitis and Endodontic Treatment in an Adult Brazilian Population. J. Endod. 2012, 38, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Martellucci, S.; Santacroce, C.; Santilli, F.; Piccoli, L.; Delle Monache, S.; Angelucci, A.; Misasi, R.; Sorice, M.; Mattei, V. Cellular and Molecular Mechanisms Mediated by recPrPC Involved in the Neuronal Differentiation Process of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 345. [Google Scholar] [CrossRef] [Green Version]
- Cannataro, R.; Fazio, A.; La Torre, C.; Caroleo, M.C.; Cione, E. Polyphenols in the Mediterranean Diet: From Dietary Sources to microRNA Modulation. Antioxidants 2021, 10, 328. [Google Scholar] [CrossRef]
- Tsoumani, M.; Georgoulis, A.; Nikolaou, P.E.; Kostopoulos, I.V.; Dermintzoglou, T.; Papatheodorou, I.; Zoga, A.; Efentakis, P.; Konstantinou, M.; Gikas, E.; et al. Acute Administration of the Olive Constituent, Oleuropein, Combined with Ischemic Postconditioning Increases Myocardial Protection by Modulating Oxidative Defense. Free Radic. Biol. Med. 2021, 166, 18–32. [Google Scholar] [CrossRef]
- Motawea, M.H.; Elmaksoud, H.A.A.; Elharrif, M.G.; Desoky, A.A.E.; Ibrahimi, A. Evaluation of Anti-inflammatory and Antioxidant Profile of Oleuropein in Experimentally Induced Ulcerative Colitis. Int. J. Mol. Cell. Med. 2020, 9, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Hilkens, P.; Fanton, Y.; Martens, W.; Gervois, P.; Struys, T.; Politis, C.; Lambrichts, I.; Bronckaers, A. Pro-Angiogenic Impact of Dental Stem Cells In Vitro and In Vivo. Stem Cell Res. 2014, 12, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Mattei, V.; Martellucci, S.; Pulcini, F.; Santilli, F.; Sorice, M. Delle Monache, S. Regenerative Potential of DPSCs and Revascularization: Direct, Paracrine or Autocrine Effect? Stem Cell Rev. Rep. 2021, 1–12. [Google Scholar] [CrossRef]
- Rasheed, Z.; Akhtar, N.; Haqqi, T.M. Advanced Glycation End Products Induce the Expression of Interleukin-6 and Interleukin-8 by Receptor for Advanced Glycation End Product-Mediated Activation of Mitogen-Activated Protein Kinases and Nuclear Factor-Kappab in Human Osteoarthritis Chondrocytes. Rheumatology 2011, 50, 838–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.H.; Hwang, Y.; Heo, S.J.; Jun, H.S. Diphlorethohydroxycarmalol Attenuates Methylglyoxal-Induced Oxidative Stress and Advanced Glycation End Product Formation in Human Kidney Cells. Oxid. Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Vomhof-Dekrey, E.E.; Picklo, M.J. The Nrf2-Antioxidant Response Element Pathway: A Target for Regulating Energy Metabolism. J. Nutr. Biochem. 2012, 23, 1201–1206. [Google Scholar] [CrossRef]
- Ghosh, N.; Ghosh, R.; Mandal, S.C. Antioxidant Protection: A Promising Therapeutic Intervention in Neurodegenerative Disease. Free Radic. Res. 2011, 45, 888–905. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, X.; Hou, C.; Yang, L.; Li, H.; Guo, J.; Huo, C.; Wang, M.; Miao, Y.; Liu, J.; et al. Oleuropein Improves Mitochondrial Function to Attenuate Oxidative Stress by Activating the Nrf2 Pathway in the Hypothalamic Paraventricular Nucleus of Spontaneously Hypertensive Rats. Neuropharmacology 2017, 113, 556–566. [Google Scholar] [CrossRef]
- Parzonko, A.; Czerwinska, M.E.; Kiss, A.K.; Naruszewicz, M. Oleuropein and Oleacein May Restore Biological Functions of Endothelial Progenitor Cells Impaired by Angiotensin II via Activation of Nrf2/Heme Oxygenase-1 Pathway. Phytomedicine 2013, 20, 1088–1094. [Google Scholar] [CrossRef]
- Chang, T.; Untereiner, A.; Liu, J.; Wu, L. Interaction of Methylglyoxal and Hydrogen Sulfide in Rat Vascular Smooth Muscle Cells. Antioxid. Redox Signal. 2010, 12, 1093–1100. [Google Scholar] [CrossRef]
- Wang, H.; Meng, Q.H.; Chang, T.; Wu, L. Fructose-Induced Peroxynitrite Production Is Mediated by Methylglyoxal in Vascular Smooth Muscle Cells. Life Sci. 2006, 79, 2448–2454. [Google Scholar] [CrossRef] [PubMed]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 Cell Defence Pathway: Keap1-Dependent and -Independent Mechanisms of Regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherif, I.O. The Effect of Natural Antioxidants in Cyclophosphamide-Induced Hepatotoxicity: Role of Nrf2/HO-1 Pathway. Int. Immunopharmacol. 2018, 61, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.Y.; Chang, S.S.; Lin, I.H.; Chen, H.I. Suppression of Antioxidant Nrf-2 and Downstream Pathway in H9c2 Cells by Advanced Glycation End Products (Ages) via ERK Phosphorylation. Biochimie 2015, 118, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Castejón, M.L.; Montoya, T.; Alarcón-De-La-Lastra, C.; González-Benjumea, A.; Vázquez-Román, M.V.; Sánchez-Hidalgo, M. Dietary Oleuropein and Its Acyl Derivative Ameliorate Inflammatory Response in Peritoneal Macrophages from Pristane-Induced SLE Mice via Canonical and Noncanonical NLRP3 Inflammasomes Pathway. Food Funct. 2020, 11, 6622–6631. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delle Monache, S.; Pulcini, F.; Frosini, R.; Mattei, V.; Talesa, V.N.; Antognelli, C. Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants 2021, 10, 716. https://doi.org/10.3390/antiox10050716
Delle Monache S, Pulcini F, Frosini R, Mattei V, Talesa VN, Antognelli C. Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants. 2021; 10(5):716. https://doi.org/10.3390/antiox10050716
Chicago/Turabian StyleDelle Monache, Simona, Fanny Pulcini, Roberta Frosini, Vincenzo Mattei, Vincenzo Nicola Talesa, and Cinzia Antognelli. 2021. "Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway" Antioxidants 10, no. 5: 716. https://doi.org/10.3390/antiox10050716
APA StyleDelle Monache, S., Pulcini, F., Frosini, R., Mattei, V., Talesa, V. N., & Antognelli, C. (2021). Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants, 10(5), 716. https://doi.org/10.3390/antiox10050716