
Curcugreen Treatment Prevented Splenomegaly and Other Peripheral Organ Abnormalities in 3xTg and 5xFAD Mouse Models of Alzheimer’s Disease




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Curcugreen Solubility and Treatment
2.4. Tissue Collection
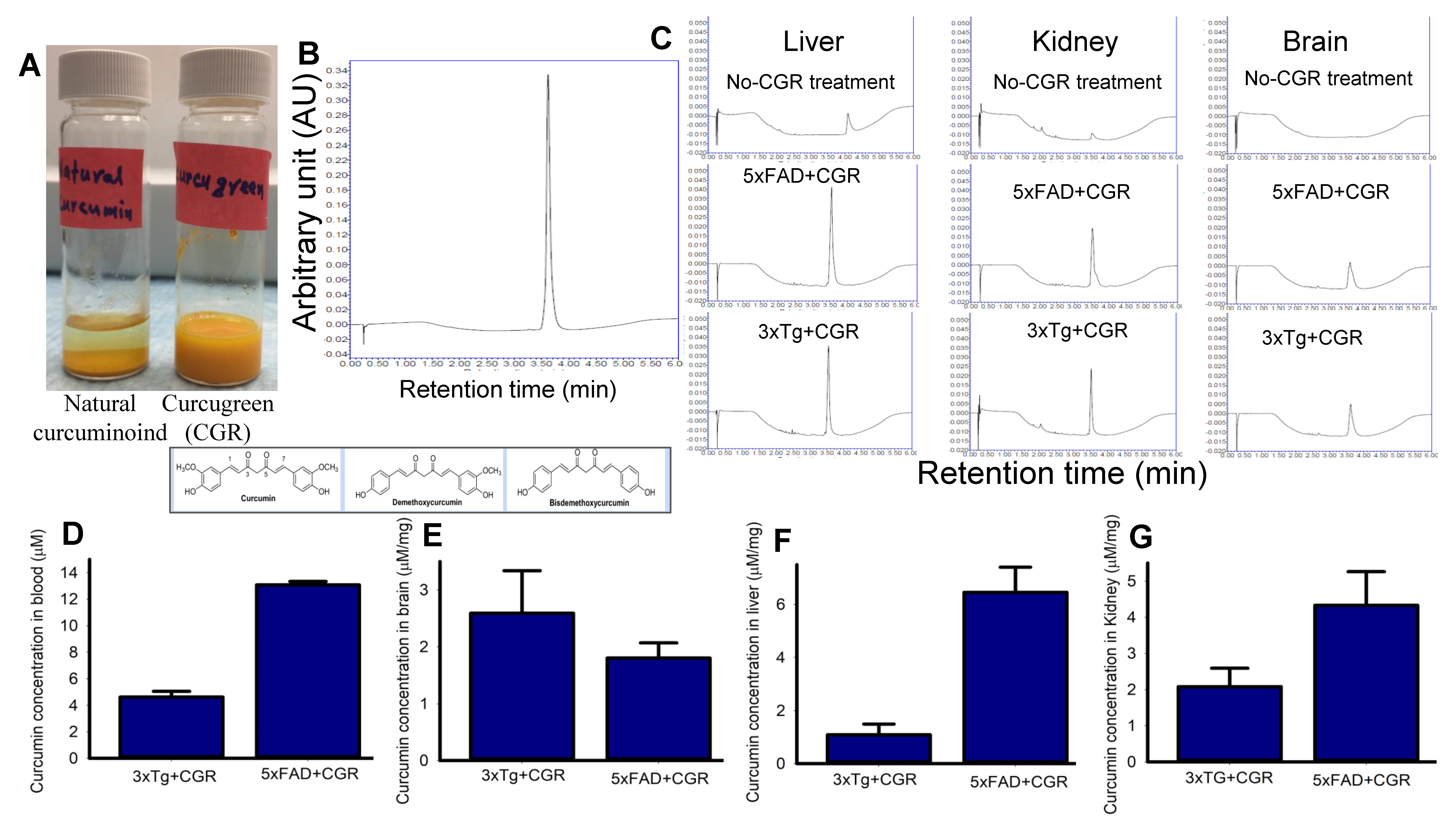
2.5. Curcumin Measurement by Ultra Performance Liquid Chromatography (UPLC)
2.6. Tissue Processing for Histology
2.6.1. Hematoxylin and Eosin (H&E) Stain
2.6.2. Terminal Deoxyribonucleic Acid Nick End Labeling (TUNEL)
2.6.3. Labeling of Amyloid Deposition with Congo Red (CR) Staining
2.6.4. DAPI and Propidium Iodide Staining
2.6.5. Immunohistochemistry of Phosphorylated Tau
2.6.6. Immunoperoxidase Staining for Cleaved Caspase-3
2.6.7. Western Blot
2.6.8. Statistical Analysis
3. Results
3.1. Curucmin Levels in Different Tissues in 3xTg and 5xFAD Mice after Treatment with CGR
3.2. Splenomegaly in 3xTg Mice Was Prevented by CGR Treatment
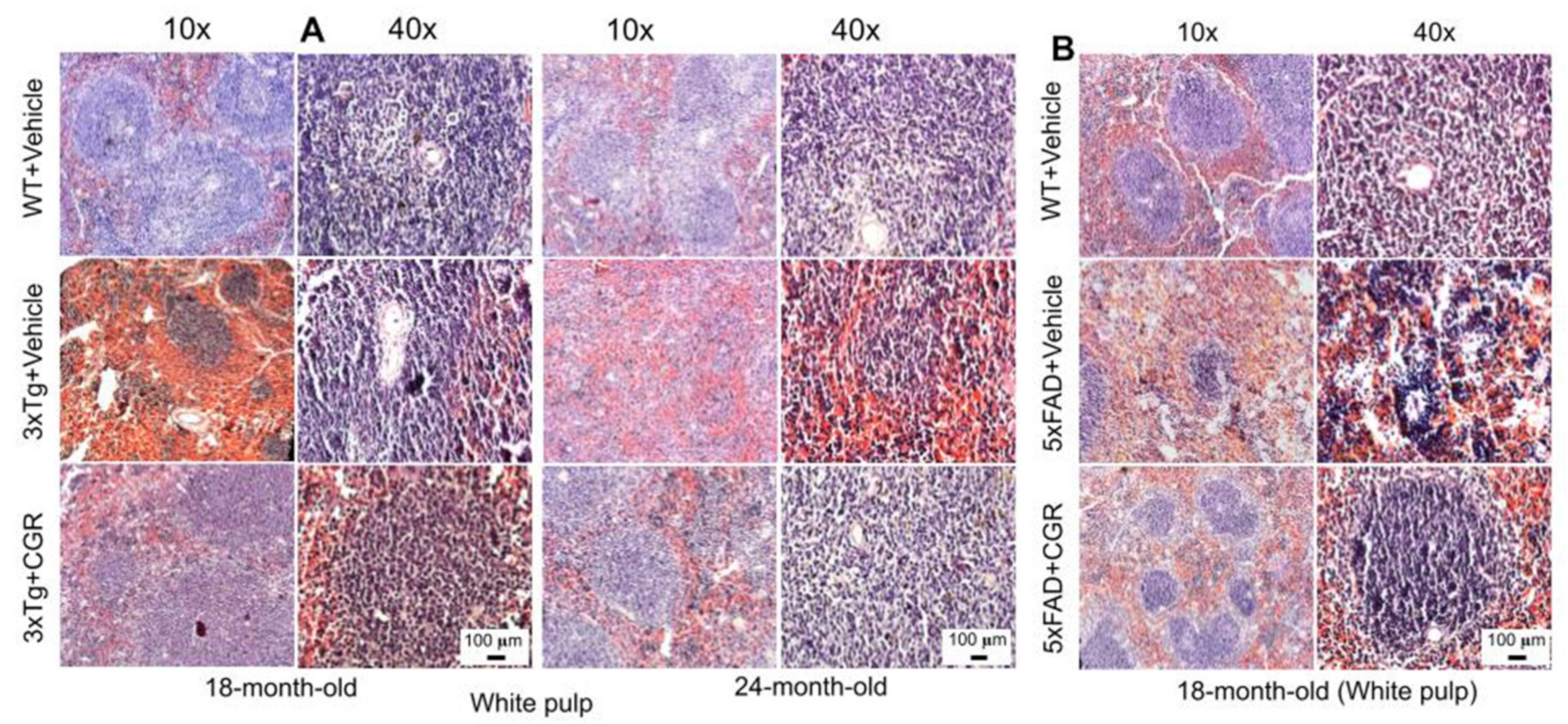
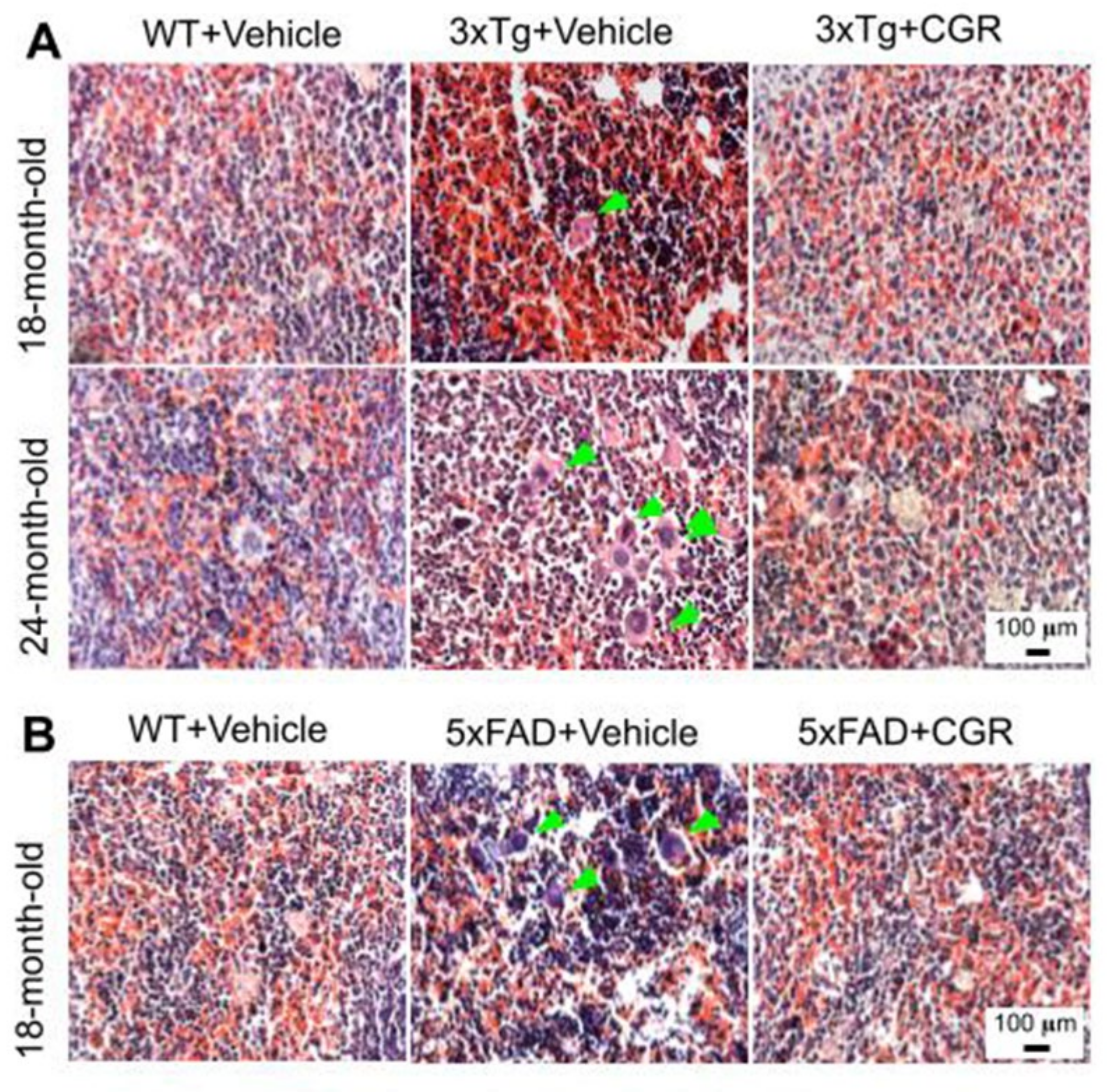
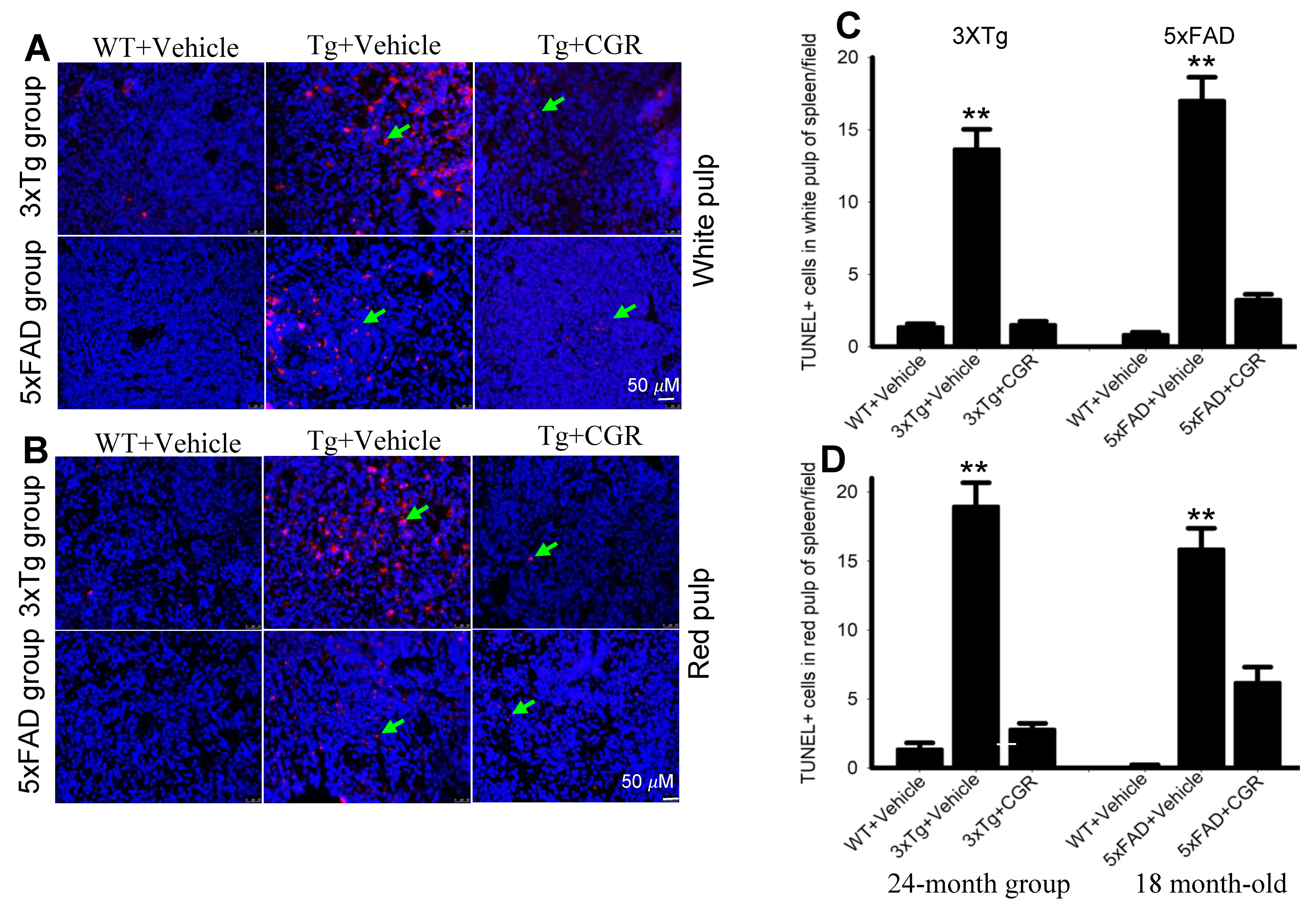
3.3. Morphological Damage of Splenic White- and Red-Pulp Were Prevented by CGR Treatment in Both 3xTg and 5xFAD Mice
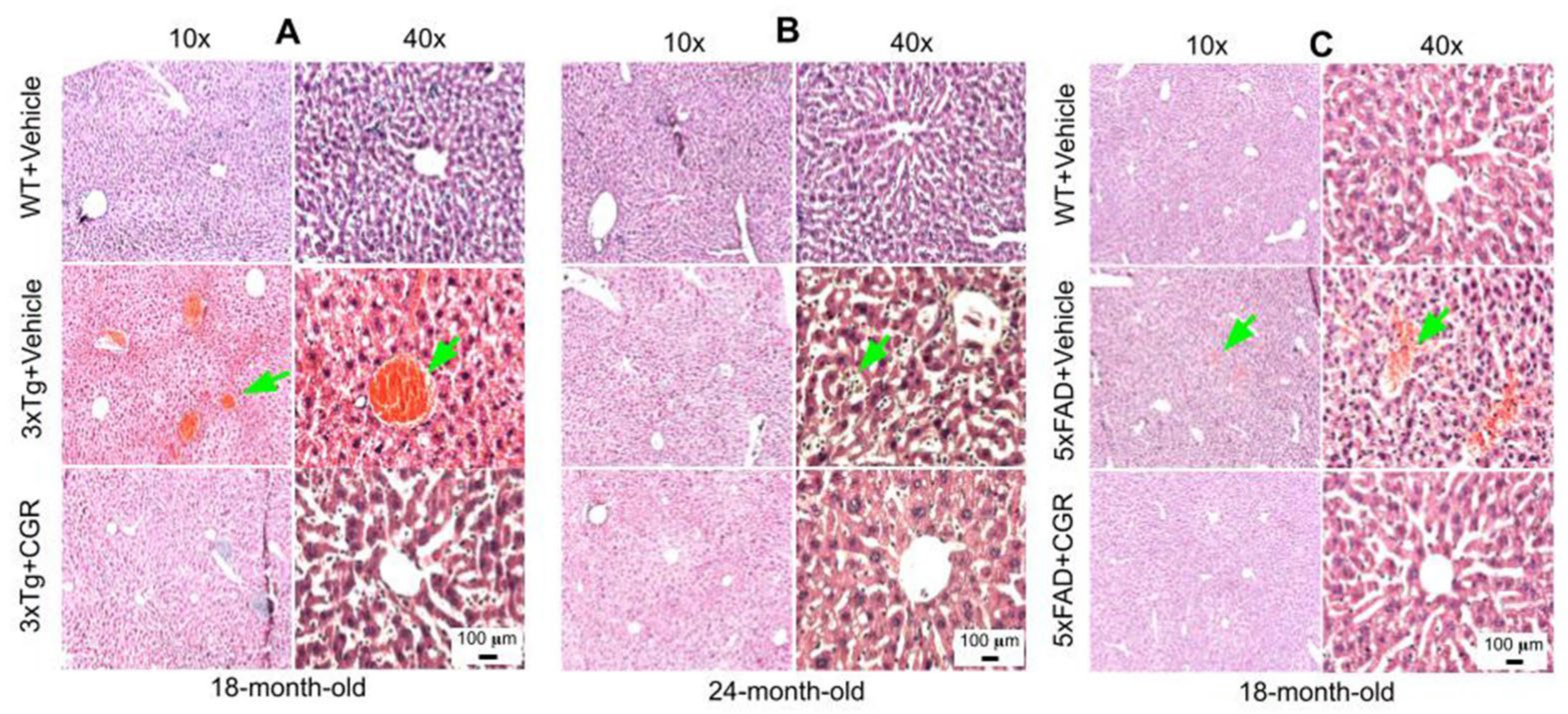
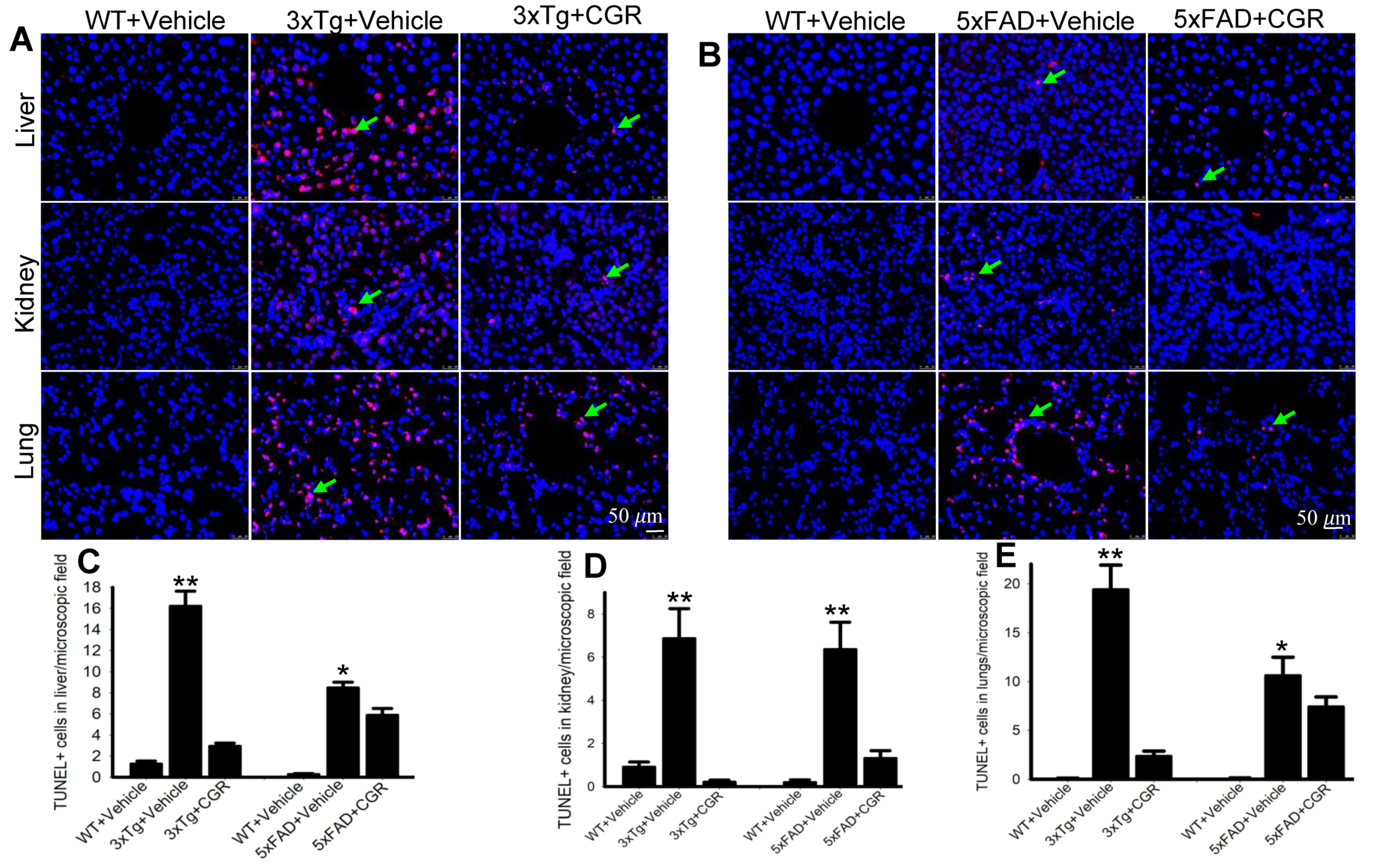
3.4. Morphology of the Liver Was Restored by CGR Treatment in Both 3xTg and 5xFAD Mice
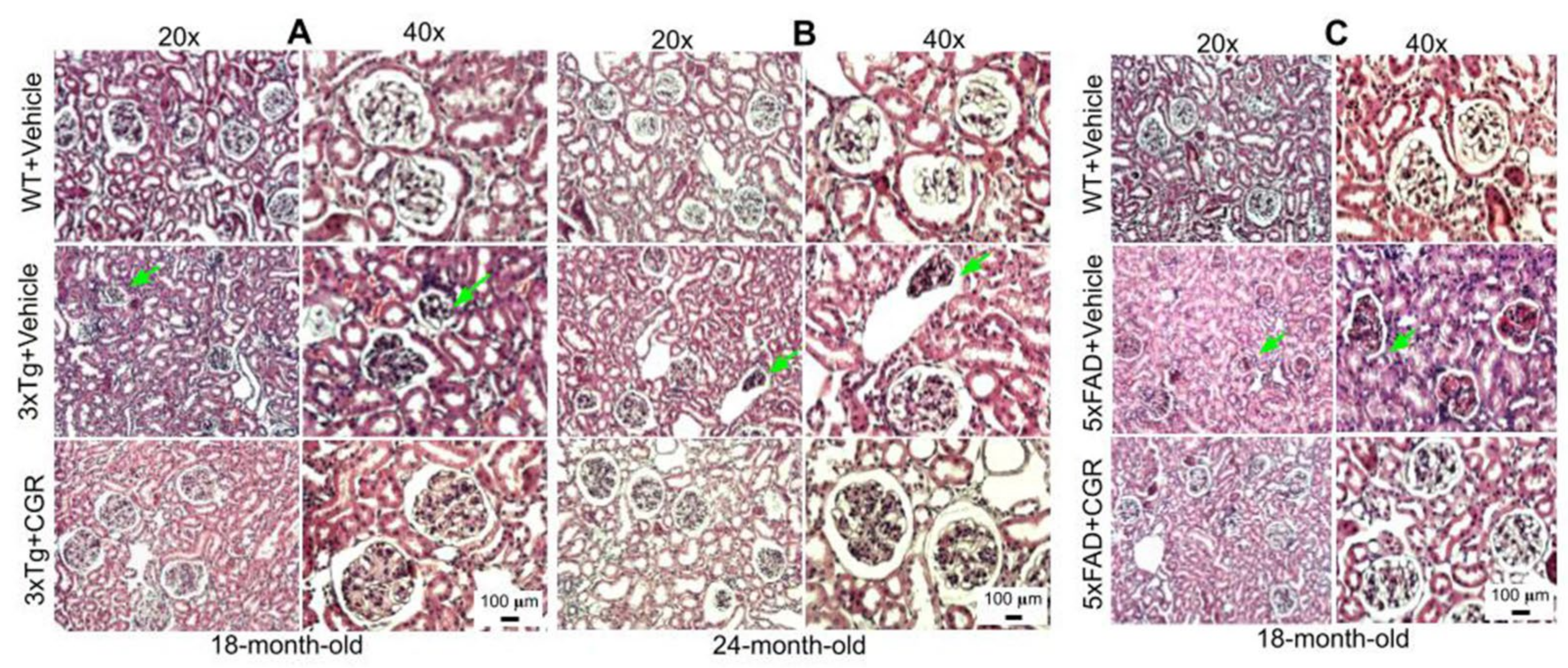
3.5. Kidney Morphology Was Restored by CGR Treatment in 3xTg and 5xFAD Mice
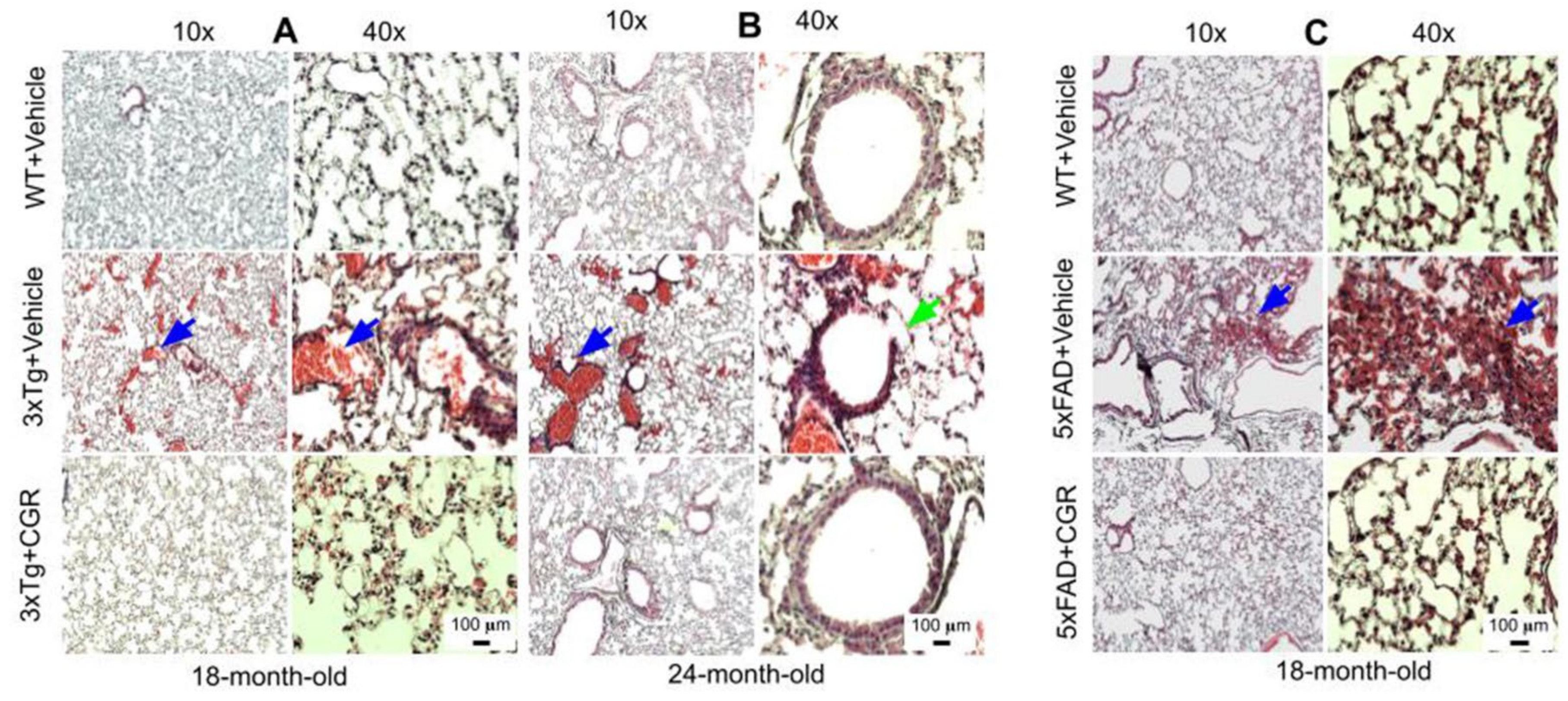
3.6. Lung Morphology Was Restored by CGR Treatment in Both the 3xTg and 5xFAD Mice
3.7. Apoptotic Death (DNA Fragmentation) Was Prevented by CGR Treatment in the Spleen, Liver, Kidney and Lung Tissue of 3xTg and 5xFAD Mice
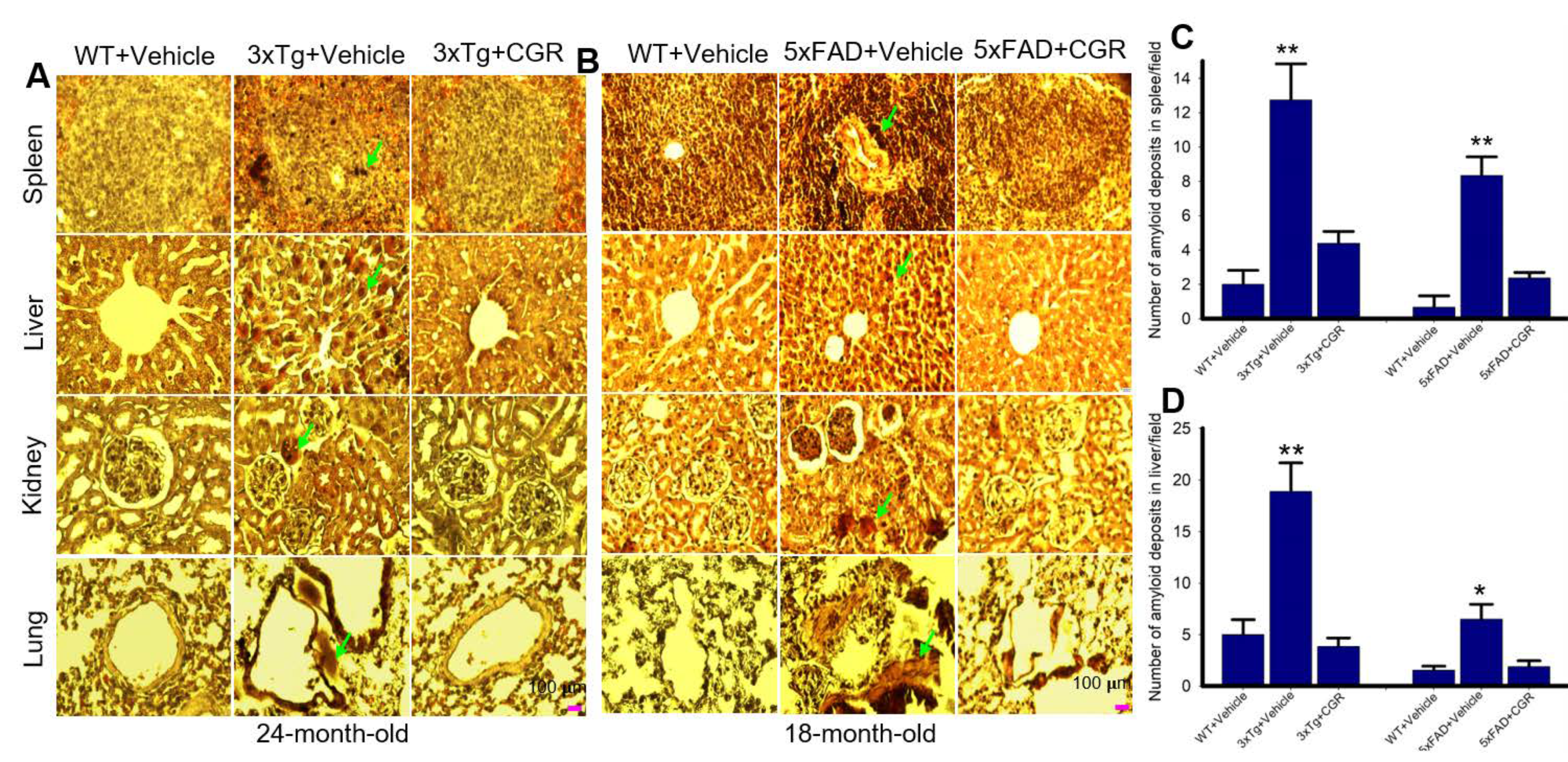
3.8. Amyloid Deposits in the Peripheral Organs in the 3xTg and 5xFAD Mice Were Reduced by CGR Treatment
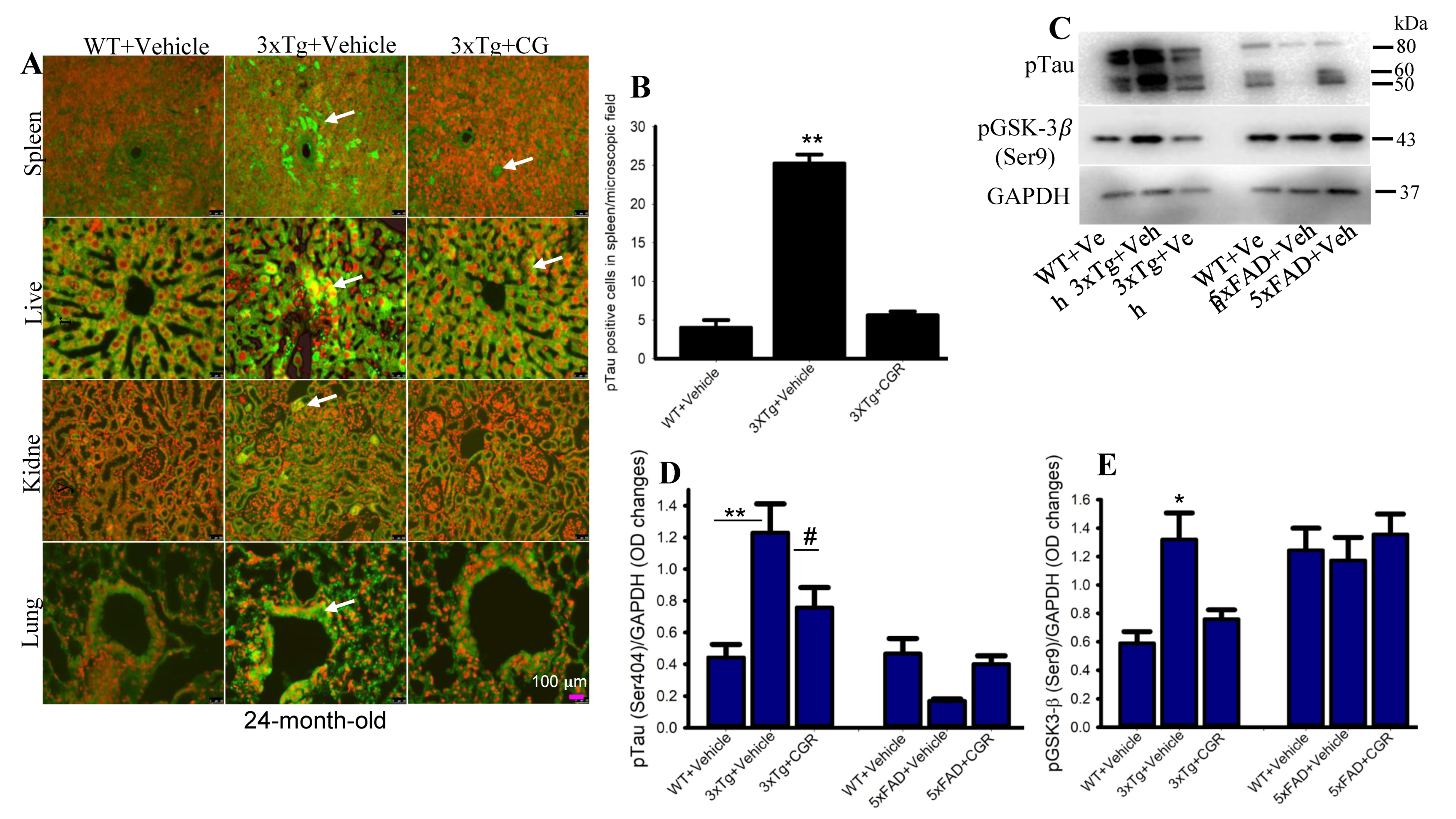
3.9. Phosphorylated Tau Immunoreactivity Was Reduced by CGR Treatment in 3xTg Mice
3.10. Cleaved Caspase-3 Immunoreactivity in 3xTg and 5xFAD Mice after Treatment with CGR
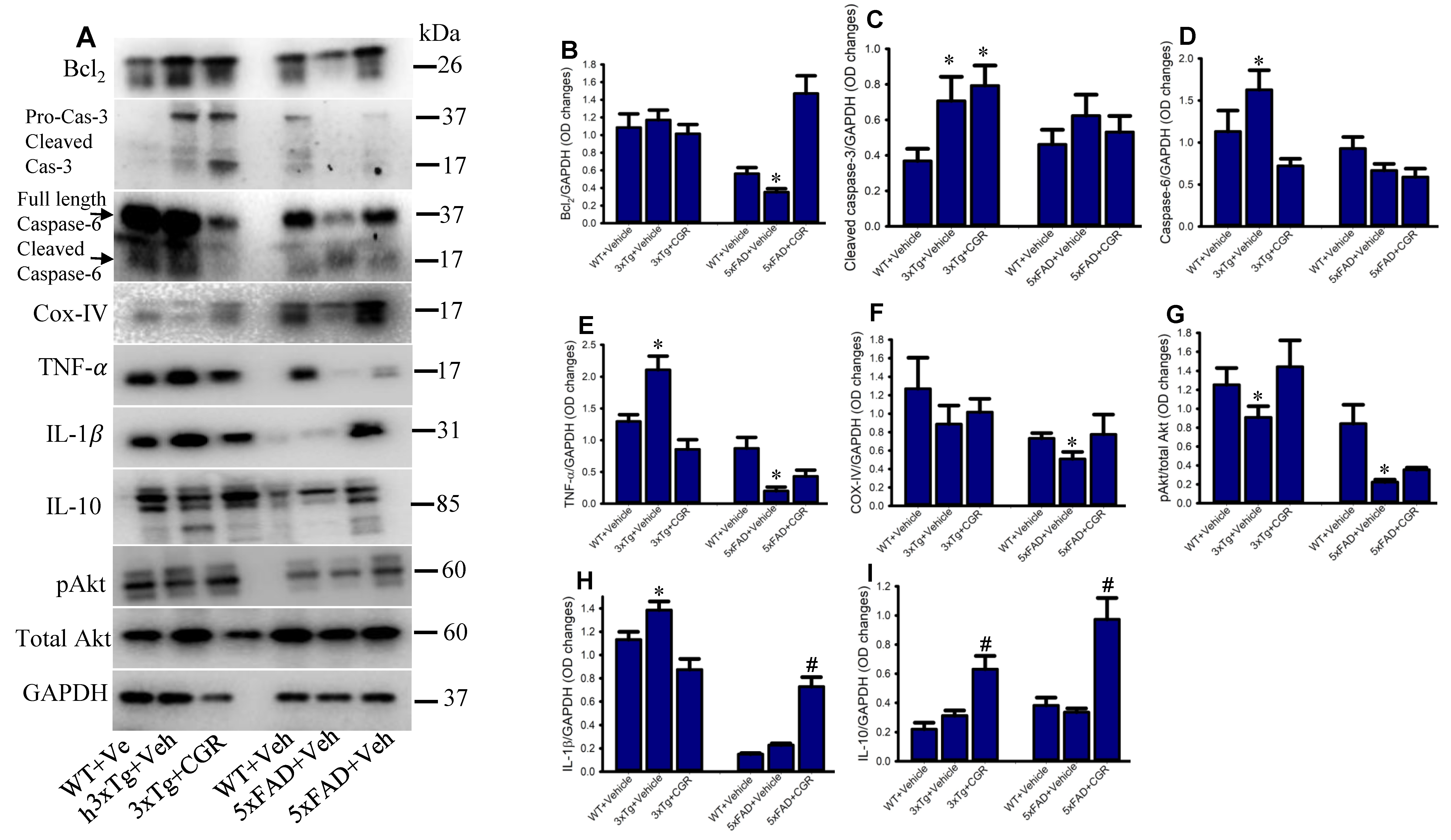
3.11. Cell-Death and Cell-Survival, Inflammatory and Anti-Inflammatory Markers Were Normalized by CGR Treatment in the Spleen of 3xTg and 5xFAD Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cai, H.; Cong, W.-N.; Ji, S.; Rothman, S.; Maudsley, S.; Martin, B. Metabolic Dysfunction in Alzheimer’s Disease and Related Neurodegenerative Disorders. Curr. Alzheimer Res. 2012, 9, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T.; Rogers, J. Inflammation in Alzheimer Disease--A Brief Review of the Basic Science and Clinical Literature. Cold Spring Harb. Perspect. Med. 2011, 2, a006346. [Google Scholar] [CrossRef] [PubMed]
- Trollor, J.N.; Smith, E.; Baune, B.T.; Kochan, N.A.; Campbell, L.; Samaras, K.; Crawford, J.; Brodaty, H.; Sachdev, P. Systemic Inflammation Is Associated with MCI and Its Subtypes: The Sydney Memory and Aging Study. Dement. Geriatr. Cogn. Disord. 2010, 30, 569–578. [Google Scholar] [CrossRef]
- Tarkowski, E.; Andreasen, N.; Tarkowski, A.; Blennow, K. Intrathecal inflammation precedes development of Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1200–1205. [Google Scholar] [CrossRef]
- Craft, S. The Role of Metabolic Disorders in Alzheimer’s Disease and Vascular Dementia: Two Roads Converged? Arch. Neurol. 2009, 66, 300–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildreth, K.L.; Van-Pelt, R.E.; Schwartz, R.S. Obesity, Insulin Resistance, and Alzheimer’s Disease. Obesity 2012, 20, 1549–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cynthia, M. Carlsson. Type 2 Diabetes Mellitus, Dyslipidemia, and Alzheimer’s Disease. J. Alzheimers Dis. 2010, 20, 711–722. [Google Scholar]
- Nho, K.; Kueider-Paisley, A.; Ahmad, S.; Mahmoudian, D.S.; Arnold, M.; Risacher, S.L.; Louie, G.; Blach, C.; Baillie, R.; Han, X.; et al. Association of Altered Liver Enzymes with Alzheimer Disease Diagnosis, Cognition, Neuroimaging Measures, and Cerebrospinal Fluid Biomarkers. JAMA Netw. Open 2019, 2, e197978. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Liao, Y.; Chen, X.; Mai, N.; Ouyang, C.; Chen, B.; Zhang, M.; Peng, Q.; Liang, W.; Zhang, W.; et al. Abnormal Serum Bilirubin/Albumin Concentrations in Dementia Patients with Aβ Deposition and the Benefit of Intravenous Albumin Infusion for Alzheimer’s Disease Treatment. Front. Neurosci. 2020, 14, 859. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ding, D.; Zhao, Q.; Wu, W.; Xiao, Z.; Liang, X.; Luo, J.; Chen, J. Kidney function and dementia risk in community-dwelling older adults: The Shanghai Aging Study. Alzheimer’s Res. Ther. 2021, 13, 1–9. [Google Scholar] [CrossRef]
- Dodd, J.W. Lung disease as a determinant of cognitive decline and dementia. Alzheimer’s Res. Ther. 2015, 7, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Lutsey, P.L.; Chen, N.; Mirabelli, M.C.; Lakshminarayan, K.; Knopman, D.S.; Vossel, K.A.; Gottesman, R.F.; Mosley, T.H.; Alonso, A. Impaired Lung Function, Lung Disease, and Risk of Incident Dementia. Am. J. Respir. Crit. Care Med. 2019, 199, 1385–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, R.; LaFerla, F.M. Transgenic Mouse Models of Alzheimer Disease: Developing a Better Model as a Tool for Therapeutic Interventions. Curr. Pharm. Des. 2012, 18, 1131–1147. [Google Scholar] [CrossRef] [Green Version]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-transgenic model of Alzheimer’s disease with plaques and tangles: Intracellular Abeta and synaptic dysfunction. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Billings, L.M.; Oddo, S.; Green, K.N.; McGaugh, J.L.; LaFerla, F.M. Intraneuronal Abeta causes the onset of early Alzheimer’s disease-related cognitive deficits in transgenic mice. Neuron 2005, 45, 675–688. [Google Scholar] [CrossRef] [Green Version]
- Stover, K.R.; Campbell, M.A.; Van Winssen, C.M.; Brown, R.E. Early detection of cognitive deficits in the 3xTg-AD mouse model of Alzheimer’s disease. Behav. Brain Res. 2015, 289, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Maya-Vetencourta, J.F.; Caruccia, N.M.; Capsonia, S.; Cattaneo, A.A. Amyloid Plaque-Independent Deficit of Early Postnatal Visual Cortical Plasticity in the 5xFAD Transgenic Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 42, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Kimura, R.; Ohno, M. Impairments in remote memory stabilization precede hippocampal synaptic and cognitive failures in 5XFAD Alzheimer mouse model. Neurobiol. Dis. 2009, 33, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Marchese, M.; Cowan, D.; Head, E.; Ma, D.; Karimi, K.; Ashthorpe, V.; Kapadia, M.; Zhao, H.; Davis, P.; Sakicd, B. Autoimmune Manifestations in the 3xTg-AD Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 39, 191–210. [Google Scholar] [CrossRef] [Green Version]
- Montacute, R.; Foley, K.; Forman, R.; Else, K.J.; Cruickshank, S.M.; Allan, S.M. Enhanced susceptibility of triple transgenic Alzheimer’s disease (3xTg-AD) mice to acute infection. J. Neuroinflamm. 2017, 14, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.-L.; Zuo, X.; Yang, F.; Ubeda, O.J.; Gant, D.J.; Alaverdyan, M.; Teng, E.; Hu, S.; Chen, P.-P.; Maiti, P.; et al. Curcumin Suppresses Soluble Tau Dimers and Corrects Molecular Chaperone, Synaptic, and Behavioral Deficits in Aged Human Tau Transgenic Mice. J. Biol. Chem. 2013, 288, 4056–4065. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Maiti, P.; Ma, Q.; Zuo, X.; Jones, M.R.; Cole, G.M.; Frautschy, S.A. Clinical development of curcumin in neurodegenerative disease. Expert Rev. Neurother. 2015, 15, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Hall, T.C.; Paladugu, L.; Kolli, N.; Learman, C.; Rossignol, J.; Dunbar, G.L. A comparative study of dietary curcumin, nanocurcumin, and other classical amyloid-binding dyes for labeling and imaging of amyloid plaques in brain tissue of 5×-familial Alzheimer’s disease mice. Histochem. Cell Biol. 2016, 146, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Dunbar, G.L. Comparative Neuroprotective Effects of Dietary Curcumin and Solid Lipid Curcumin Particles in Cultured Mouse Neuroblastoma Cells after Exposure to Aβ42. Int. J. Alzheimer’s Dis. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Paladugu, L.; Dunbar, G.L. Solid lipid curcumin particles provide greater anti-amyloid, anti-inflammatory and neuroprotective effects than curcumin in the 5xFAD mouse model of Alzheimer’s disease. BMC Neurosci. 2018, 19, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Dunbar, G.L. Use of Curcumin, a Natural Polyphenol for Targeting Molecular Pathways in Treating Age-Related Neurodegenerative Diseases. Int. J. Mol. Sci. 2018, 19, 1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, P.; Plemmons, A.; Bowers, Z.; Weaver, C.; Dunbar, G. Labeling and Imaging of Amyloid Plaques in Brain Tissue Using the Natural Polyphenol Curcumin. J. Vis. Exp. 2019, e60377. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Bowers, Z.; Bourcier-Schultz, A.; Morse, J.; Dunbar, G.L. Preservation of dendritic spine morphology and postsynaptic signaling markers after treatment with solid lipid curcumin particles in the 5xFAD mouse model of Alzheimer’s amyloidosis. Alzheimer’s Res. Ther. 2021, 13, 1–22. [Google Scholar] [CrossRef]
- Gharaibeh, A.; Maiti, P.; Culver, R.; Heileman, S.; Srinageshwar, B.; Story, D.; Spelde, K.; Paladugu, L.; Munro, N.; Muhn, N.; et al. Solid Lipid Curcumin Particles Protect Medium Spiny Neuronal Morphology, and Reduce Learning and Memory Deficits in the YAC128 Mouse Model of Huntington’s Disease. Int. J. Mol. Sci. 2020, 21, 9542. [Google Scholar] [CrossRef]
- Peng, Y.; Gao, P.; Shi, L.; Chen, L.; Liu, J.; Long, J. Central and Peripheral Metabolic Defects Contribute to the Pathogenesis of Alzheimer’s Disease: Targeting Mitochondria for Diagnosis and Prevention. Antioxid. Redox Signal. 2020, 32, 1188–1236. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.L. Clinical Research Overview BCM-95®/Curcugreen turmeric rhizome/curucmin extract preparation. In Proprietary Botanical Ingredient Scientific and Clinical Monograph for BCM-95®/CurcugreenTM Turmeric Rhizome/Curcumin Extract Preparation; American Botanical Council: Austin, TX, USA, 2019; Available online: https://www.herbalgram.org/media/13291/bcm95_mono-03-2019.pdf (accessed on 2 April 2021).
- Maiti, P.; Gharaibeh, A.; Kolli, N.; Dunbar, G.L. Solid Lipid Curcumin Particles Induce More DNA Fragmentation and Cell Death in Cultured Human Glioblastoma Cells than Does Natural Curcumin. Oxidative Med. Cell. Longev. 2017, 2017, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, P.; Peruzzaro, S.; Kolli, N.; Andrews, M.; Al-Gharaibeh, A.; Rossignol, J.; Dunbar, G.L. Transplantation of mesenchymal stem cells overexpressing interleukin-10 induces autophagy response and promotes neuroprotection in a rat model of TBI. J. Cell. Mol. Med. 2019, 23, 5211–5224. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2011, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Su, I.-J.; Chang, H.-Y.; Wang, H.-C.; Tsai, K.-J. A Curcumin Analog Exhibits Multiple Biologic Effects on the Pathogenesis of Alzheimer’s Disease and Improves Behavior, Inflammation, and β-Amyloid Accumulation in a Mouse Model. Int. J. Mol. Sci. 2020, 21, 5459. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-H.; Kim, J.; Lee, M.J.; Kim, Y. Abnormalities of plasma cytokines and spleen in senile APP/PS1/Tau transgenic mouse model. Sci. Rep. 2015, 5, 15703. [Google Scholar] [CrossRef] [Green Version]
- Estrada, L.D.; Ahumada, P.; Cabrera, D.; Arab, J.P. Liver Dysfunction as a Novel Player in Alzheimer’s Progression: Looking Outside the Brain. Front. Aging Neurosci. 2019, 11, 174. [Google Scholar] [CrossRef] [Green Version]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manna, J.; Dunbar, G.L.; Maiti, P. Curcugreen Treatment Prevented Splenomegaly and Other Peripheral Organ Abnormalities in 3xTg and 5xFAD Mouse Models of Alzheimer’s Disease. Antioxidants 2021, 10, 899. https://doi.org/10.3390/antiox10060899
Manna J, Dunbar GL, Maiti P. Curcugreen Treatment Prevented Splenomegaly and Other Peripheral Organ Abnormalities in 3xTg and 5xFAD Mouse Models of Alzheimer’s Disease. Antioxidants. 2021; 10(6):899. https://doi.org/10.3390/antiox10060899
Chicago/Turabian StyleManna, Jayeeta, Gary L. Dunbar, and Panchanan Maiti. 2021. "Curcugreen Treatment Prevented Splenomegaly and Other Peripheral Organ Abnormalities in 3xTg and 5xFAD Mouse Models of Alzheimer’s Disease" Antioxidants 10, no. 6: 899. https://doi.org/10.3390/antiox10060899
APA StyleManna, J., Dunbar, G. L., & Maiti, P. (2021). Curcugreen Treatment Prevented Splenomegaly and Other Peripheral Organ Abnormalities in 3xTg and 5xFAD Mouse Models of Alzheimer’s Disease. Antioxidants, 10(6), 899. https://doi.org/10.3390/antiox10060899