
Phytochemical Study and In Vitro Screening Focusing on the Anti-Aging Features of Various Plants of the Greek Flora

 , , ,
, , ,  , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction
2.3. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
2.4. Tyrosinase Inhibition
2.5. Hyaluronidase Inhibition
2.6. Elastase Inhibitory Activity
2.7. High-Performance Thin-Layer Chromatography (HPTLC) Profiling
2.8. Ultra-Performance Liquid Chromatography High-Resolution Mass Spectrometry (UPLC-HRMS) Analysis
2.9. Cell Lines and Cell Culture Conditions
2.10. Assessment of Cytotoxicity
2.11. Intracellular ROS Assay
2.12. Induction of Premature Senescence by H2O2 and SA β-Gal Staining
2.13. Measurement of Cellular Tyrosinase Activity
2.14. Measurement of Proteasome Chymotrypsin-Like Activity
2.15. Measurement of Cathepsins B, L Activity
2.16. Sirtuin 1 Deacetylase Activity Assay
2.17. Immunofluorescence Antigen Staining
2.18. MMP Activity Assay
2.19. Assessment of Cell Protective Activity against UV-B Irradiation
2.20. Statistical Analysis
3. Results
3.1. Selection and Extraction of Plant Species
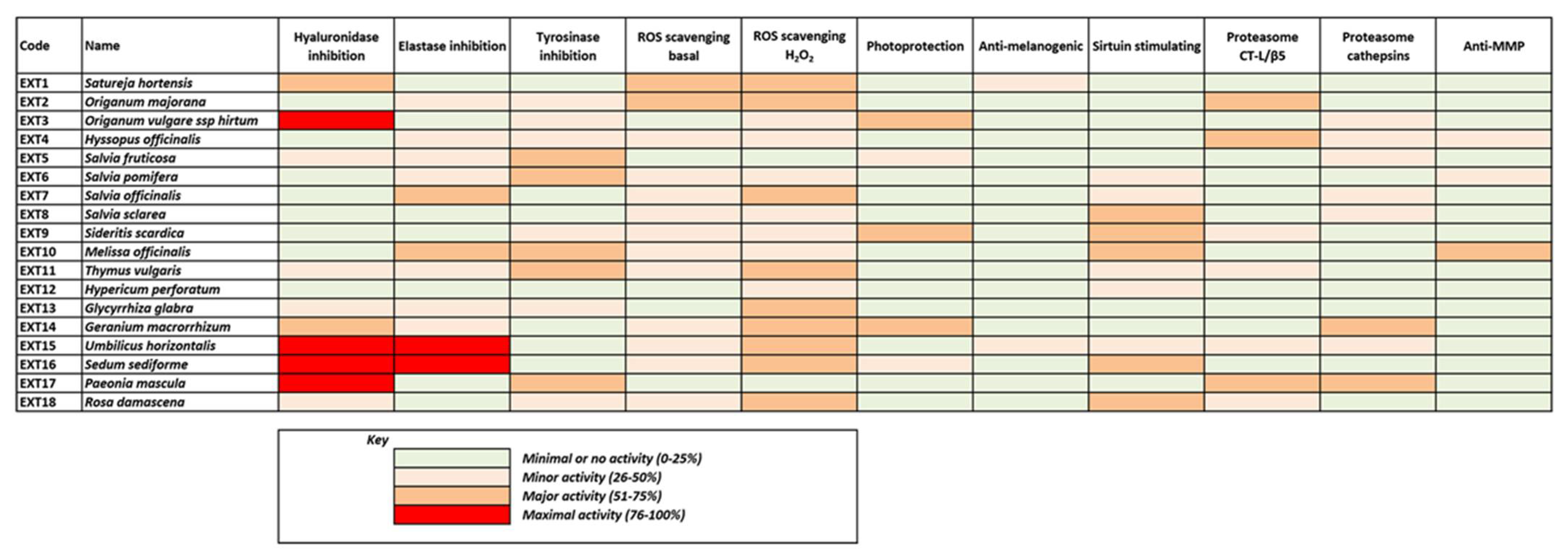
3.2. Antioxidant and Anti-Melanogenic Activities of Plant Extracts
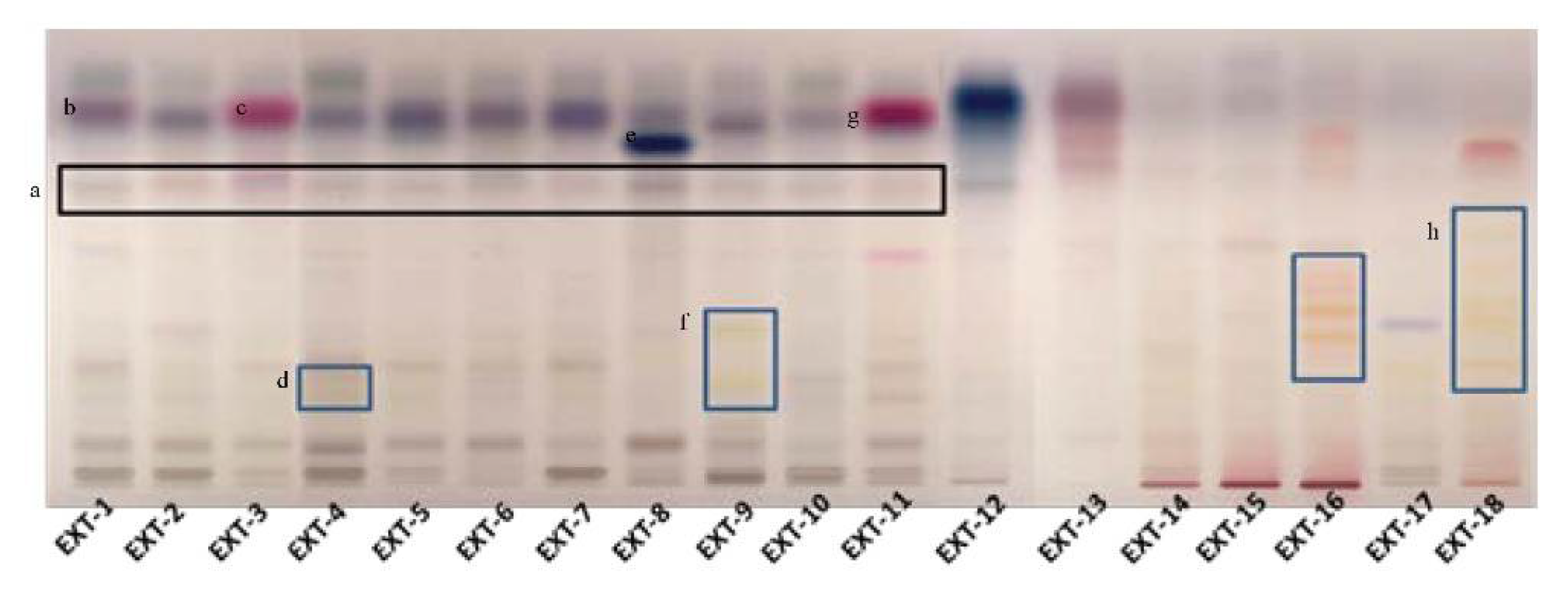
3.3. Phytochemical Profile of the Extracts
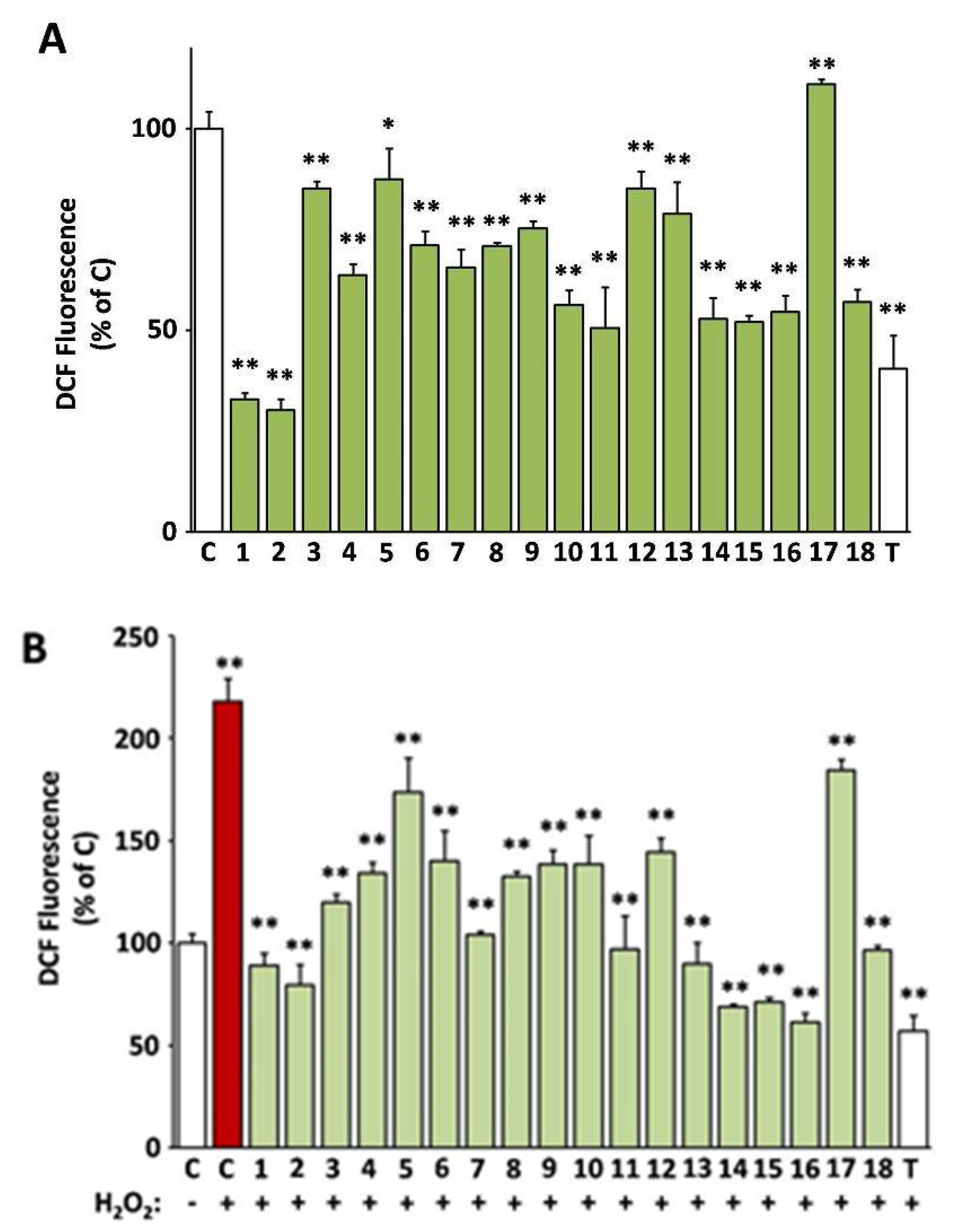
3.4. Intracellular Antioxidant Activity of the Extracts
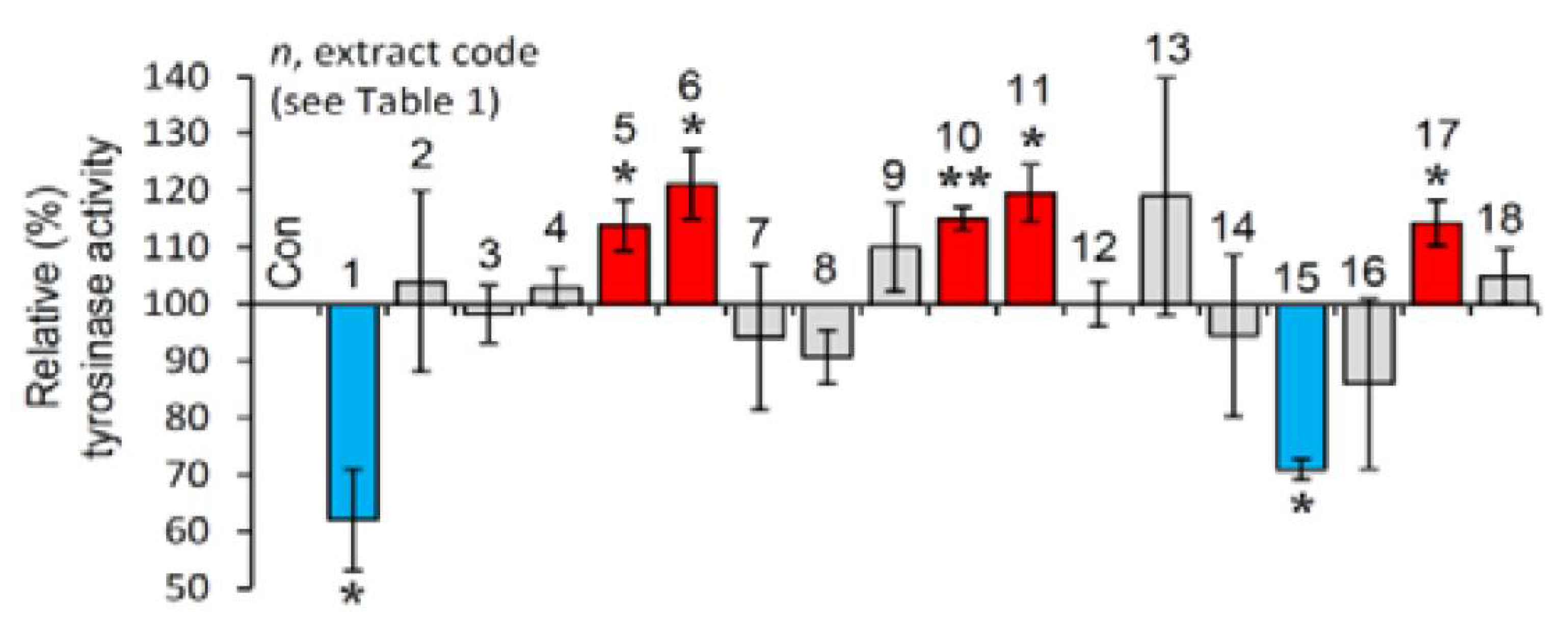
3.5. Cell-Based Evaluation of the Anti-Melanogenic Activity of the Extracts
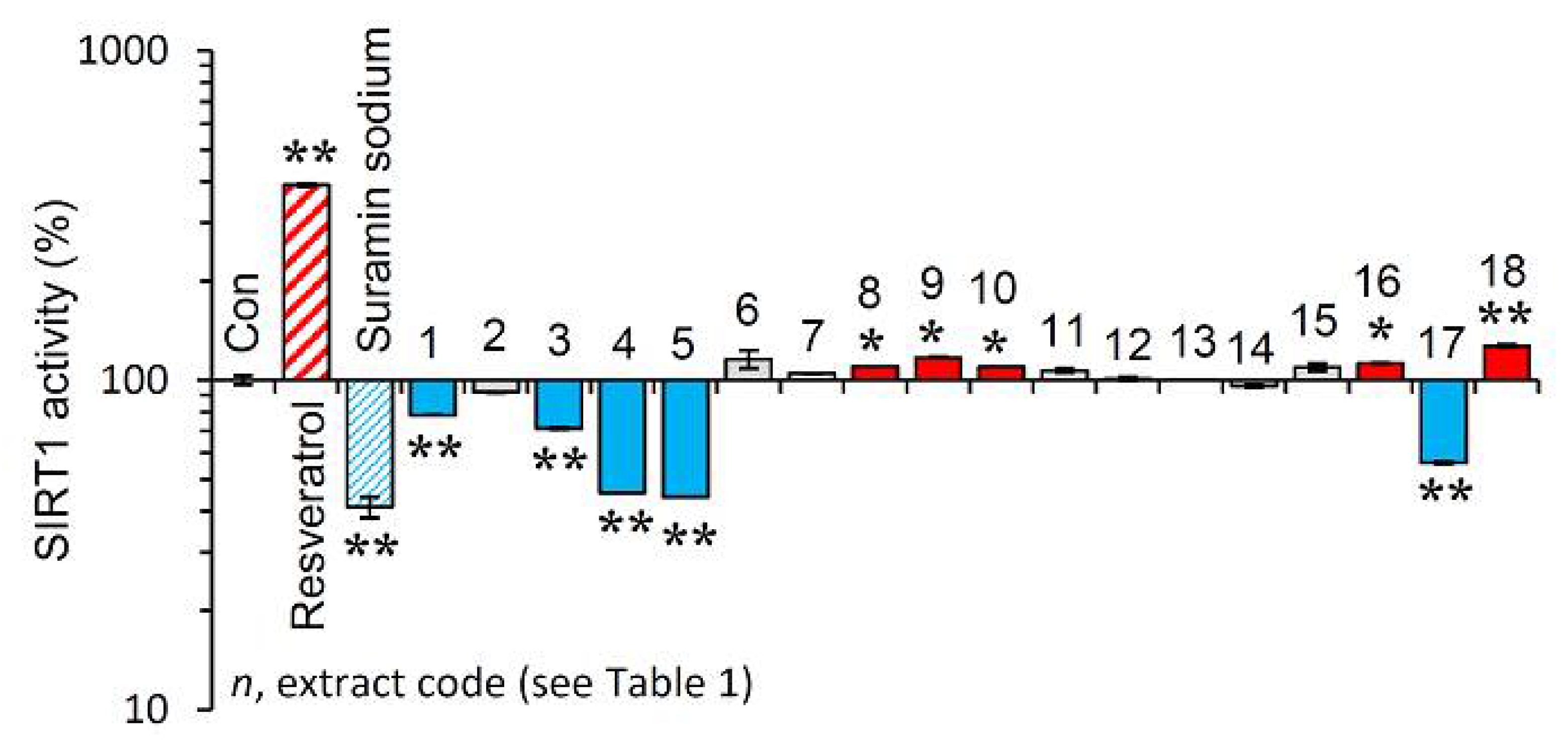
3.6. Effect of the Extracts on Sirtuin 1 Enzymatic Activity
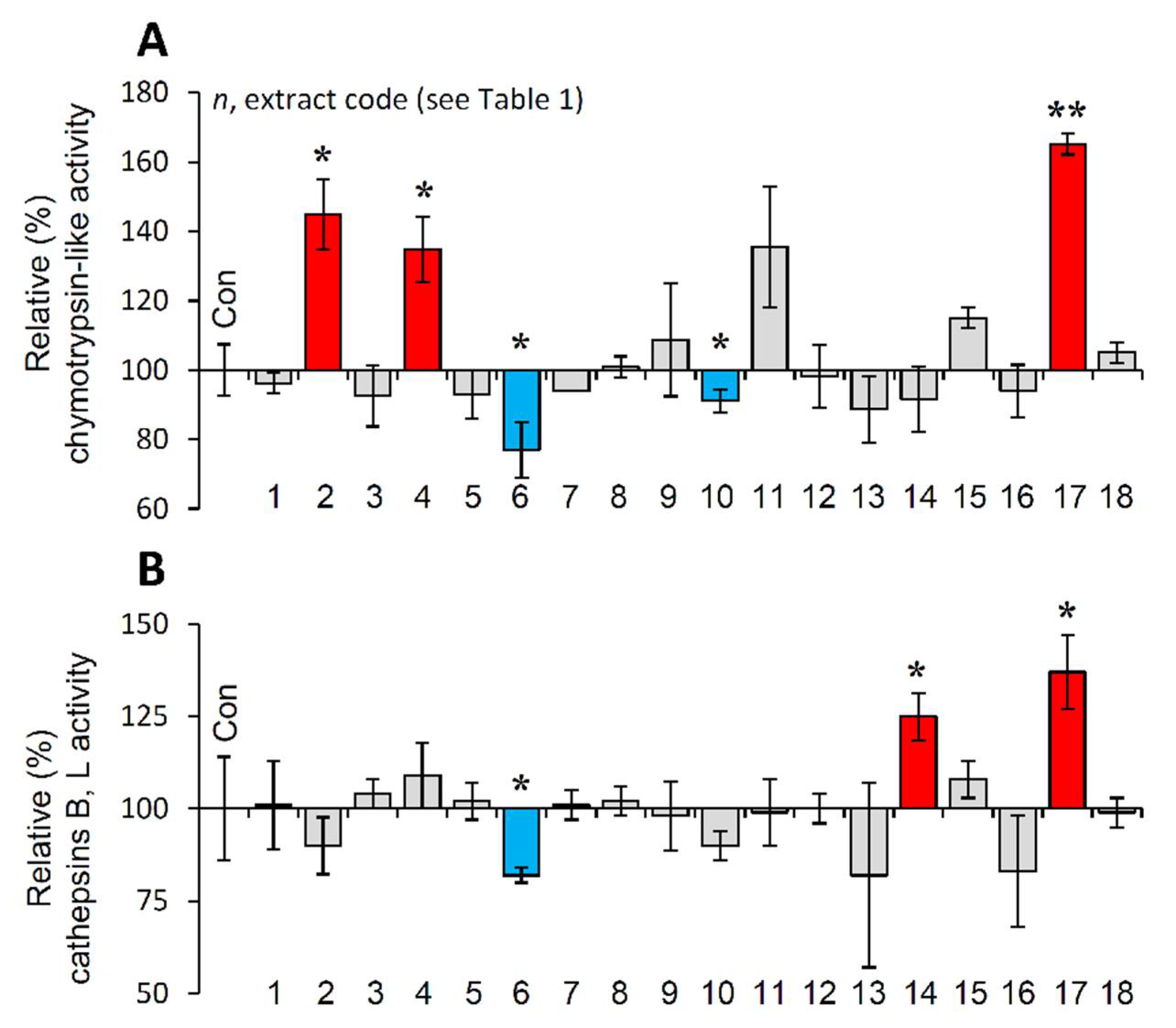
3.7. Effect of the Extracts on the Main Proteostatic Pathways
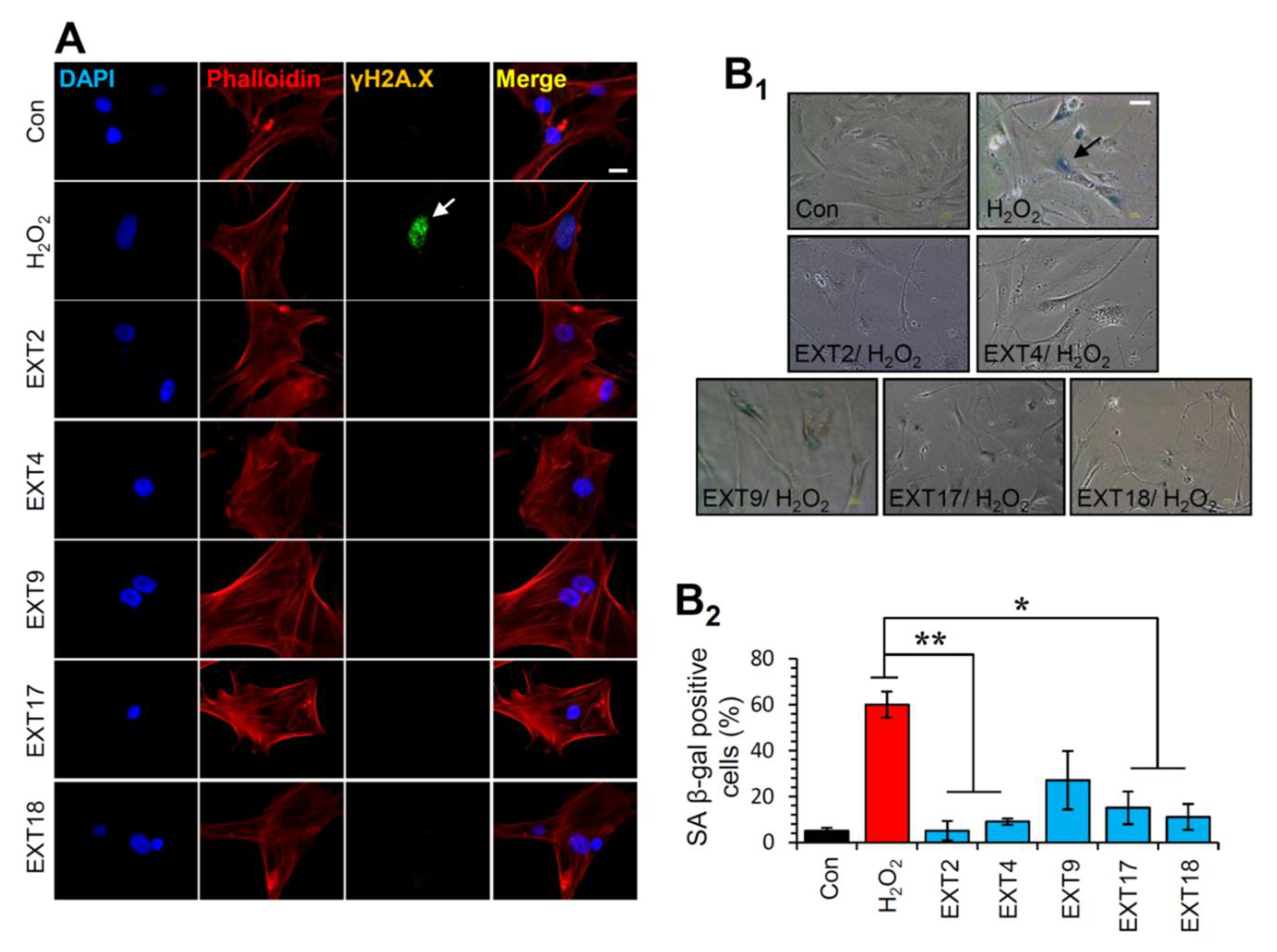
3.8. Protection of Cells against Oxidative Stress-Mediated Premature Senescence by the Extracts
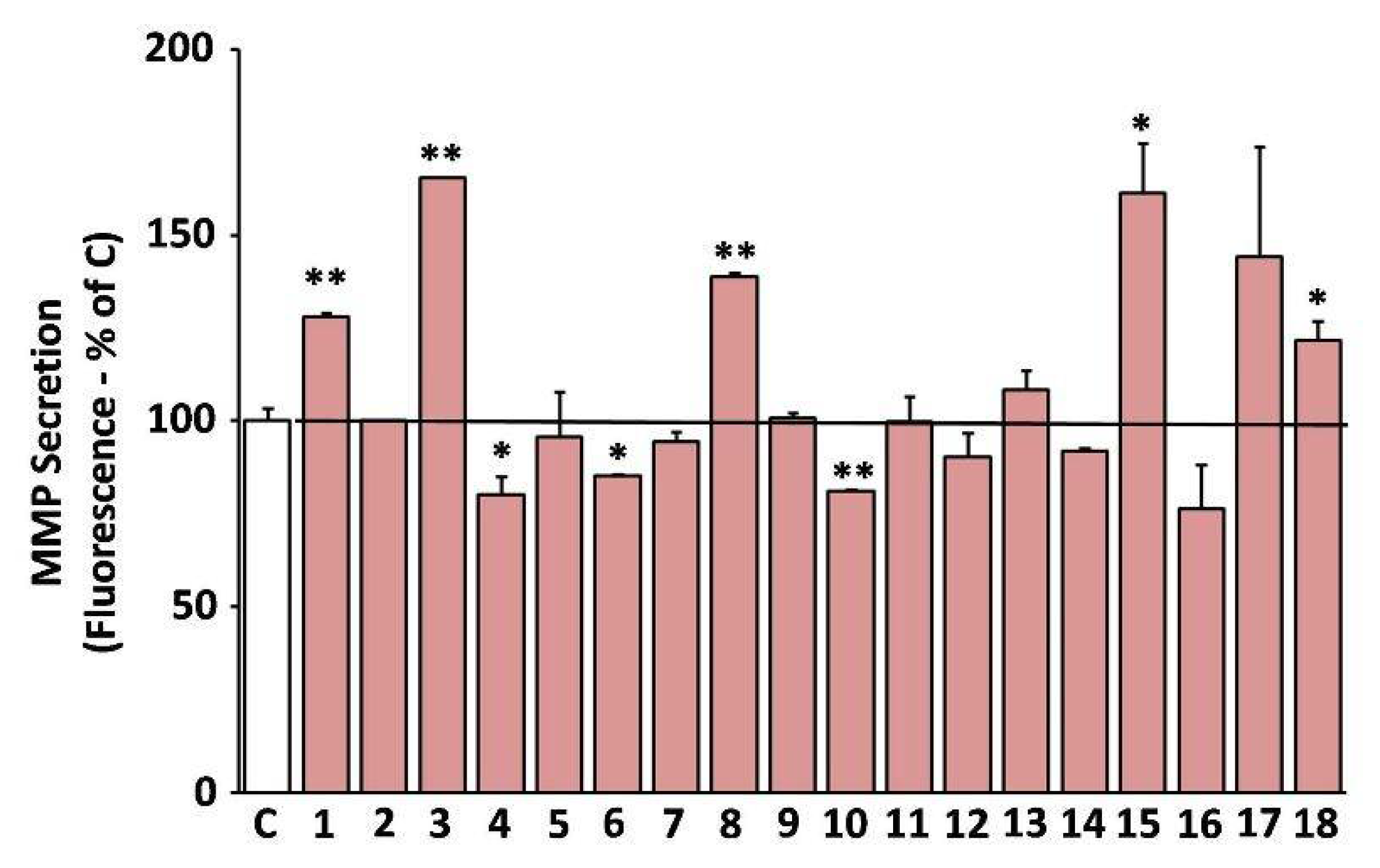
3.9. Suppression of the Secreted Matrix Metalloproteinases (MMP)-Activity by the Extracts
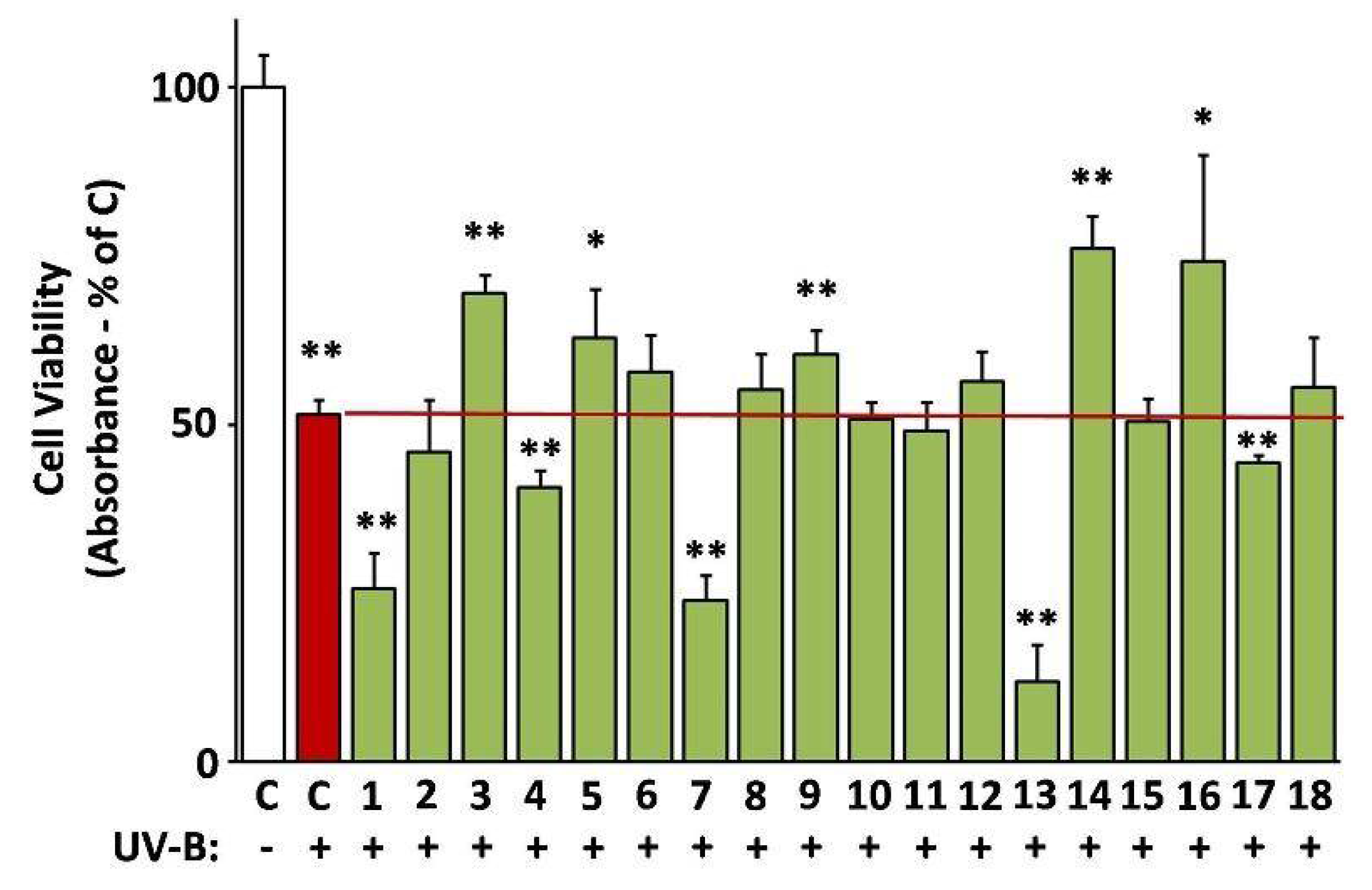
3.10. Protective Activity of the Extracts against UV-B Cytotoxicity
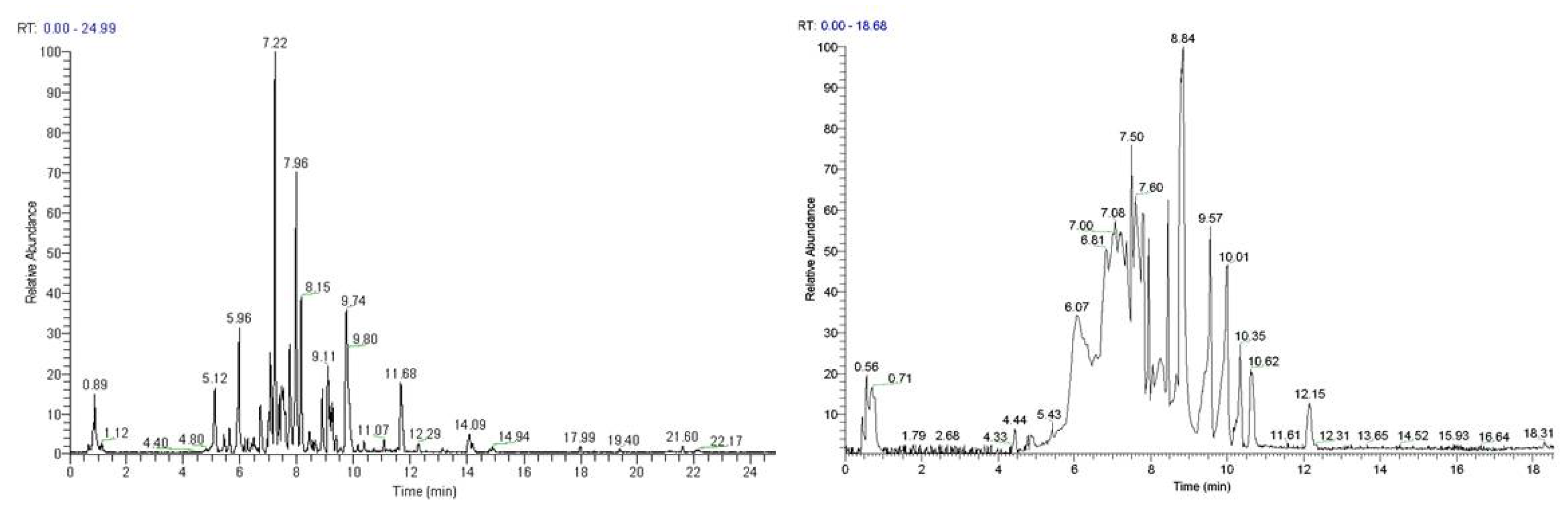
3.11. LC-MS Analyses of Selected Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Sklirou, A.; Papanagnou, E.-D.; Fokialakis, N.; Trougakos, I.P. Cancer chemoprevention via activation of proteostatic modules. Cancer Lett. 2018, 413, 110–121. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Cuervo, A.M. Proteostasis and the aging proteome in health and disease. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S33–S38. [Google Scholar] [CrossRef] [Green Version]
- Trougakos, I.P.; Sesti, F.; Tsakiri, E.; Gorgoulis, V.G. Non-enzymatic post-translational protein modifications and proteostasis network deregulation in carcinogenesis. J. Proteom. 2013, 92, 274–298. [Google Scholar] [CrossRef] [PubMed]
- Niforou, K.; Cheimonidou, C.; Trougakos, I.P. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol. 2014, 2, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Gumeni, S.; Evangelakou, Z.; Gorgoulis, V.G.; Trougakos, I.P. Proteome Stability as a Key Factor of Genome Integrity. Int. J. Mol. Sci. 2017, 18, 2036. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, S.; Cuervo, A.M. Proteostasis and aging. Nat. Med. 2015, 21, 1406–1415. [Google Scholar] [CrossRef]
- Vilchez, D.; Saez, I.; Dillin, A. The role of protein clearance mechanisms in organismal ageing and age-related diseases. Nat. Commun. 2014, 5, 5659. [Google Scholar] [CrossRef]
- Sala, A.J.; Bott, L.C.; Morimoto, R.I. Shaping proteostasis at the cellular, tissue, and organismal level. J. Cell Biol. 2017, 216, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Tsakiri, E.N.; Trougakos, I.P. The amazing ubiquitin-proteasome system: Structural components and implication in aging. Int. Rev. Cell Mol. Biol. 2015, 314, 171–237. [Google Scholar] [CrossRef] [PubMed]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavker, R.M.; Zheng, P.S.; Dong, G. Morphology of aged skin. Clin. Geriatr. Med. 1989, 5, 53–67. [Google Scholar] [CrossRef]
- Meyer, L.J.; Stern, R. Age-dependent changes of hyaluronan in human skin. J. Investig. Dermatol. 1994, 102, 385–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, L.; Labat-Robert, J.; Robert, A.M. Physiology of skin aging. Pathol. Biol. 2009, 57, 336–341. [Google Scholar] [CrossRef]
- Rijken, F.; Kiekens, R.C.; Bruijnzeel, P.L. Skin-infiltrating neutrophils following exposure to solar-simulated radiation could play an important role in photoageing of human skin. Br. J. Dermatol. 2005, 152, 321–328. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases: From induction via signaling to initial events. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef]
- Takeuchi, H.; Gomi, T.; Shishido, M.; Watanabe, H.; Suenobu, N. Neutrophil elastase contributes to extracellular matrix damage induced by chronic low-dose UV irradiation in a hairless mouse photoaging model. J. Dermatol. Sci. 2010, 60, 151–158. [Google Scholar] [CrossRef]
- Argyropoulou, A.; Aligiannis, N.; Trougakos, I.P.; Skaltsounis, A.-L. Natural compounds with anti-ageing activity. Nat. Prod. Rep. 2013, 30, 1412–1437. [Google Scholar] [CrossRef]
- Wedel, S.; Manola, M.; Cavinato, M.; Trougakos, I.P.; Jansen-Dürr, P. Targeting Protein Quality Control Mechanisms by Natural Products to Promote Healthy Ageing. Molecules 2018, 23, 1219. [Google Scholar] [CrossRef] [Green Version]
- Evangelakou, Z.; Manola, M.; Gumeni, S.; Trougakos, I.P. Nutrigenomics as a tool to study the impact of diet on aging and age-related diseases: The Drosophila approach. Genes Nutr. 2019, 14, 12. [Google Scholar] [CrossRef]
- Saewan, N.; Jimtaisong, A. Natural products as photoprotection. J. Cosmet. Dermatol. 2015, 14, 47–63. [Google Scholar] [CrossRef]
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Guldbrandsen, N.; De Mieri, M.; Gupta, M.; Liakou, E.; Pratsinis, H.; Kletsas, D.; Chaita, E.; Aligiannis, N.; Skaltsounis, A.-L.; Hamburger, M. Screening of Panamanian plants for cosmetic properties, and HPLC-based identification of constituents with antioxidant and UV-B protecting activities. Sci. Pharm. 2015, 83. [Google Scholar] [CrossRef] [Green Version]
- Reid, W.V. Biodiversity hotspots. Trends Ecol. Evol. 1998, 13, 275–280. [Google Scholar] [CrossRef]
- Lee, S.K.; Mbwambo, Z.H.; Chung, H.; Luyengi, L.; Gamez, E.J.C.; Mehta, R.G.; Kinghorn, A.D.; Pezzuto, J.M. Evaluation of the antioxidant potential of natural products. Comb. Chem. High. Throughput Screen. 1998, 1, 35–46. [Google Scholar] [PubMed]
- Masuda, T.; Yamashita, D.; Takeda, Y.; Yonemori, S. Screening for tyrosinase inhibitors among extracts of seashore plants and identification of potent inhibitors from Garcinia subelliptica. Biosci. Biotechnol. Biochem. 2005, 69, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Algul, O.; Kaessler, A.; Apcin, Y.; Yilmaz, A.; Jose, J. Comparative studies on conventional and microwave synthesis of some benzimidazole, benzothiazole and indole derivatives and testing on inhibition of hyaluronidase. Molecules 2008, 13, 736–748. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.C.; Taori, K.; Paul, V.J.; Luesch, H. Lyngbyastatins 8-10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sklirou, A.D.; Ralli, M.; Dominguez, M.; Papassideri, I.; Skaltsounis, A.L.; Trougakos, I.P. Hexapeptide-11 is a novel modulator of the proteostasis network in human diploid fibroblasts. Redox Biol. 2015, 5, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Chaita, E.; Lambrinidis, G.; Cheimonidi, C.; Agalou, A.; Beis, D.; Trougakos, I.; Mikros, E.; Skaltsounis, A.L.; Aligiannis, N. Anti-melanogenic properties of Greek plants. A novel depigmenting agent from Morus alba wood. Molecules 2017, 22, 514. [Google Scholar] [CrossRef] [Green Version]
- Sklirou, A.D.; Gaboriaud-Kolar, N.; Papassideri, I.; Skaltsounis, A.-L.; Trougakos, I.P. 6-bromo-indirubin-3’-oxime (6BIO), a Glycogen synthase kinase-3β inhibitor, activates cytoprotective cellular modules and suppresses cellular senescence-mediated biomolecular damage in human fibroblasts. Sci. Rep. 2017, 7, 11713. [Google Scholar] [CrossRef] [Green Version]
- Beekman, B.; Drijfhout, J.W.; Bloemhoff, W.; Ronday, H.K.; Tak, P.P.; te Koppele, J.M. Convenient fluorometric assay for matrix metalloproteinase activity and its application in biological media. FEBS Lett. 1996, 390, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Anastasiadi, M.; Pratsinis, H.; Kletsas, D.; Skaltsounis, A.-L.; Haroutounian, S.A. Bioactive non-coloured polyphenols content of grapes, wines and vinification by-products: Evaluation of the antioxidant activities of their extracts. Food Res. Int. 2010, 43. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX phosphorylation: A marker for DNA damage. Methods Mol. Biol. 2012, 920, 613–626. [Google Scholar] [CrossRef]
- Pratsinis, H.; Armatas, A.; Dimozi, A.; Lefaki, M.; Vassiliu, P.; Kletsas, D. Paracrine anti-fibrotic effects of neonatal cells and living cell constructs on young and senescent human dermal fibroblasts. Wound Repair Regen. 2013, 21, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Barberan, F.A.; Tomas, F.; Ferreres, F. Isoscutellarein-7-O-[allosyl (1-2) glucoside] from Sideritis leucantha. J. Nat. Prod. 1985, 48, 28–32. [Google Scholar] [CrossRef]
- Petreska, J.; Stefkov, G.; Kulevanova, S.; Alipieva, K.; Bankova, V.; Stefova, M. Phenolic compounds of mountain tea from the Balkans: LC/DAD/ESI/MSn profile and content. Nat. Prod. Commun. 2011, 6, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturella, P.; Bellino, A.; Marino, M.L. Three acylated flavone glycosides from Sideritis syriaca. Phytochemistry 1995, 38, 527–530. [Google Scholar] [CrossRef]
- Avila Acevedo, J.G.; Espinosa Gonzalez, A.M.; De Maria y Campos, D.M.; Benitez Flores Jdel, C.; Hernandez Delgado, T.; Flores Maya, S.; Campos Contreras, J.; Munoz Lopez, J.L.; Garcia Bores, A.M. Photoprotection of Buddleja cordata extract against UVB-induced skin damage in SKH-1 hairless mice. BMC Complement. Altern. Med. 2014, 14, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyuk, V.; Potapovich, A.; Suhan, T.; De Luca, C.; Pressi, G.; Dal Toso, R.; Korkina, L. Plant polyphenols against UV-C-induced cellular death. Planta Med. 2008, 74, 509–514. [Google Scholar] [CrossRef]
- Peng, X.M.; Gao, L.; Huo, S.X.; Liu, X.M.; Yan, M. The mechanism of memory enhancement of acteoside (verbascoside) in the senescent mouse model induced by a combination of D-gal and AlCl3. Phytother. Res. 2015, 29, 1137–1144. [Google Scholar] [CrossRef]
- Zivkovic, J.C.; Barreira, J.C.M.; Savikin, K.P.; Alimpic, A.Z.; Stojkovic, D.S.; Dias, M.I.; Santos-Buelga, C.; Duletic-Lausevic, S.N.; Ferreira, I. Chemical profiling and assessment of antineurodegenerative and antioxidant properties of Veronica teucrium L. and Veronica jacquinii Baumg. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef] [Green Version]
- Schriner, S.E.; Katoozi, N.S.; Pham, K.Q.; Gazarian, M.; Zarban, A.; Jafari, M. Extension of Drosophila lifespan by Rosa damascena associated with an increased sensitivity to heat. Biogerontology 2012, 13, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Jafari, M.; Zarban, A.; Pham, S.; Wang, T. Rosa damascena decreased mortality in adult Drosophila. J. Med. Food 2008, 11, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodagam, L.; Lewinska, A.; Kwasniewicz, E.; Kokhanovska, S.; Wnuk, M.; Siems, K.; Rattan, S.I.S. Phytochemicals rosmarinic acid, ampelopsin, and amorfrutin-A can modulate age-related phenotype of serially passaged human skin fibroblasts in vitro. Front. Genet. 2019, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.J.; Kim, K.B.; An, I.S.; Ahn, K.J.; Han, H.J. Protective effects of rosmarinic acid against hydrogen peroxideinduced cellular senescence and the inflammatory response in normal human dermal fibroblasts. Mol. Med. Rep. 2017, 16, 9763–9769. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, E.; Younis, I.Y.; Farag, M.A. Metabolites profiling of Egyptian Rosa damascena Mill. flowers as analyzed via ultra-high-performance liquid chromatography-mass spectrometry and solid-phase microextraction gas chromatography-mass spectrometry in relation to its anti-collagenase skin effect. Ind. Crops Prod. 2020, 1551, 112818. [Google Scholar]
- Solimine, J.; Garo, E.; Wedler, J.; Rusanov, K.; Fertig, O.; Hamburger, M.; Atanassov, I.; Butterweck, V. Tyrosinase inhibitory constituents from a polyphenol enriched fraction of rose oil distillation wastewater. Fitoterapia 2016, 108, 13–19. [Google Scholar] [CrossRef]
- Lee, Y.R.; Cho, H.M.; Park, E.J.; Zhang, M.; Doan, T.P.; Lee, B.W.; Cho, K.A.; Oh, W.K. Metabolite profiling of Rambutan (Nephelium lappaceum L.) seeds using UPLC-qTOF-MS/MS and senomorphic effects in aged human dermal fibroblasts. Nutrients 2020, 12, 1430. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Code | Highest Non-Cytotoxic Concentration (μg/mL) |
|---|---|---|
| Satureja hortensis | EXT1 | 100 |
| Origanum majorana | EXT2 | 20 |
| Origanum vulgare subsp. hirtum | EXT3 | 100 |
| Hyssopus officinalis | EXT4 | 20 |
| Salvia fruticosa | EXT5 | 4 |
| Salvia pomifera | EXT6 | 20 |
| Salvia officinalis | EXT7 | 20 |
| Salvia sclarea | EXT8 | 20 |
| Sideritis scardica | EXT9 | 20 |
| Melissa officinalis | EXT10 | 100 |
| Thymus vulgaris | EXT11 | 20 |
| Hypericum perforatum | EXT12 | 4 |
| Glycyrrhiza glabra | EXT13 | 20 |
| Geranium macrorrhizum | EXT14 | 100 |
| Umbilicus horizontalis | EXT15 | 100 |
| Sedum sediforme | EXT16 | 100 |
| Paeonia mascula | EXT17 | 0.16 |
| Rosa damascena | EXT18 | 20 |
| Extract | Hyaluronidase Inhibition (% at 300 μg/mL) | Elastase Inhibition (% at 300 μg/mL) |
|---|---|---|
| Geranium macrorrhizum | 51.09 | 46.64 |
| Glycyrrhiza glabra | 35.81 | 48.75 |
| Hypericum perforatum | 8.86 | −4.24 |
| Hyssopus officinalis | 24.73 | 28.20 |
| Melissa officinalis | −0.98 | 58.02 |
| Origanum majorana | 11.27 | 41.14 |
| Origanum vulgare subsp. hirtum | 81.04 | 1.01 |
| Paeonia mascula | 99.67 | 5.80 |
| Rosa damascena | 27.52 | 17.28 |
| Salvia officinalis | 8.57 | 55.31 |
| Salvia pomifera | 23.07 | 40.40 |
| Salvia sclarea | −6.01 | −14.56 |
| Salvia fruticosa | 28.93 | 42.71 |
| Satureja hortensis | 52.92 | −26.38 |
| Sedum sediforme | 98.49 | 89.07 |
| Sideritis scardica | 13.95 | −19.95 |
| Thymus vulgaris | 31.07 | 31.18 |
| Umbilicus horizontalis | 89.04 | 87.84 |
| No. | Compounds | Rt (min) | Theoretical | Experimental | Δm (ppm) | RDBeq | Molecular Formula |
|---|---|---|---|---|---|---|---|
| [M-H]− m/z | |||||||
| 1 | Feruloylquinic acid derivative | 0.84 | 367.1035 | 367.1021 | 0.310 | 8.5 | C17H20O9 |
| 2 | Melittoside derivative | 1.12 | 569.1719 | 569.1709 | 0.069 | 15.5 | C29H30O12 |
| 3 | 5-Caffeoylquinic acid | 5.96 | 353.0870 | 353.0855 | 0.259 | 8.5 | C16H18O9 |
| 4 | Apigenin 7-O-allosyl(1→2)-glucoside | 6.28 | 593.1492 | 593.1472 | 0.189 | 13.5 | C27H30O15 |
| 5 | Echinacoside | 7.01 | 785.2484 | 785.2475 | 0.465 | 13.5 | C35H46O20 |
| 6 | Lavandulifolioside | 7.08 | 755.2380 | 755.2360 | 1.189 | 13.5 | C34H44O19 |
| 7 | Verbascoside | 7.22 | 623.1981 | 623.1981 | 0.598 | 12.5 | C29H36O15 |
| 8 | Hypolaetin 7-O-allosyl(1→2)-glucoside | 7.48 | 625.2020 | 625.2015 | 1.100 | 13.5 | C31H34O18 |
| 9 | Samioside | 7.08 | 755.2404 | 755.2402 | 0.254 | 13.5 | C34H44O19 |
| 10 | Isoscutellarein 7-O-allosyl(1→2)-glucoside | 7.38 | 609.1461 | 609.1459 | 0.456 | 13.5 | C27H30O16 |
| 11 | Allysonoside | 7.59 | 769.2561 | 769.2555 | 0.278 | 13.5 | C35H46O19 |
| 12 | Hypolaetin 7-O-[6‴-O-acetyl]-allosyl(1→2)-glucoside | 7.52 | 667.1516 | 667.1511 | 0.512 | 14.5 | C29H32O18 |
| 13 | Leucosceptoside A | 7.74 | 637.2138 | 637.2125 | 0.357 | 12.5 | C30H38O15 |
| 14 | 3′-O-Methylhypolaetin 7-O-allosyl(1→2)-glucoside | 7.56 | 639.1567 | 639.1561 | 0.632 | 13.5 | C28H32O17 |
| 15 | Apigenin 7-O-glucoside | 7.79 | 431.0984 | 431.0979 | 0.558 | 12.5 | C21H20O10 |
| 16 | Isoscutellarein 7-O-[6‴-O-acetyl]-allosyl(1→2)-glucoside | 7.96 | 653.1618 | 653.1612 | 0.234 | 13.5 | C29H32O17 |
| 17 | Apigenin 7-O-[6‴-O-acetyl]-allosyl(1→2)-glucoside | 7.88 | 653.1618 | 635.1610 | 0.589 | 14.5 | C29H32O16 |
| 18 | Isoscutellarein 7-O-allosyl-(1→2)-[6″-O-acetyl]-glucoside | 7.96 | 651.1567 | 651.1565 | 0.676 | 14.5 | C29H32O17 |
| 19 | 4′-O-Methylhypolaetin 7-O-[6‴-O-acetyl] -allosyl(1→2)-glucoside | 8.15 | 681.1672 | 681.1669 | 1.347 | 14.5 | C30H34O18 |
| 20 | 3′-O-Methylhypolaetin 7-O-[6‴-O-acetyl]-allosyl-(1→2)-[6″-O-acetyl]-glucoside or 4′-O-Methylhypolaetin 7-O-[6‴-O-acetyl] -allosyl-(1→2) [6″-O-acetyl]-glucoside | 9.27 | 723.1767 | 723.1765 | 0.457 | 15.5 | C32H36O19 |
| 21 | 4′-O-Methylisoscutellarein 7-O-[6‴-O-acetyl]-allosyl(1→2)-glucoside | 9.15 | 665.1723 | 665.1720 | 0.968 | 14.5 | C30H34O17 |
| 22 | Isoscutellarein 7-O-[6‴-O-acetyl]-allosyl(1→2)-[6″-O-acetyl]-glucoside | 9.11 | 693.1672 | 693.1668 | 0.436 | 15.5 | C31H34O18 |
| 23 | Apigenin | 9.78 | 269.0455 | 269.0450 | 0.969 | 11.5 | C15H10O5 |
| 24 | Apigenin 7-(6″-p-coumaroylglucoside) or Apigenin 7-(4″-p-coumaroylglucoside) | 9.74 | 577.1351 | 577.1348 | 0.336 | 18.5 | C30H26O12 |
| No. | Compounds | Rt (min) | Theoretical | Experimental | Δm (ppm) | RDBeq | Molecular Formula |
|---|---|---|---|---|---|---|---|
| [M-H]− m/z | |||||||
| 1 | Quinic acid | 0.65 | 165.0405 | 165.0408 | 1.972 | 1.5 | C7H12O6 |
| 2 | Gallic acid | 2.69 | 169.0142 | 169.0147 | 0.753 | 5.5 | C7H6O5 |
| 3 | Protocatechuic acid | 4.70 | 153.0193 | 153.0196 | 2.000 | 5.5 | C7H6O5 |
| 4 | Quercetin 3-O-glucoside | 6.07 | 463.0882 | 463.0885 | 0.718 | 12.5 | C21H20O12 |
| 5 | Rutin | 6.57 | 609.1461 | 609.1462 | 0.217 | 13.5 | C27H30O16 |
| 6 | Quercetin 3-O-pentoside | 7.04 | 433.0776 | 433.0778 | 0.357 | 12.5 | C20H18O11 |
| 7 | Hyperoside | 7.08 | 463.0882 | 463.0883 | 0.322 | 12.5 | C21H20O12 |
| 8 | Quercetin-glucuronide | 7.08 | 477.0675 | 477.0674 | −0.042 | 13.5 | C21H18O13 |
| 9 | Quercetin 3-O-rhamnoside | 7.22 | 447.0933 | 447.0934 | 0.427 | 12.5 | C21H20O11 |
| 10 | Kaempferol 3-O-glucoside | 7.22 | 447.0933 | 447.0934 | 0.1537 | 12.5 | C21H20O11 |
| 11 | Kaempferol-3-O-rutinoside | 7.26 | 593.1512 | 593.1513 | 0.190 | 13.5 | C27H30O15 |
| 12 | Rosmarinic acid | 7.50 | 359.0772 | 359.0773 | 0.253 | 11.5 | C18H16O8 |
| 13 | Caffeic acid | 7.60 | 179.0350 | 179.0354 | 2.251 | 6.5 | C9H8O4 |
| 14 | Kaempferol pentoside | 7.70 | 417.0827 | 417.0828 | 0.178 | 12.5 | C20H18O10 |
| 15 | Kaempferol-3-O-rhamnoside | 8.05 | 431.0984 | 431.0984 | 0.1142 | 12.5 | C21H20O10 |
| 16 | Kaempferol 3-O-(6″-O-E-π-coumaroyl)-β-D-glucopyranoside | 8.84 | 593.1301 | 593.1302 | 0.209 | 18.5 | C30H26O13 |
| 17 | Quercetin | 8.95 | 301.0354 | 301.0354 | 0.183 | 11.5 | C15H10O7 |
| 18 | Kaempferol | 9.57 | 285.0405 | 285.0404 | −0.197 | 11.5 | C15H10O6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sklirou, A.D.; Angelopoulou, M.T.; Argyropoulou, A.; Chaita, E.; Boka, V.I.; Cheimonidi, C.; Niforou, K.; Mavrogonatou, E.; Pratsinis, H.; Kalpoutzakis, E.; et al. Phytochemical Study and In Vitro Screening Focusing on the Anti-Aging Features of Various Plants of the Greek Flora. Antioxidants 2021, 10, 1206. https://doi.org/10.3390/antiox10081206
Sklirou AD, Angelopoulou MT, Argyropoulou A, Chaita E, Boka VI, Cheimonidi C, Niforou K, Mavrogonatou E, Pratsinis H, Kalpoutzakis E, et al. Phytochemical Study and In Vitro Screening Focusing on the Anti-Aging Features of Various Plants of the Greek Flora. Antioxidants. 2021; 10(8):1206. https://doi.org/10.3390/antiox10081206
Chicago/Turabian StyleSklirou, Aimilia D., Maria T. Angelopoulou, Aikaterini Argyropoulou, Eliza Chaita, Vasiliki Ioanna Boka, Christina Cheimonidi, Katerina Niforou, Eleni Mavrogonatou, Harris Pratsinis, Eleftherios Kalpoutzakis, and et al. 2021. "Phytochemical Study and In Vitro Screening Focusing on the Anti-Aging Features of Various Plants of the Greek Flora" Antioxidants 10, no. 8: 1206. https://doi.org/10.3390/antiox10081206
APA StyleSklirou, A. D., Angelopoulou, M. T., Argyropoulou, A., Chaita, E., Boka, V. I., Cheimonidi, C., Niforou, K., Mavrogonatou, E., Pratsinis, H., Kalpoutzakis, E., Aligiannis, N., Kletsas, D., Trougakos, I. P., & Skaltsounis, A. L. (2021). Phytochemical Study and In Vitro Screening Focusing on the Anti-Aging Features of Various Plants of the Greek Flora. Antioxidants, 10(8), 1206. https://doi.org/10.3390/antiox10081206