
Taxifolin and Sorghum Ethanol Extract Protect against Hepatic Insulin Resistance via the miR-195/IRS1/PI3K/AKT and AMPK Signalling Pathways



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
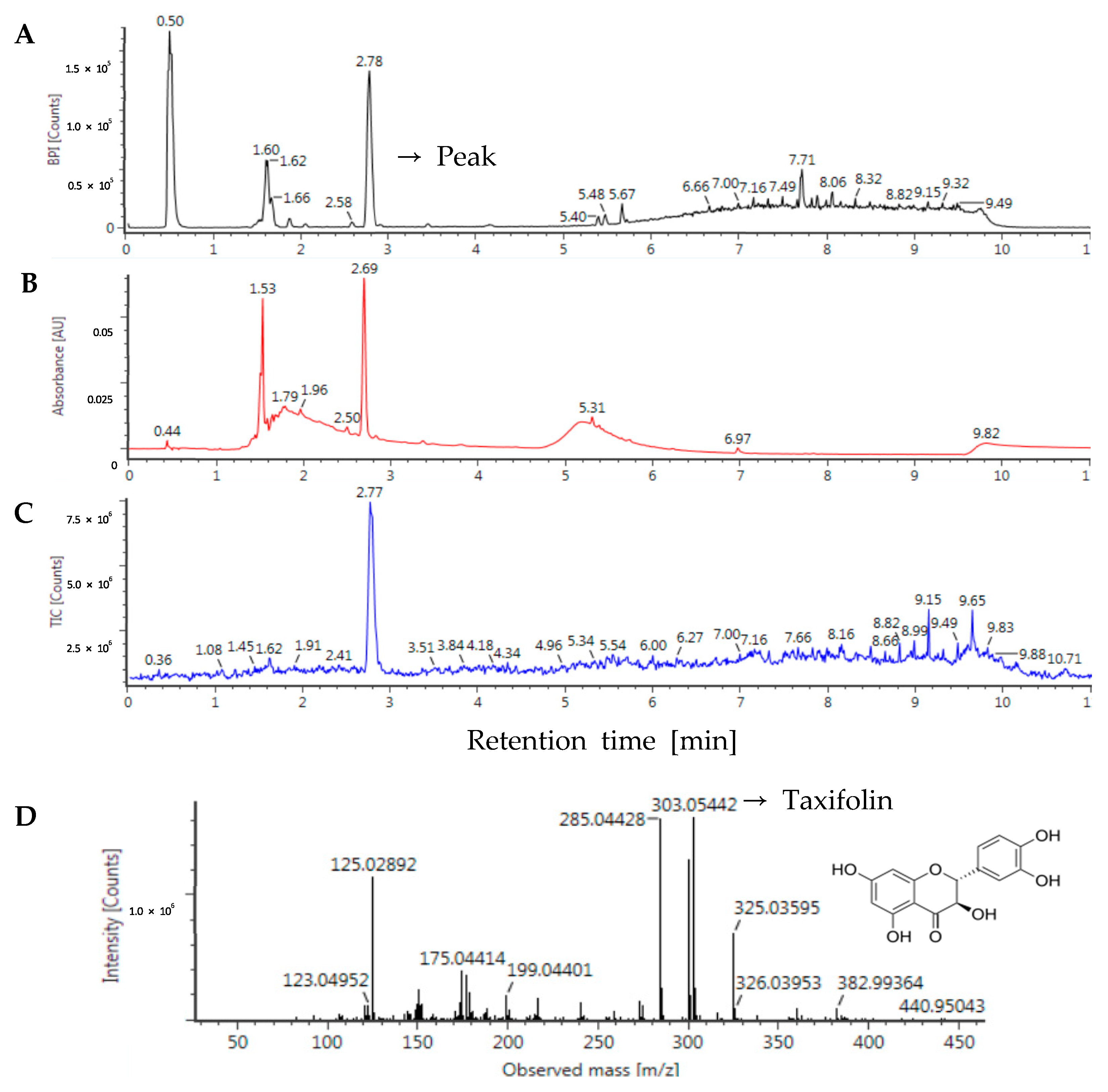
2.2. Ultra-Performance Liquid Chromatography-Electrospray Ionization Quadrupole Time-of-Flight Mass Spectrometry (UPLC-ESI-Q-TOF/MS)
2.3. Free Fatty Acid Mixture Preparation
2.4. Cell Culture and Cytotoxicity
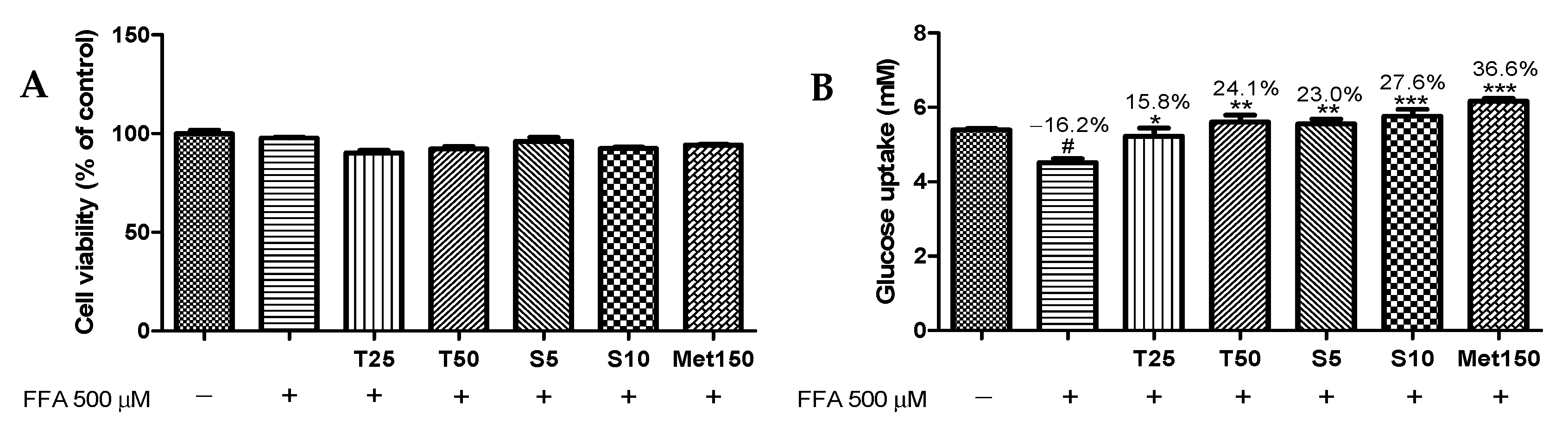
2.5. Glucose Uptake
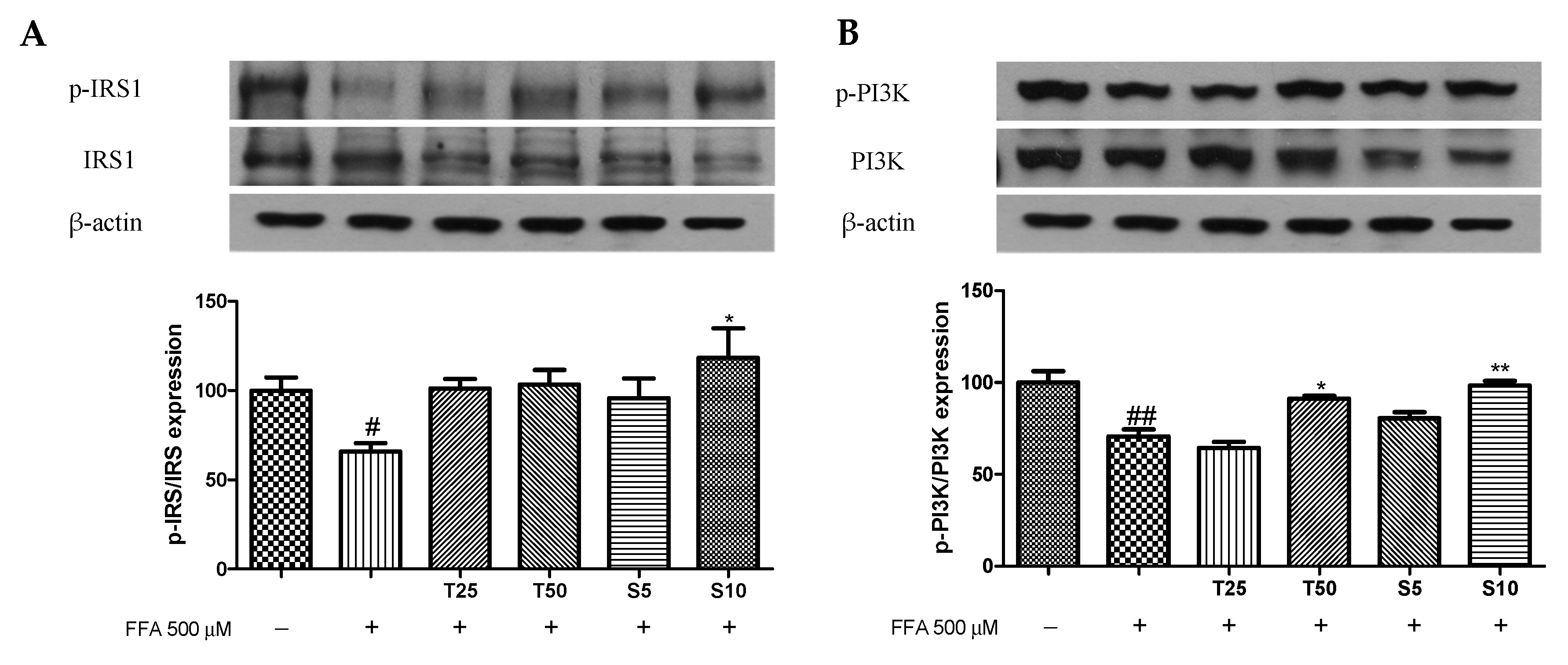
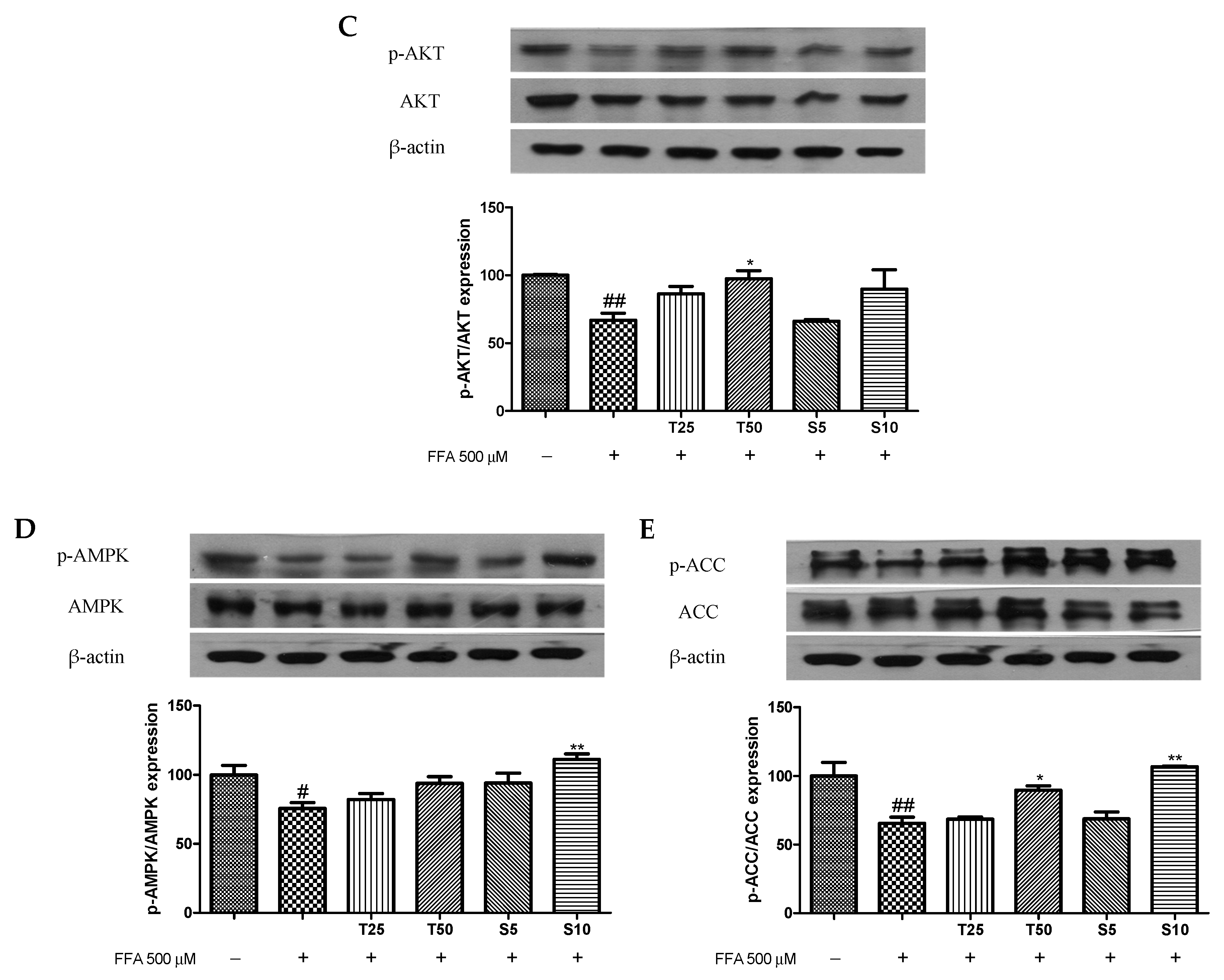
2.6. Western Blotting
2.7. Transfection
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results and Discussion
3.1. Identification of Taxifolin in Sorghum Ethanol Extract
3.2. Effects of Taxifolin and Sorghum Ethanol Extract on Cell Viability and Glucose Uptake
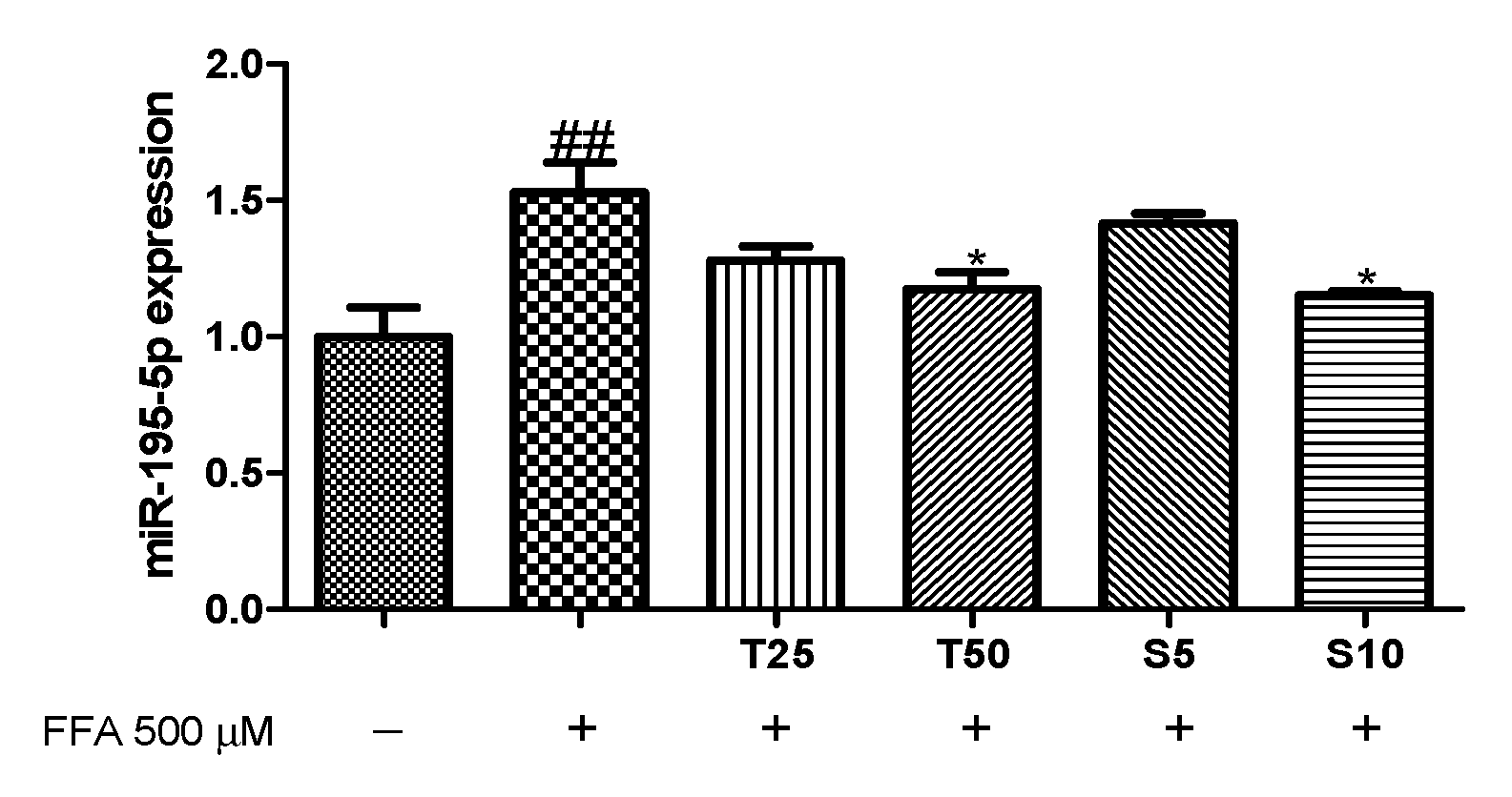
3.3. Effects of Taxifolin and Sorghum Ethanol Extract on the IRS1/PI3K/AKT and AMPK Signalling Pathways
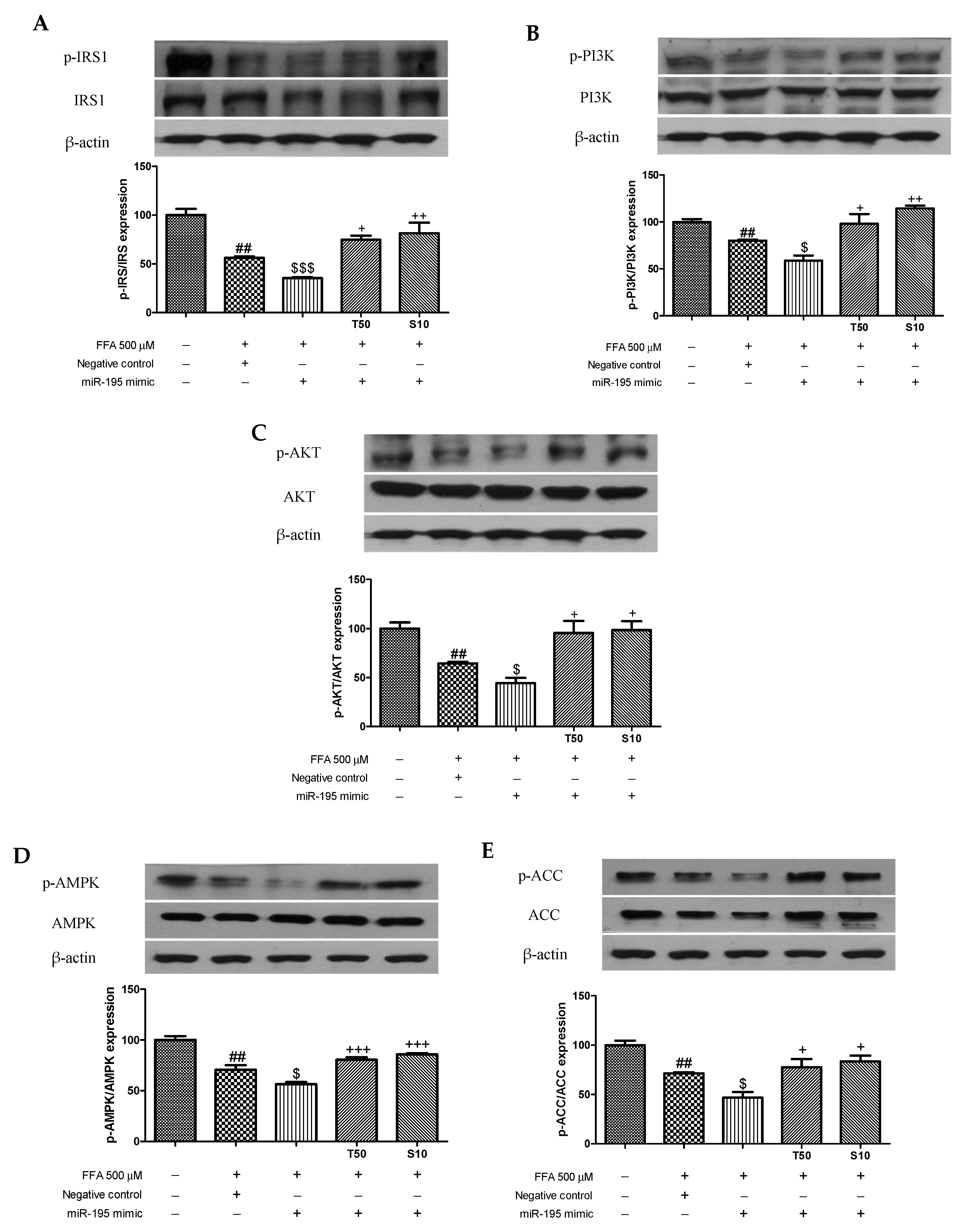
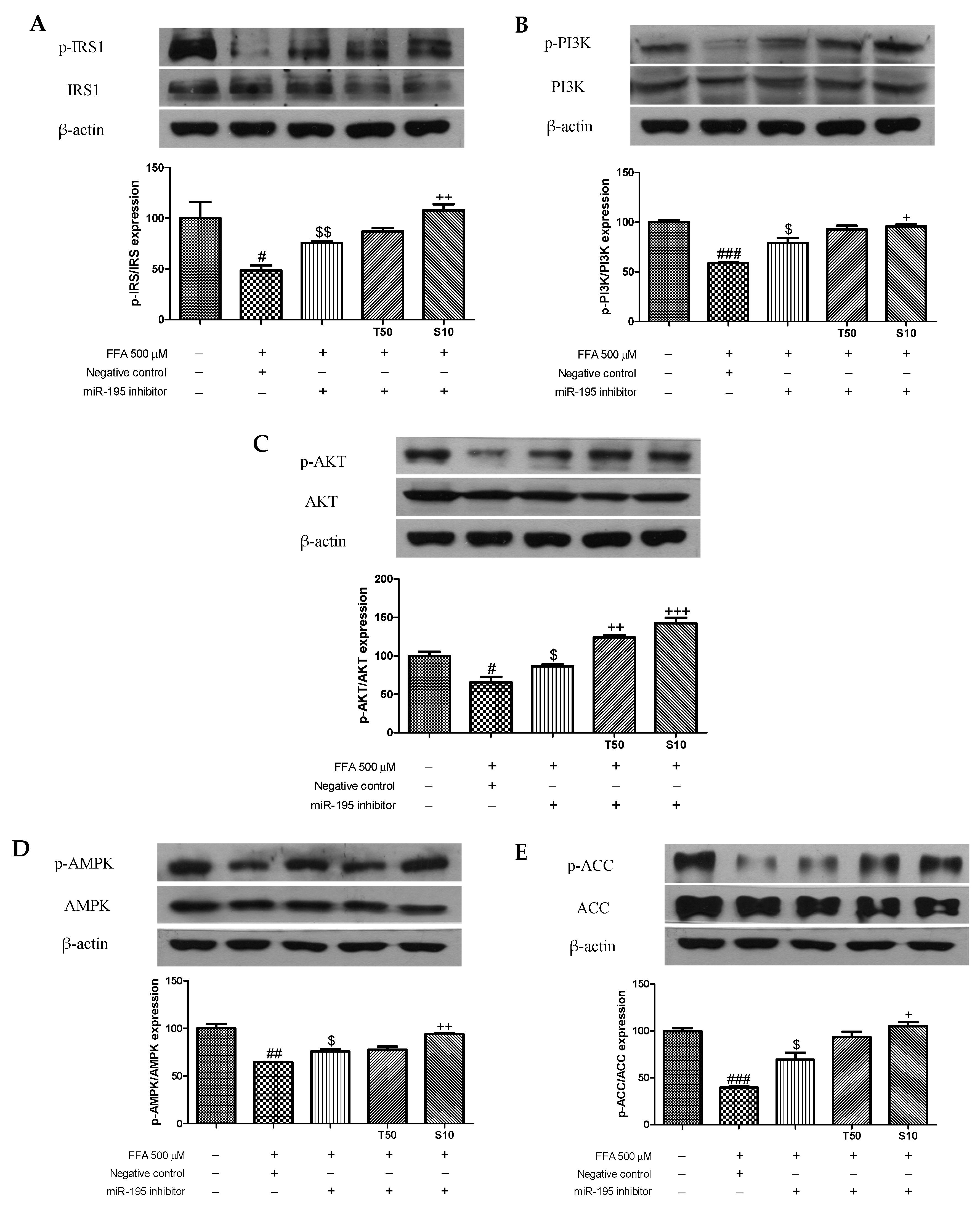
3.4. Effects of Taxifolin and Sorghum Ethanol Extract on IRS1, PI3K, AKT, AMPK, and ACC Phosphorylation in miR-195 Mimic- and Inhibitor-Transfected Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, S.Y.; Huang, R.; Li, C.; Wu, W.Y.; Chen, H.L.; Shi, J.; Chen, S.G.; Ye, X.Q. Phenolic compositions and antioxidant activities differ significantly among sorghum grains with different applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.-H.; Dung, V.V.; Zhao, B.T.; Kim, Y.H.; Min, B.S.; Woo, M.H. Antithrombotic and antidiabetic flavonoid glycosides from the grains of Sorghum bicolor (L.) Moench var. hwanggeumchal. Arch. Pharmacal Res. 2014, 37, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Poquette, N.M.; Gu, X.; Lee, S.-O. Grain sorghum muffin reduces glucose and insulin responses in men. Food Funct. 2014, 5, 894–899. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum grain: From genotype, nutrition, and phenolic profile to its health benefits and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotnikov, M.B.; Aliev, O.; Sidekhmenova, A.; Shamanaev, A.; Anishchenko, A.; Nosarev, A.; Pushkina, E.A. Modes of hypotensive action of dihydroquercetin in arterial hypertension. Bull. Exp. Biol. Med. 2017, 162, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Johnson, S.; Bornman, J.F.; Bennett, S.J.; Fang, Z. Changes in whole grain polyphenols and antioxidant activity of six sorghum genotypes under different irrigation treatments. Food Chem. 2017, 214, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, J.; Dong, X.-D.; Ji, H.-Y. Research on characteristics, antioxidant and antitumor activities of dihydroquercetin and its complexes. Molecules 2017, 23, 20. [Google Scholar] [CrossRef] [Green Version]
- Makena, P.S.; Pierce, S.C.; Chung, K.T.; Sinclair, S.E. Comparative mutagenic effects of structurally similar flavonoids quercetin and taxifolin on tester strains salmonella typhimurium TA102 and escherichia coli WP-2 uvrA. Environ. Mol. Mutagen. 2009, 50, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S. The potential protective action of vitamin d in hepatic insulin resistance and pancreatic islet dysfunction in type 2 diabetes mellitus. Nutrients 2016, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yuan, P.; Zhang, Q.; Fu, Y.; Hou, Y.; Wei, Y.; Zheng, X.; Feng, W. Taxifolin improves disorders of glucose metabolism and water-salt metabolism in kidney via PI3K/AKT signaling pathway in metabolic syndrome rats. Life Sci. 2020, 263, 118713. [Google Scholar] [CrossRef] [PubMed]
- Teselkin, Y.O.; Babenkova, I.V.; Kolhir, V.K.; Baginskaya, A.I.; Tjukavkina, N.A.; Kolesnik, Y.A.; Selivanova, I.; Eichholz, A.A. Dihydroquercetin as a means of antioxidative defence in rats with tetrachloromethane hepatitis. Phytother. Res. 2000, 14, 160–162. [Google Scholar] [CrossRef]
- Hu, X.; Wang, S.; Xu, J.; Wang, D.B.; Chen, Y.; Yang, G.Z. Triterpenoid saponins from Stauntonia chinensis ameliorate insulin resistance via the AMP-activated protein kinase and IR/IRS-1/PI3K/Akt pathways in insulin-resistant HepG2 cells. Int. J. Mol. Sci. 2014, 15, 10446–10458. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, R.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2017, 19, 31–44. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ogawa, W.; Teshigawara, K.; Inoue, H.; Miyake, K.; Sakaue, H.; Kasuga, M. Role of the insulin receptor substrate 1 and phosphatidylinositol 3-kinase signaling pathway in insulin-induced expression of sterol regulatory element binding protein 1c and glucokinase genes in rat hepatocytes. Diabetes 2002, 51, 1672–1680. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Lai, F.Y.; Shi, L.S.; Chou, Y.C.; Yen, I.C.; Chang, T.C. Rhodiola crenulata extract suppresses hepatic gluconeogenesis via activation of the AMPK pathway. Phytomedicine 2015, 22, 477–486. [Google Scholar] [CrossRef]
- Hao, J.; Huang, K.P.; Chen, C.; Liang, Y.; Wang, Y.; Zhan, X.J.; Huang, H.Q. Polydatin improves glucose and lipid metabolisms in insulin resistant HepG2 cells through the AMPK Pathway. Biol. Pharm. Bull. 2018, 41, 891–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.D.; Mitchell, G.M. MicroRNAs in insulin resistance and obesity. Exp. Diabetes Res. 2012, 2012, 484696. [Google Scholar] [CrossRef]
- Ling, H.-Y.; Ou, H.-S.; Feng, S.-D.; Zhang, X.-Y.; Tuo, Q.-H.; Chen, L.-X.; Zhu, B.-Y.; Gao, Z.-P.; Tang, C.-K.; Yin, W.-D.; et al. Changes in microRNA (miR) profile and effects of miR-320 in insulin-resistant 3T3-L1 adipocytes. Clin. Exp. Pharmacol. Physiol. 2009, 36, e32–e39. [Google Scholar] [CrossRef]
- Shen, Y.F.; Liu, M.M.; Chen, C.; Lai, X.Y.; Zhang, M.Y. MicroRNA profile changes in liraglutide treated steatotic HepG2 cells. Int. J. Clin. Exp. Med. 2017, 10, 4856–4864. [Google Scholar]
- Yang, W.-M.; Jeong, H.-J.; Park, S.-Y.; Lee, W. Saturated fatty acid-induced miR-195 impairs insulin signaling and glycogen metabolism in HepG2 cells. FEBS Lett. 2014, 588, 3939–3946. [Google Scholar] [CrossRef]
- Herrera, B.M.; Lockstone, H.E.; Taylor, J.M.; Ria, M.; Barrett, A.; Collins, S.; Kaisaki, P.; Argoud, K.; Fernandez, C.; Travers, M.E.; et al. Global microRNA expression pro-files in insulin target tissues in a spontaneous rat model of type 2 diabetes. Diabetologia 2010, 53, 1099–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.T.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA profiles in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271–E2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, W.T.; Lim, H.B. A UPLC-ESI-Q-TOF method for rapid and reliable identification and quantification of major indole alkaloids in Catharanthus roseus. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1080, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Wobser, H.; Dorn, C.; Weiss, T.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid accumulation in hepatocytes induces fibrogenic activation of hepatic stellate cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Kong, X.; Yuan, H.; Guan, H.; Li, Y.; Niu, Y. Mangiferin improved palmitate-induced-insulin resistance by promoting free fatty acid metabolism in HepG2 and C2C12 cells via PPAR-α: Mangiferin improved insulin resistance. J. Diabetes Res. 2019, 2019, 2052675. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Sung, J.; Yang, J.; Ham, H.; Kim, Y.; Jeong, H.-S.; Lee, J. Inhibitory effect of esculetin on free-fatty-acid-induced lipid accumulation in human HepG2 cells through activation of AMP-activated protein kinase. Food Sci. Biotechnol. 2017, 26, 263–269. [Google Scholar] [CrossRef]
- Ding, X.; Jian, T.; Wu, Y.; Zuo, Y.; Li, J.; Lv, H.; Ma, L.; Ren, B.; Zhao, L.; Li, W.; et al. Ellagic acid ameliorates oxidative stress and insulin resistance in high glucose-treated HepG2 cells via miR-223/keap1-Nrf2 pathway. Biomed. Pharmacother. 2018, 110, 85–94. [Google Scholar] [CrossRef]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Syed Mohamad, S.N.A.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. BioMed Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2015, 30, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-D.; Lu, C.-J.; Zhao, R.-Z. Qualitative and quantitative analysis of rhizoma smilacis glabrae by ultra-high performance liquid chromatography coupled with LTQ OrbitrapXL hybrid mass spectrometry. Molecules 2014, 19, 10427–10439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of flavonoids in rhamnus davurica and its antiproliferative activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Yamamoto, Y.; Maki, T.; Hattori, Y.; Ito, H.; Mizuno, K.; Harada-Shiba, M.; Kalaria, R.N.; Fukushima, M.; Takahashi, R.; et al. Taxifolin inhibits amyloid beta oligomer formation and fully restores vascular integrity and memory in cerebral amyloid angiopathy. Acta. Neuropathol. Commun. 2017, 5, 26. [Google Scholar] [CrossRef]
- Yang, P.; Xu, F.; Li, H.-F.; Wang, Y.; Li, F.-C.; Shang, M.-Y.; Liu, G.-X.; Wang, X.; Cai, S.-Q. Detection of 191 taxifolin metabolites and their distribution in rats using HPLC-ESI-IT-TOF-MSn. Molecules 2016, 21, 1209. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.-M.; Kim, E.-H.; Yeo, M.-A.; Kim, S.-J.; Seo, M.; Moon, H.-I. Antidiabetic effects of three Korean sorghum phenolic extracts in normal and streptozotocin-induced diabetic rats. Food Res. Int. 2011, 44, 127–132. [Google Scholar] [CrossRef]
- Sadi, G.; Sahin, G.; Bostanci, A. Modulation of renal insulin signaling pathway and antioxidant enzymes with streptozotocin-induced diabetes: Effects of resveratrol. Medicina 2018, 55, 3. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, K.A.; Valentine, R.J.; Ruderman, N.B.; Saha, A.K. AMPK activation: A therapeutic target for type 2 diabetes? Diabetes Metab. Syndr. Obes. 2014, 7, 241–253. [Google Scholar]
- Kim, J.; Park, Y. Anti-diabetic effect of sorghum extract on hepatic gluconeogenesis of streptozotocin-induced diabetic rats. Nutr. Metab. 2012, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Luan, B.; Sun, C. MiR-138-5p affects insulin resistance to regulate type 2 diabetes progression through inducing autophagy in HepG2 cells by regulating SIRT1. Nutr. Res. 2018, 59, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Hou, Y.; Wu, J.; Li, G.; Cao, P.; Chen, W.; Hu, L.; Gan, D. MiR-93-5p promotes insulin resistance to regulate type 2 diabetes progression in HepG2 cells by targeting HGF. Mol. Med. Rep. 2021, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xue, X.-C.; Wang, Y.; Cao, F.-F.; You, J.; Uzan, G.; Peng, B.; Zhang, D.-H. Celastrol reverses palmitic acid-induced insulin resistance in HepG2 cells via restoring the miR-223 and GLUT4 pathway. Can. J. Diabetes 2018, 43, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Zhao, H.; Huang, W.; Hou, G.; Song, G.; Ma, H. Resveratrol upregulates mmu-miR-363-3p via the PI3K-Akt pathway to improve insulin resistance induced by a high-fat diet in mice. Diabetes Metab. Syndr. Obes. 2020, 13, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Lu, C.; Chu, W.; Zhang, B.; Zhen, Q.; Wang, R.; Zhang, Y.; Li, Z.; Lv, B.; Li, H.; et al. MicroRNA-124 suppresses proliferation and glycolysis in non–small cell lung cancer cells by targeting AKT–GLUT1/HKII. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Mortuza, R.; Feng, B.; Chakrabarti, S. miR-195 regulates SIRT1-mediated changes in diabetic retinopathy. Diabetologia 2014, 57, 1037–1046. [Google Scholar] [CrossRef]
- Chuang, J.C.; Jones, P.A. Epigenetics and microRNAs. Pediatr. Res. 2007, 61, 24R–29R. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Ørom, U.A.; Lund, A.H. Experimental identification of microRNA targets. Gene 2010, 451, 1–5. [Google Scholar] [CrossRef]
- Villeneuve, L.M.; Natarajan, R. The role of epigenetics in the pathology of diabetic complications. Am. J. Physiol. Physiol. 2010, 299, F14–F25. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, J.; Kumarswamy, R.; Dangwal, S.; Thum, T. MicroRNAs in diabetes and diabetes-associated complications. RNA Biol. 2012, 9, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantharidis, P.; Wang, B.; Carew, R.M.; Lan, H.Y. Diabetes complications: The microRNA perspective. Diabetes 2011, 60, 1832–1837. [Google Scholar] [CrossRef] [Green Version]
- Kang, H. MicroRNA-mediated health-promoting effects of phytochemicals. Int. J. Mol. Sci. 2019, 20, 2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.K.; Arora, S.; Averett, C.; Singh, S.; Singh, A.P. Modulation of microRNAs by phytochemicals in cancer: Underlying mechanisms and translational significance. BioMed Res. Int. 2015, 2015, 848710. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Jeong, W.-T.; So, Y.-S.; Lim, H.-B.; Lee, J. Taxifolin and Sorghum Ethanol Extract Protect against Hepatic Insulin Resistance via the miR-195/IRS1/PI3K/AKT and AMPK Signalling Pathways. Antioxidants 2021, 10, 1331. https://doi.org/10.3390/antiox10091331
Lee H, Jeong W-T, So Y-S, Lim H-B, Lee J. Taxifolin and Sorghum Ethanol Extract Protect against Hepatic Insulin Resistance via the miR-195/IRS1/PI3K/AKT and AMPK Signalling Pathways. Antioxidants. 2021; 10(9):1331. https://doi.org/10.3390/antiox10091331
Chicago/Turabian StyleLee, Hana, Won-Tae Jeong, Yoon-Sup So, Heung-Bin Lim, and Junsoo Lee. 2021. "Taxifolin and Sorghum Ethanol Extract Protect against Hepatic Insulin Resistance via the miR-195/IRS1/PI3K/AKT and AMPK Signalling Pathways" Antioxidants 10, no. 9: 1331. https://doi.org/10.3390/antiox10091331
APA StyleLee, H., Jeong, W. -T., So, Y. -S., Lim, H. -B., & Lee, J. (2021). Taxifolin and Sorghum Ethanol Extract Protect against Hepatic Insulin Resistance via the miR-195/IRS1/PI3K/AKT and AMPK Signalling Pathways. Antioxidants, 10(9), 1331. https://doi.org/10.3390/antiox10091331