
An Integrated NMR, LC-DAD-MS, LC-QTOF Metabolomic Characterization of Sartoria hedysaroides: Correlation of Antioxidant and Enzyme Inhibitory Activity with Chemical Composition by Multivariate Data Analysis

 ,
, 

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extraction
2.2. NMR Analysis
2.3. LC-DAD-ESI-MSn
2.4. UPLC-QTOF Analysis
2.5. Antioxidant and Enzyme Inhibitory Assays
2.6. Data Analysis
3. Results and Discussion
3.1. Phytochemical Characterization of S. hedysaroides Extracts
3.2. Antioxidant Effects
3.3. Inhibitory Activity of S. hedysaroides Extracts against Selected Enzymes
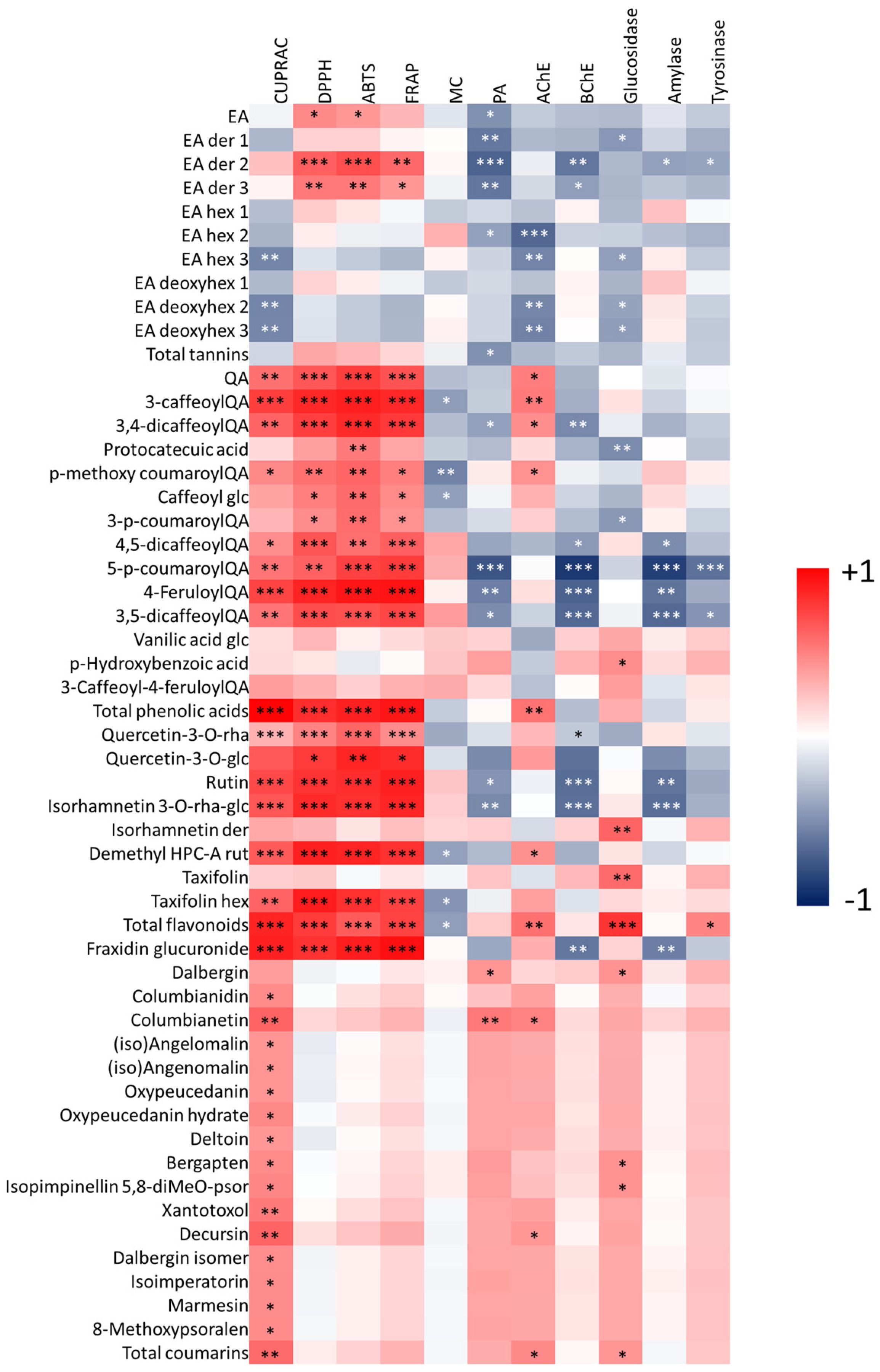
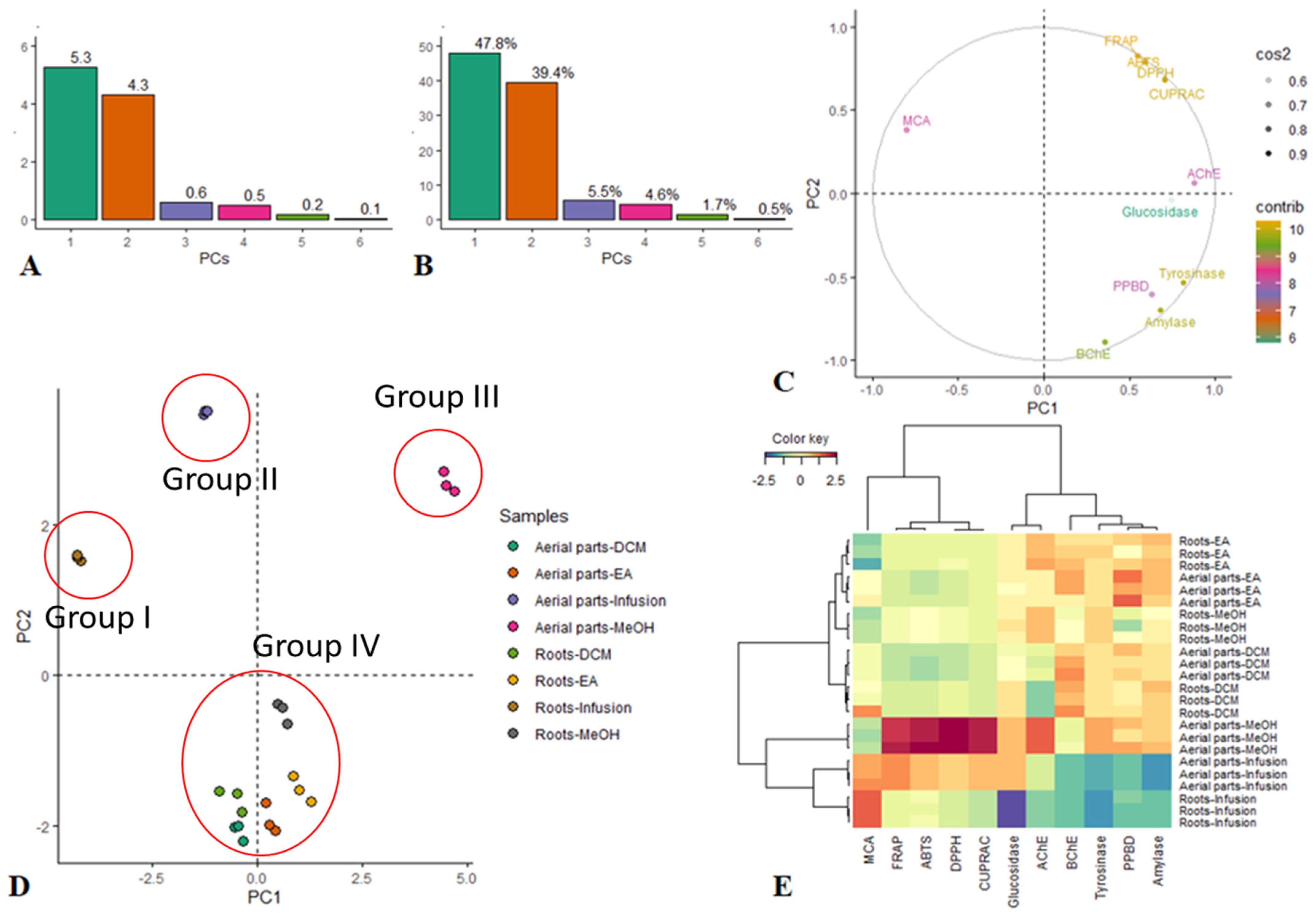
3.4. Multivariate Analysis of Biological Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polat, R. Ethnobotanical study on medicinal plants in Bingöl (City center) (Turkey). J. Herb. Med. 2019, 16, 100211. [Google Scholar] [CrossRef]
- Dalar, A.; Mukemre, M.; Unal, M.; Ozgokce, F. Traditional medicinal plants of Ağrı Province, Turkey. J. Ethnopharmacol. 2018, 226, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Macêdo, M.J.F.; Ribeiro, D.A.; Santos, M.D.; Macêdo, D.G.; Macedo, J.G.; Almeida, B.V.; Saraiva, M.E.; Lacerda, M.N.; Souza, M.M. Fabaceae medicinal flora with therapeutic potential in Savanna areas in the Chapada do Araripe, Northeastern Brazil. Rev. Bras. Farmacogn. 2018, 28, 738–750. [Google Scholar] [CrossRef]
- Gao, T.; Yao, H.; Song, J.; Liu, C.; Zhu, Y.; Ma, X.; Pang, X.; Xu, H.; Chen, S. Identification of medicinal plants in the family Fabaceae using a potential DNA barcode ITS2. J. Ethnopharmacol. 2010, 130, 116–121. [Google Scholar] [CrossRef]
- Sargın, S.A.; Akçicek, E.; Selvi, S. An ethnobotanical study of medicinal plants used by the local people of Alaşehir (Manisa) in Turkey. J. Ethnopharmacol. 2013, 150, 860–874. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, G.; Kamiloglu, S.; Ozdal, T.; Boyacioglu, D.; Capanoglu, E. Potential Use of Turkish Medicinal Plants in the Treatment of Various Diseases. Molecules 2016, 21, 257. [Google Scholar] [CrossRef]
- Arslan, E.; Ertuğrul, K.; Tugay, O.; Dural, H. Karyological studies of the genus Onobrychis Mill. and the related genera Hedysarum L. and Sartoria Boiss. & Heldr (Fabaceae, Hedysareae) from Turkey. Caryologia 2012, 65, 11–17. [Google Scholar]
- Erdoğmuş, S.F.; Özkara, A.; Korcan, S.E.; Bağci, Y.; Dural, H. Sartoria hedysaroides Boiss. & Heldr. Ekstrelerinin Antimikrobiyal Aktivitesinin Belirlenmesi. Afyon Kocatepe Üniversitesi Fen Ve Mühendislik Bilimleri Derg. 2012, 12, 17–22. [Google Scholar]
- Huang, W.-J.; Zhang, X.; Chen, W.-W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Wright, E., Jr.; Scism-Bacon, J.L.; Glass, L.C. Oxidative stress in type 2 diabetes: The role of fasting and postprandial glycaemia. Int. J. Clin. Pract. 2006, 60, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Nahhas, A.F.; Abdel-Malek, Z.A.; Kohli, I.; Braunberger, T.L.; Lim, H.W.; Hamzavi, I.H. The potential role of antioxidants in mitigating skin hyperpigmentation resulting from ultraviolet and visible light-induced oxidative stress. Photodermatol. Photoimmunol. Photomed. 2019, 35, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef]
- Chong, J.; Yamamoto, M.; Xia, J. MetaboAnalystR 2.0: From raw spectra to biological insights. Metabolites 2019, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Perel’son, M.; Sheinker, Y.N.; Syrova, G.; Turchin, K. NMR spectra of natural coumarin derivatives. Chem. Nat. Compd. 1970, 6, 5–11. [Google Scholar] [CrossRef]
- Špirtović-Halilović, S.; Salihović, M.; Trifunović, S.; Roca, S.; Veljović, E.; Osmanović, A.; Vinković, M.; Završnik, D. Density functional theory: 1H and 13C-NMR spectra of some coumarin derivatives. J. Serb. Chem. Soc. 2014, 79, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef]
- Tsugawa, H.; Rai, A.; Saito, K.; Nakabayashi, R. Metabolomics and complementary techniques to investigate the plant phytochemical cosmos. Nat. Prod. Rep. 2021, 38, 1729–1759. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical Scheme for LC-MSn Identification of Chlorogenic Acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the Six Isomers of Dicaffeoylquinic Acid by LC-MSn. J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef] [PubMed]
- Kachlicki, P.; Piasecka, A.; Stobiecki, M.; Marczak, Ł. Structural Characterization of Flavonoid Glycoconjugates and Their Derivatives with Mass Spectrometric Techniques. Molecules 2016, 21, 1494. [Google Scholar] [CrossRef] [Green Version]
- Fabre, N.; Rustan, I.; de Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Ablajan, K.; Abliz, Z.; Shang, X.Y.; He, J.M.; Zhang, R.P.; Shi, J.G. Structural characterization of flavonol 3,7-di-O-glycosides and determination of the glycosylation position by using negative ion electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2006, 41, 352–360. [Google Scholar] [CrossRef]
- Cissé, L.; Alphonse, T.; Kaboré, L.; Adama, S. Mass Spectrometry Study of Coumarins: Correlation between Charges of Atoms and Fragmentation Processes. Spectrosc. Lett. Int. J. Rapid Commun. 2009, 42, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Liu, X.; Zhou, A.; Meng, M.; Li, Q. Simultaneous analysis of coumarin derivatives in extracts of Radix Angelicae pubescentis (Duhuo) by HPLC-DAD-ESI-MSn technique. Anal. Methods 2014, 6, 7996–8002. [Google Scholar] [CrossRef]
- Hung, T.M.; Na, M.; Thuong, P.T.; Su, N.D.; Sok, D.; Song, K.S.; Seong, Y.H.; Bae, K. Antioxidant activity of caffeoyl quinic acid derivatives from the roots of Dipsacus asper Wall. J. Ethnopharmacol. 2006, 108, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, L.; Xue, Q.; Yao, F.; Sun, J.; Yang, F.; Liu, Y. Antioxidant evaluation-guided chemical profiling and structure-activity analysis of leaf extracts from five trees in Broussonetia and Morus (Moraceae). Sci. Rep. 2020, 10, 4808. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Hammoudi, N.-E.-H.; Sobhi, W.; Attoui, A.; Lemaoui, T.; Erto, A.; Benguerba, Y. In silico drug discovery of Acetylcholinesterase and Butyrylcholinesterase enzymes inhibitors based on Quantitative Structure-Activity Relationship (QSAR) and drug-likeness evaluation. J. Mol. Struct. 2021, 1229, 129845. [Google Scholar] [CrossRef]
- Lima, J.A.; Costa, T.W.; da Fonseca, A.C.; do Amaral, R.F.; Maria do Desterro, S.B.; Santos-Filho, O.A.; de Miranda, A.L.P.; Ferreira Neto, D.C.; Lima, F.R.S.; Hamerski, L.; et al. A promising alkaloid for Alzheimer’s disease: Inhibition of human cholinesterases, anti-inflammatory effects and molecular docking. Bioorg. Chem. 2020, 104, 104215. [Google Scholar] [CrossRef]
- Chai, W.-M.; Yu, Z.-Y.; Lin, M.-Z.; Wei, Q.-M.; Song, S. 5-Methoxy-2-mercaptobenzimidazole as an efficient inhibitor on tyrosinase: Inhibitory activity and mechanism. J. Biosci. Bioeng. 2021, 131, 356–363. [Google Scholar] [CrossRef]
- Fallarero, A.; Oinonen, P.; Gupta, S.; Blom, P.; Galkin, A.; Mohan, C.G.; Vuorela, P.M. Inhibition of acetylcholinesterase by coumarins: The case of coumarin 106. Pharmacol. Res. 2008, 58, 215–221. [Google Scholar] [CrossRef]
- Anand, P.; Singh, B.; Singh, N. A review on coumarins as acetylcholinesterase inhibitors for Alzheimer’s disease. Bioorg. Med. Chem. 2012, 20, 1175–1180. [Google Scholar] [CrossRef]
- Amat-ur-Rasool, H.; Symes, F.; Tooth, D.; Schaffert, L.-N.; Elmorsy, E.; Ahmed, M.; Hasnain, S.; Carter, W.G. Potential nutraceutical properties of leaves from several commonly cultivated plants. Biomolecules 2020, 10, 1556. [Google Scholar] [CrossRef]
- Grzelczyk, J.; Budryn, G. Analysis of the activity of hydroxycinnamic acids from green and roasted coffee extracts as acetylcholinesterase inhibitors using an isothermal method of titration calorimetry. Folia Pomeranae Univ. Technol. Stetin. Agric. Aliment. Piscaria Zootech. 2019, 349, 15–23. [Google Scholar] [CrossRef]
- Budryn, G.; Grzelczyk, J.; Jaśkiewicz, A.; Żyżelewicz, D.; Pérez-Sánchez, H.; Cerón-Carrasco, J.P. Evaluation of butyrylcholinesterase inhibitory activity by chlorogenic acids and coffee extracts assed in ITC and docking simulation models. Food Res. Int. 2018, 109, 268–277. [Google Scholar] [CrossRef]
- Takao, K.; Toda, K.; Saito, T.; Sugita, Y. Synthesis of Amide and Ester Derivatives of Cinnamic Acid and Its Analogs: Evaluation of Their Free Radical Scavenging and Monoamine Oxidase and Cholinesterase Inhibitory Activities. Chem. Pharm. Bull. 2017, 65, 1020–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.; Sousa, J.L.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzym. Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Park, J.Y.; Lee, S.; Kang, K.S. In Vitro Studies to Assess the α-Glucosidase Inhibitory Activity and Insulin Secretion Effect of Isorhamnetin 3-O-Glucoside and Quercetin 3-O-Glucoside Isolated from Salicornia herbacea. Processes 2021, 9, 483. [Google Scholar] [CrossRef]
- Davò, F.; Alessandrini, S.; Sperati, S.; Delle Monache, L.; Airoldi, D.; Vespucci, M.T. Post-processing techniques and principal component analysis for regional wind power and solar irradiance forecasting. Sol. Energy 2016, 134, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Piluzza, G.; Bullitta, S. Correlations between phenolic content and antioxidant properties in twenty-four plant species of traditional ethnoveterinary use in the Mediterranean area. Pharm. Biol. 2011, 49, 240–247. [Google Scholar] [CrossRef] [PubMed]







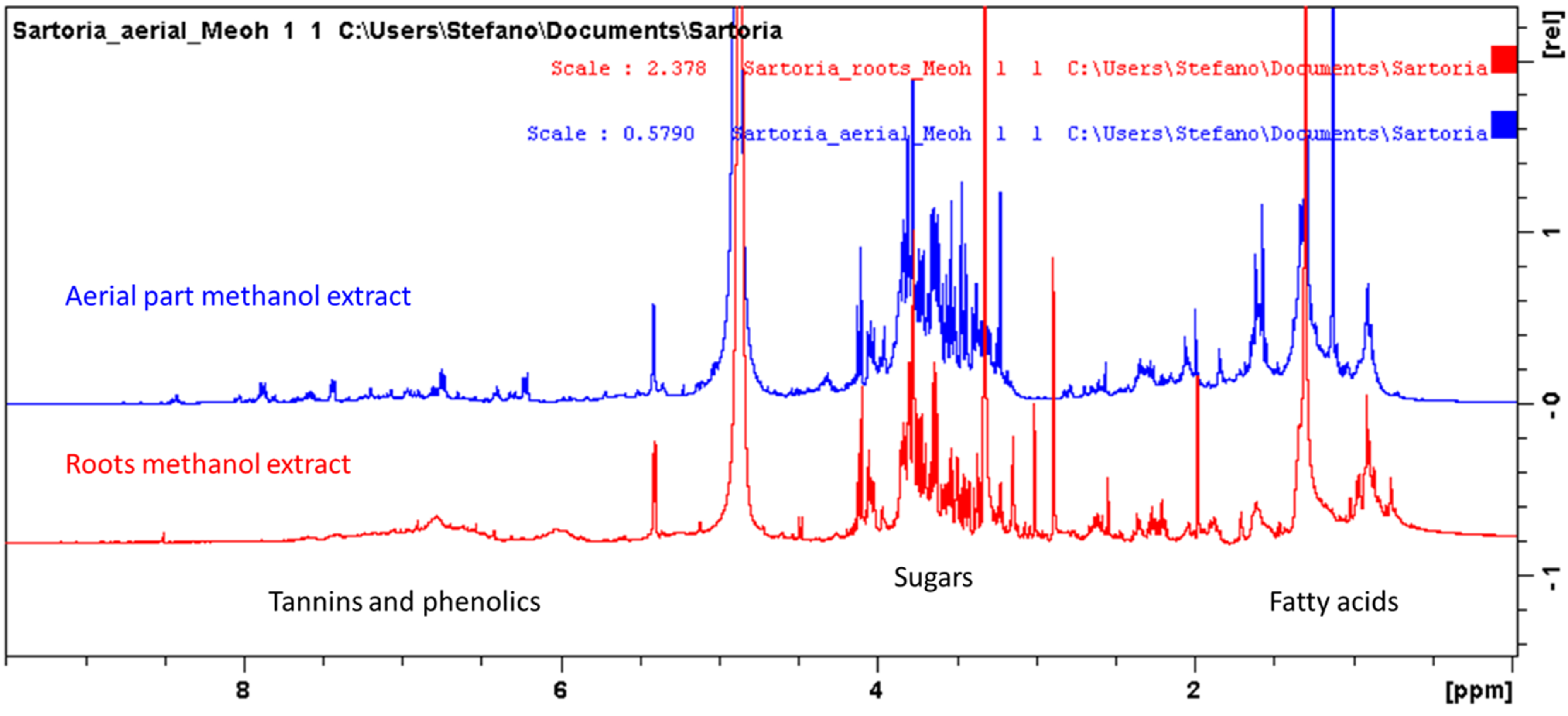{kind=link}
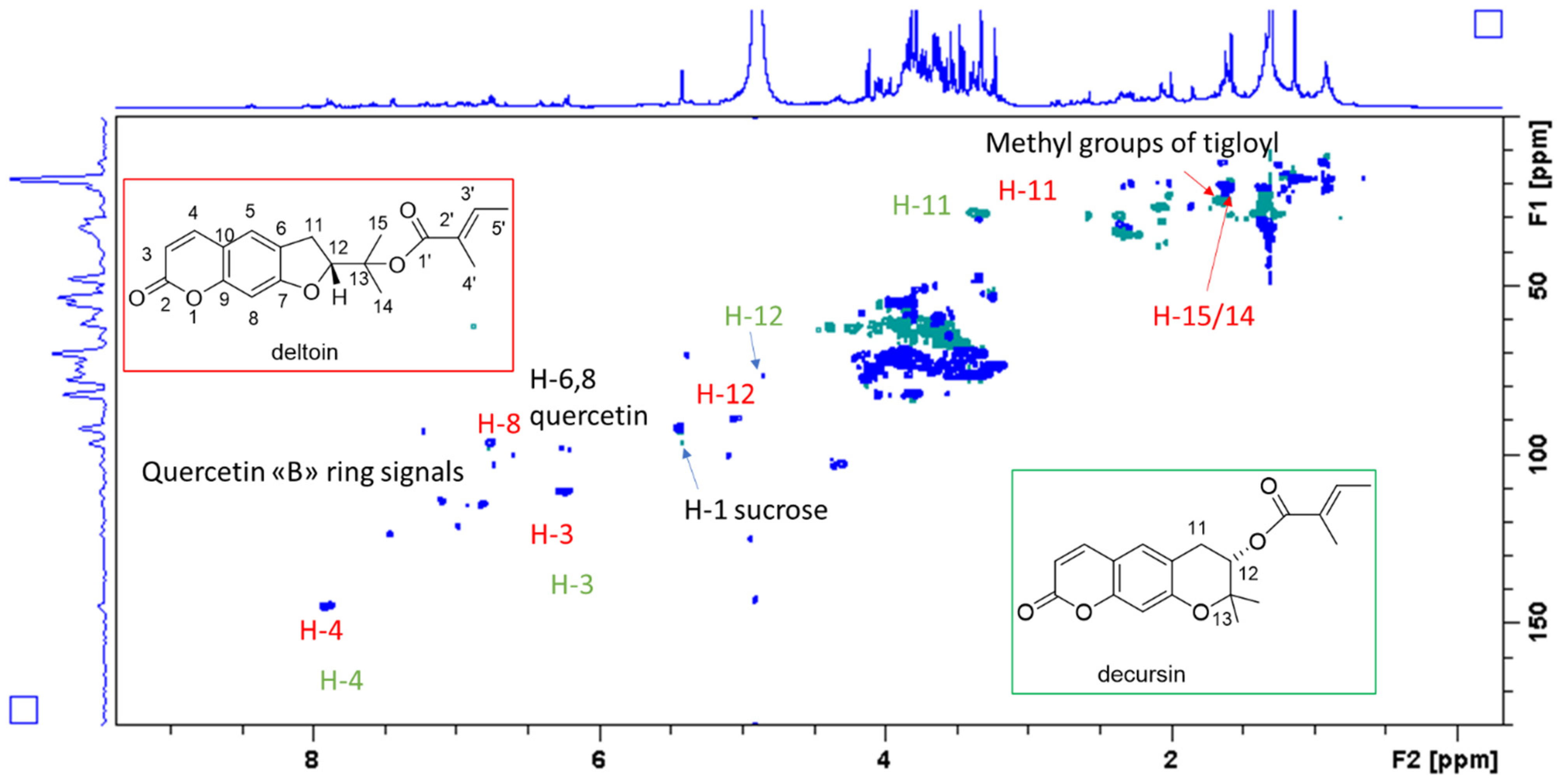{kind=link}
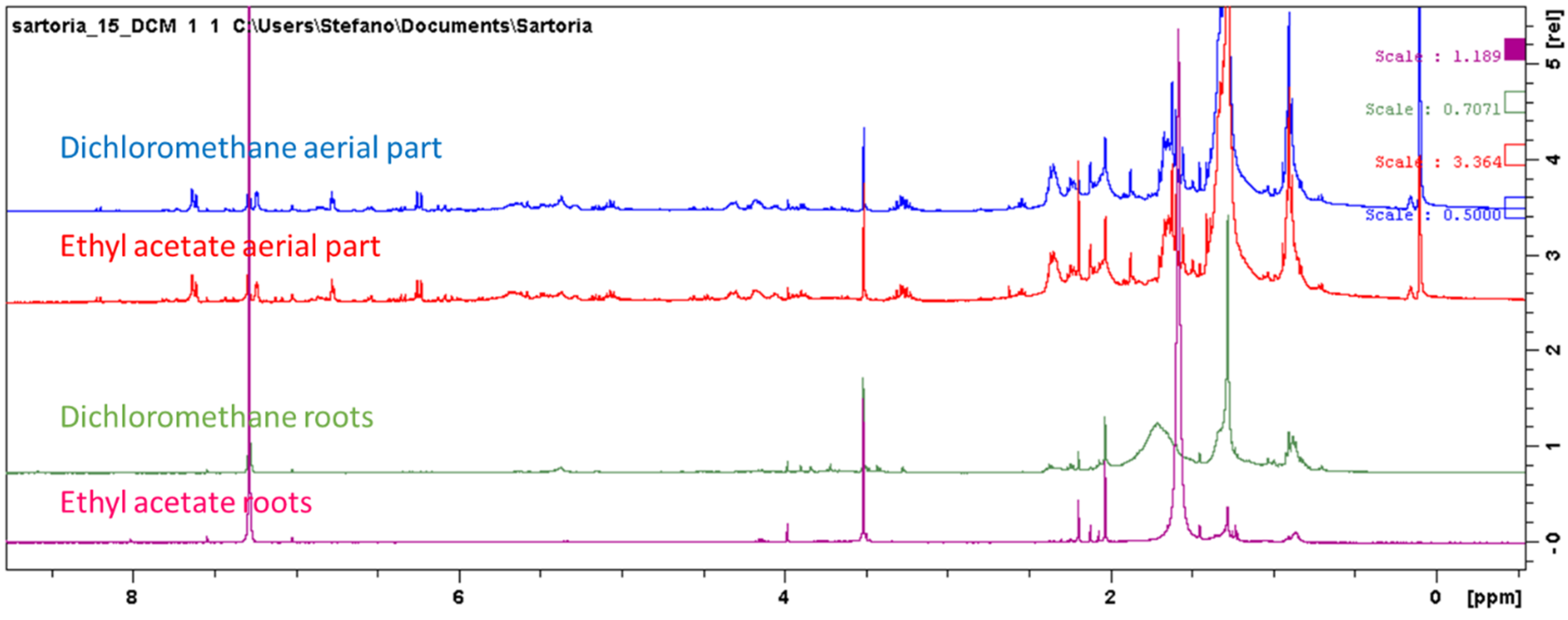{kind=link}
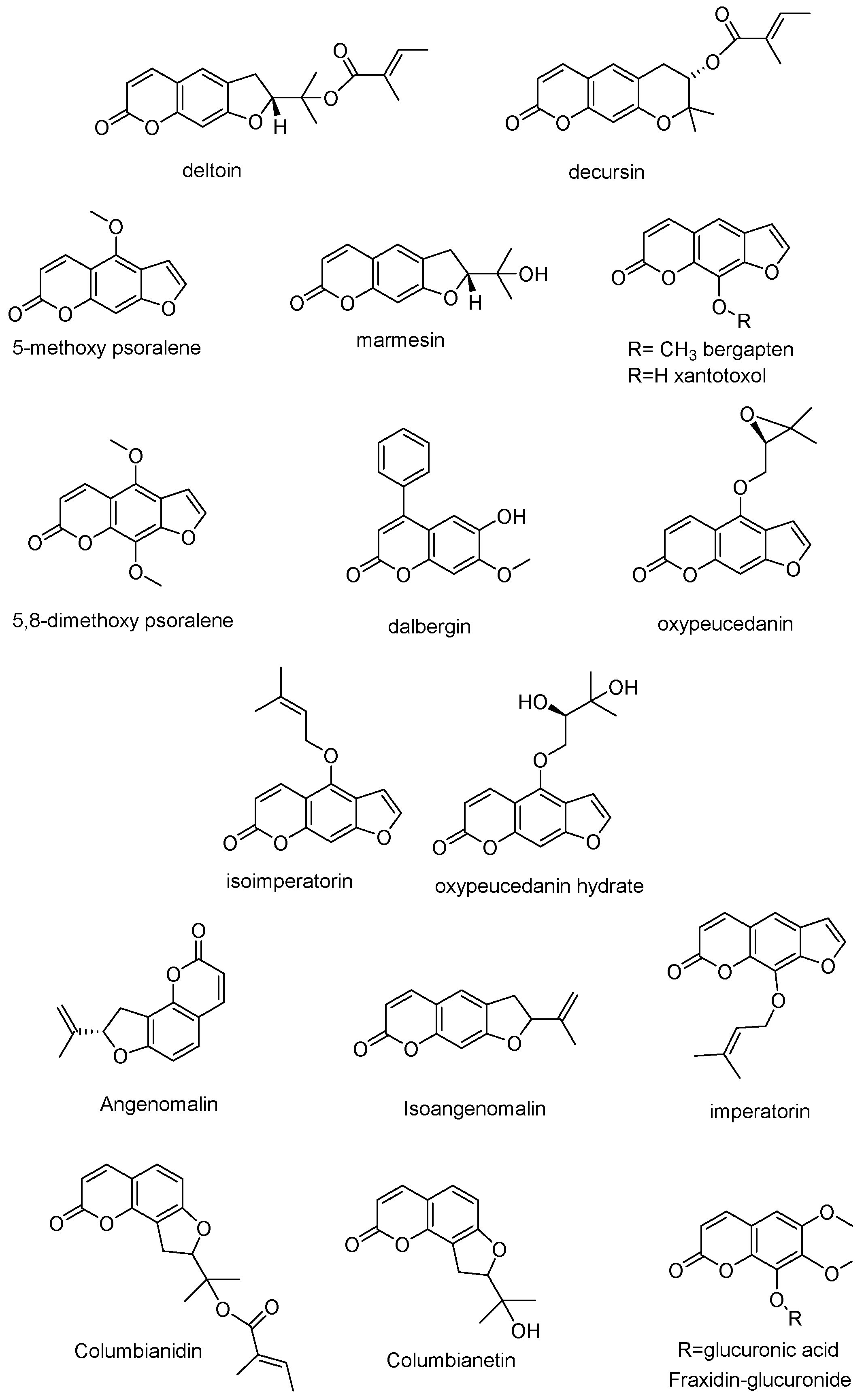{kind=link}
{kind=link}
{kind=link}
| Solvents | TPC (mg GAE/g) * | TFC (mg RE/g) * | |
|---|---|---|---|
| Aerial parts | DCM | 24.70 ± 0.59 e | 8.30 ± 0.55 e |
| EA | 29.17 ± 0.10 c | 8.33 ± 0.40 e | |
| MeOH | 67.64 ± 0.63 a | 92.03 ± 0.43 a | |
| Infusion | 45.33 ± 0.21 b | 59.54 ± 0.80 b | |
| Roots | DCM | 22.17 ± 0.06 f | 15.06 ± 0.54 d |
| EA | 29.82 ± 0.36 c | 16.91 ± 0.24 c | |
| MeOH | 27.62 ± 0.17 d | 16.23 ± 0.09 cd | |
| Infusion | 24.56 ± 0.14 e | 1.27 ± 0.22 f |
| Class of Compounds | δH | δC | HMBC | COSY/NOESY |
|---|---|---|---|---|
| Deltoin and decursin | ||||
| 2 | - | 162 | - | - |
| 3 | 6.23 | 111.0 | 163 112.6 | 7.85 H-4 COSY |
| 4 | 7.85 | 144.0 | 163 154 123 | 6.20 H-3 COSY |
| 5 | 7.44 | 123.0 | 144 163 154 | - |
| 6 | - | 127 | - | - |
| 7 | - | 164 | - | - |
| 8 | 6.75 | 96.0 | 163 154 127 113 | - |
| 9 | - | 154 | - | - |
| 10 | - | 113 | - | - |
| 11 Deltoin | 3.36–3.30 | 28.7 | 127 89 82 | 4.90 COSY |
| 12 Deltoin | 4.90 (under methanol) | 89 | 3.30–3.36 COSY | |
| 11 Decursin | 3.40 | 28.0 | 127 89 82 | 4.87 COSY |
| 12 Decursin | 4.86 (under methanol) | 77 | 3.30–3.36 COSY | |
| 14 | 1.59 | 20.9 | 82 89 20 | |
| 15 | 1.63 | 20.5 | 82 89 20 | |
| Tigloyl moiety of deltoin and decursin | ||||
| 1′ | - | 167 | - | - |
| 2′ | - | 130 | - | - |
| 3′ | 6.40 | 137 | 167 137 130 | |
| 4′ | 1.60 | 20 superimposed | 167 137 130 | |
| 5′ | 1.64 | 20 superimposed | 137 130 | |
| Furocumarins | ||||
| 11 | 6.68–6.60 | 106–104 | - | - |
| 12 | 7.60–7.65 | 145 | 158 116 | - |
| Flavonols | ||||
| 6, 8 | 6.40 6.30 6.19 | 99.1 96.5 92.5 | ||
| 2′ | 7.68 | 126.5 | ||
| 5′ | 6.73 | 115 | ||
| 6′ | 7.25 | 114.5 | ||
| Sucrose | ||||
| 1 | 5.30 | 92.8 | - | - |
| 2 | 3.55 | 76.4 | ||
| 6 | 3.82 | 62.6 | - | - |
| 1′ | 3.67 | 76.4 | - | - |
| 3′ | 4.21 d | 77.4 | - | - |
| Aerial | Roots | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | Formula (Neutral Compound) | Theorical m/z * | Exp m/z * | ppm | DCM | EA | MeOH | W | DCM | EA | MeOH | W |
| Phenolics and Organic acid | ||||||||||||
| Quinic acid | C7H12O6 | 191.0555 | 191.0565 | −5.55 | - | 0.10 | 2.16 | 0.25 | - | 0.06 | 5.09 | 0.19 |
| Caffeoyl glucose | C15H18O9 | 341.0872 | 341.0887 | −4.66 | - | - | 0.09 | - | - | 0.03 | 0.01 | 0.03 |
| 3-p-Coumaroyl quinic acid | C13H16O9 | 337.0923 | 337.0937 | −4.40 | - | - | 0.12 | - | - | 0.03 | 0.02 | 0.08 |
| Protocatecuic acid hexoside | C16H18O9 | 315.0716 | 315.0731 | −5.05 | - | - | 0.20 | - | - | 0.04 | 0.17 | 0.30 |
| p-Methoxy coumaroyl quinic acid | C17H20O8 | 351.1079 | 351.1095 | −4.83 | - | - | 0.46 | - | - | 0.14 | 0.02 | 0.04 |
| Quercetin-3-O-rhamnoside | C21H20O11 | 447.0927 | 447.0939 | −2.84 | - | - | 5.00 | - | - | 1.50 | 1.93 | 2.81 |
| 3-Caffeoyl quinic acid | C16H18O9 | 353.0872 | 353.0888 | −4.80 | - | - | 4.47 | 0.96 | - | 1.34 | 0.52 | 0.21 |
| 5-p-Coumaroyl quinic acid | C13H16O9 | 337.0923 | 337.0940 | −5.35 | - | - | 0.40 | 7.75 | - | 0.12 | 0.12 | 0.54 |
| Rutin | C27H30O16 | 609.1455 | 609.1456 | −0.17 | 0.04 | 0.05 | 3.93 | 11.93 | 0.39 | 0.28 | 0.08 | 0.45 |
| 4-Feruloyl quinic acid | C17H20O9 | 367.1029 | 367.1038 | −2.60 | 0.05 | 0.06 | 5.97 | 17.08 | 0.19 | 0.67 | 0.61 | 0.83 |
| 3,4-Dicaffeoyl quinic acid | C25H24O12 | 515.1189 | 515.1201 | −2.47 | - | 0.01 | 4.85 | 1.69 | - | 1.46 | 1.75 | 0.45 |
| Quercetin-3-O-glucuronide | C21H18O13 | 477.0669 | 477.0687 | −4.00 | - | 0.01 | 0.75 | 2.36 | - | 0.22 | 0.46 | 0.12 |
| Isorhamnetin 3-O-rhamnoglucoside | C28H32O16 | 623.1612 | 623.1614 | −0.34 | - | 0.07 | 0.74 | 5.20 | 0.22 | 0.22 | 0.14 | 0.19 |
| Demethyl hedysarimpterocarpene-A rutinoside | C28H32O15 | 607.1663 | 607.1661 | 0.35 | - | 0.01 | 8.25 | 1.46 | 0.03 | 2.48 | 3.06 | 0.14 |
| Taxifolin hexoside | C21H22O12 | 465.1033 | 465.1044 | −2.51 | - | 0.07 | 2.18 | 0.22 | 0.21 | 0.65 | 0.62 | 0.15 |
| 3,5-Dicaffeoyl quinic acid | C25H24O12 | 515.1189 | 515.1191 | −0.41 | - | 0.05 | 0.26 | 2.54 | 0.15 | 0.08 | 0.02 | 0.16 |
| 4,5-Dicaffeoyl quinic acid | C25H24O12 | 515.1189 | 515.1191 | −0.41 | - | 0.08 | 0.48 | 4.37 | 1.74 | 0.14 | 0.06 | 0.16 |
| Taxifolin | C15H12O7 | 303.0504 | 303.0521 | −5.95 | 0.23 | 0.17 | 0.26 | 1.40 | 5.02 | 0.97 | 0.03 | 0.02 |
| 3-Caffeoyl-4-feruloyl quinic acid | C26H26O12 | 529.1346 | 529.1369 | −4.61 | 0.12 | 0.20 | 0.48 | 0.74 | 1.58 | 0.04 | 0.02 | 0.04 |
| Isorhamnetin derivative | - | - | 777.5536 | - | 0.14 | 0.14 | 0.24 | 0.56 | 1.16 | 0.05 | 0.05 | 0.02 |
| p-Hydroxy-benzoic acid | C7H6O3 | 137.0238 | 137.0245 | −5.42 | 0.05 | 0.39 | 0.12 | 0.17 | 1.16 | 0.04 | 0.02 | 0.02 |
| Vanilic acid glucoside | C14H18O9 | 329.0872 | 329.0885 | −4.20 | 0.02 | 0.08 | 0.10 | 0.11 | 1.52 | 0.03 | 0.02 | 0.02 |
| total | 0.65 | 1.48 | 41.52 | 58.82 | 13.37 | 10.57 | 14.82 | 6.97 | ||||
| Aerial | Roots | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tannins | Formula (Neutral Compound) | Theorical m/z * | Exp m/z * | ppm | DCM | EA | MeOH | W | DCM | EA | MeOH | W |
| Ellagic acid hexoside | C20H16O13 | 465.0669 | 465.0681 | −2.74 | - | - | - | - | 3.72 | 3.59 | 0.01 | 1.80 |
| Ellagic acid hexoside | C20H16O13 | 465.0669 | 465.0683 | −3.19 | - | - | 0.85 | - | 3.59 | 2.28 | 2.56 | 2.42 |
| Ellagic acid hexoside | C20H16O13 | 465.0669 | 465.0681 | −2.74 | - | - | - | 0.35 | 22.63 | 0.92 | 0.01 | 0.47 |
| Ellagic acid deoxyhexoside | C20H16O12 | 449.0720 | 449.0732 | −2.83 | - | - | 0.59 | - | 2.91 | 2.84 | 1.76 | 2.30 |
| Ellagic acid deoxyhexoside | C20H16O13 | 449.0721 | 449.0732 | −2.70 | - | - | - | - | 1.49 | 1.30 | 0.02 | 0.66 |
| Ellagic acid deoxyhexoside | C20H16O14 | 449.0720 | 449.0732 | −2.80 | - | - | - | - | 9.46 | 8.55 | 0.01 | 4.28 |
| Ellagic acid derivative | C20H13O14 | 477.0295 | 477.0305 | −2.22 | - | - | 0.29 | 0.64 | 3.61 | 3.26 | 54.86 | 29.06 |
| Ellagic acid derivative | C20H13O14 | 477.0295 | 477.0305 | −2.22 | - | - | 4.27 | 4.44 | 1.61 | 1.48 | 12.82 | 7.15 |
| Ellagic acid derivative | C20H13O14 | 477.0295 | 477.0305 | −2.22 | - | - | 2.11 | 1.87 | 2.31 | 2.11 | 6.32 | 4.21 |
| Ellagic acid | C14H6O8 | 300.9983 | 300.9985 | −0.56 | - | - | 5.42 | 3.74 | 11.21 | 10.16 | 16.25 | 13.21 |
| total | - | - | 13.53 | 11.03 | 62.55 | 36.49 | 94.63 | 65.56 | ||||
| Aerial | Roots | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Formula (Neutral Compound) | Theorical m/z * | Exp m/z * | ppm | DCM | EA | MeOH | W | DCM | EA | MeOH | W | |
| Fraxidin glucuronide § | C16H16O11 | 383.0609 | 383.0614 | −1.39 | 0.004 | 0.01 | 1.52 | 5.66 | - | 0.46 | 0.46 | 0.23 |
| Columbianetin | C14H14O4 | 247.0970 | 247.0957 | 5.62 | 0.05 | 0.23 | 0.28 | 0.03 | - | - | - | - |
| Columbianidin | C19H20O5 | 329.1389 | 329.1407 | −5.80 | 2.46 | 0.31 | 1.05 | 0.39 | 0.02 | 0.01 | 0.09 | 0.03 |
| Dalbergin isomer | C16H12O4 | 269.0814 | 269.0830 | −6.30 | 1.49 | 0.18 | 0.66 | 0.23 | - | - | - | - |
| Marmesin | C14H14O4 | 247.0970 | 247.0957 | 5.62 | 2.11 | 0.26 | 0.98 | 0.33 | - | - | - | - |
| Oxypeucedanin hydrate | C16H16O6 | 305.1025 | 305.1039 | −4.86 | 0.54 | 0.06 | 0.27 | 0.08 | - | - | - | - |
| Isoimperatorin | C16H14O4 | 271.0970 | 271.0974 | −1.56 | 1.23 | 0.15 | 0.59 | 0.18 | - | - | - | - |
| 8-Methoxypsoralene | C12H8O4 | 217.0501 | 217.0521 | −9.77 | 13.19 | 1.64 | 6.32 | 2.08 | - | - | - | - |
| Bergapten | C12H8O4 | 217.0501 | 217.0521 | −9.77 | 4.07 | 0.46 | 1.68 | 0.58 | 0.07 | - | - | - |
| Isopimpinellin 5,8-dimethoxypsoralene | C13H10O5 | 247.0610 | 247.0630 | −8.58 | 3.79 | 0.45 | 1.62 | 0.57 | 0.04 | - | - | - |
| Dalbergin | C16H12O4 | 269.0814 | 269.0830 | −6.30 | 1.31 | 0.15 | 0.53 | 0.19 | 0.09 | - | - | - |
| Angenomalin/isoangenomalin | C14H12O3 | 229.0845 | 229.0855 | −4.63 | 4.12 | 0.41 | 1.52 | 0.52 | - | - | - | - |
| Oxypeucedanin | C16H14O5 | 287.0920 | 287.0943 | −8.49 | 2.96 | 0.28 | 1.03 | 0.36 | - | - | - | - |
| Xantotoxol | C11H6O4 | 203.0344 | 203.0360 | −8.35 | 59.12 | 11.45 | 43.74 | 14.48 | - | - | - | - |
| Deltoin | C19H20O5 | 329.1389 | 329.1389 | 0 | 2.02 | 0.17 | 0.66 | 0.22 | - | - | - | - |
| Decursin | C19H20O5 | 329.1389 | 329.1404 | −4.83 | 30.53 | 9.29 | 41.31 | 11.75 | - | - | - | - |
| Angelomalin/isoangelomalin | C14H12O3 | 229.0865 | 229.0885 | −9.25 | 3.77 | 0.36 | 1.30 | 0.45 | - | - | - | - |
| Total | 132.77 | 25.86 | 105.04 | 38.09 | 0.22 | 0.47 | 0.56 | 0.26 | ||||
| Solvents | DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | MCA (mg EDTAE/g) | PBD (mmol TE/g) | |
|---|---|---|---|---|---|---|---|
| Aerial parts | DCM | 4.93 ± 0.28 h | 31.75 ± 1.31 f | 68.71 ± 1.28 e | 23.24 ± 0.42 f | 15.77 ± 0.21 c,d | 1.82 ± 0.08 a,b |
| EA | 12.46 ± 0.42 g | 47.99 ± 0.47 e | 92.47 ± 0.82 c | 33.04 ± 0.49 e | 17.60 ± 0.52 c | 2.08 ± 0.11 a | |
| MeOH | 169.97 ± 1.32 a | 285.33 ± 11.94 a | 279.23 ± 0.20 a | 162.48 ± 2.52 a | 11.87 ± 0.36 d,e | 1.87 ± 0.13 a,b | |
| Infusion | 74.32 ± 1.30 b | 164.37 ± 1.62 b | 171.80 ± 1.63 b | 121.87 ± 3.70 b | 23.44 ± 0.21 a,b | 1.08 ± 0.01 d | |
| Roots | DCM | 25.50 ± 0.25 e | 54.06 ± 1.76 e | 67.67 ± 0.73 e | 36.11 ± 0.44 e | 18.99 ± 4.67 b,c | 1.68 ± 0.04 b,c |
| EA | 30.72 ± 0.20 d | 71.03 ± 0.79 d | 92.58 ± 1.23 c | 44.63 ± 0.15 d | 9.76 ± 1.57 e | 1.73 ± 0.13 b | |
| MeOH | 37.69 ± 0.30 c | 99.65 ± 0.38 c | 88.63 ± 0.99 d | 54.68 ± 1.55 c | 11.76 ± 0.36 d,e | 1.40 ± 0.19 c | |
| Infusion | 14.79 ± 0.08 f | 80.26 ± 1.06 d | 59.09 ± 1.65 f | 41.41 ± 0.24 d | 26.67 ± 0.21 a | 1.07 ± 0.04 d |
| Solvents | AchE İnhibition (mg GALAE/g) | BchE İnhibition (mg GALAE/g) | Tyrosinase İnhibition (mg KAE/g) | Amylase İnhibition (mmol ACAE/g) | Glucosidase İnhibition (mmol ACAE/g) | |
|---|---|---|---|---|---|---|
| Aerial parts | DCM | 0.68 ± 0.08 d | 5.96 ± 0.71 a | 56.71 ± 0.34 d | 0.95 ± 0.01 b,c | 0.79 ± 0.03 d |
| EA | 1.29 ± 0.03 c | 5.45 ± 1.07 a | 57.85 ± 0.08 d | 1.07 ± 0.06 a | 0.83 ± 0.02 c,d | |
| MeOH | 2.52 ± 0.02 a | 2.75 ± 0.24 b | 74.14 ± 0.19 a | 1.07 ± 0.04 a | 1.03 ± 0.01 a | |
| Infusion | 0.54 ± 0.02 e | na | 7.02 ± 0.74 e | 0.10 ± 0.01 e | 1.02 ± 0.01 a | |
| Roots | DCM | na | 6.03 ± 0.51 a | 56.74 ± 0.07 d | 1.04 ± 0.06 a,b | 0.90 ± 0.01 b |
| EA | 1.79 ± 0.09 b | 5.14 ± 0.38 a | 59.48 ± 1.27 c | 1.08 ± 0.04 a | 0.87 ± 0.01 b,c | |
| MeOH | 1.73 ± 0.05 b | 3.39 ± 0.21 b | 64.99 ± 0.12 b | 0.87 ± 0.05 c | 0.87 ± 0.04 b,c | |
| Infusion | na | na | 1.65 ± 0.03 f | 0.26 ± 0.01 d | 0.02 ± 0.01 e |
| DPPH | ABTS | CUPRAC | FRAP | MCA | PPBD | AChE | BChE | Tyrosinase | Amylase | Glucosidase | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TPC | 0.97 | 0.96 | 0.99 | 0.96 | −0.21 | 0.08 | 0.62 | −0.36 | 0.16 | −0.02 | 0.45 |
| TFC | 0.97 | 0.95 | 0.98 | 0.98 | −0.19 | 0.01 | 0.52 | −0.36 | 0.13 | −0.06 | 0.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dall’Acqua, S.; Sut, S.; Sinan, K.I.; Zengin, G.; Ferrarese, I.; Peron, G.; Yildiztugay, E.; Picot-Allain, C.; Mahomoodally, M.F. An Integrated NMR, LC-DAD-MS, LC-QTOF Metabolomic Characterization of Sartoria hedysaroides: Correlation of Antioxidant and Enzyme Inhibitory Activity with Chemical Composition by Multivariate Data Analysis. Antioxidants 2022, 11, 110. https://doi.org/10.3390/antiox11010110
Dall’Acqua S, Sut S, Sinan KI, Zengin G, Ferrarese I, Peron G, Yildiztugay E, Picot-Allain C, Mahomoodally MF. An Integrated NMR, LC-DAD-MS, LC-QTOF Metabolomic Characterization of Sartoria hedysaroides: Correlation of Antioxidant and Enzyme Inhibitory Activity with Chemical Composition by Multivariate Data Analysis. Antioxidants. 2022; 11(1):110. https://doi.org/10.3390/antiox11010110
Chicago/Turabian StyleDall’Acqua, Stefano, Stefania Sut, Kouadio Ibrahime Sinan, Gokhan Zengin, Irene Ferrarese, Gregorio Peron, Evren Yildiztugay, Carene Picot-Allain, and Mohamad Fawzi Mahomoodally. 2022. "An Integrated NMR, LC-DAD-MS, LC-QTOF Metabolomic Characterization of Sartoria hedysaroides: Correlation of Antioxidant and Enzyme Inhibitory Activity with Chemical Composition by Multivariate Data Analysis" Antioxidants 11, no. 1: 110. https://doi.org/10.3390/antiox11010110
APA StyleDall’Acqua, S., Sut, S., Sinan, K. I., Zengin, G., Ferrarese, I., Peron, G., Yildiztugay, E., Picot-Allain, C., & Mahomoodally, M. F. (2022). An Integrated NMR, LC-DAD-MS, LC-QTOF Metabolomic Characterization of Sartoria hedysaroides: Correlation of Antioxidant and Enzyme Inhibitory Activity with Chemical Composition by Multivariate Data Analysis. Antioxidants, 11(1), 110. https://doi.org/10.3390/antiox11010110