
Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
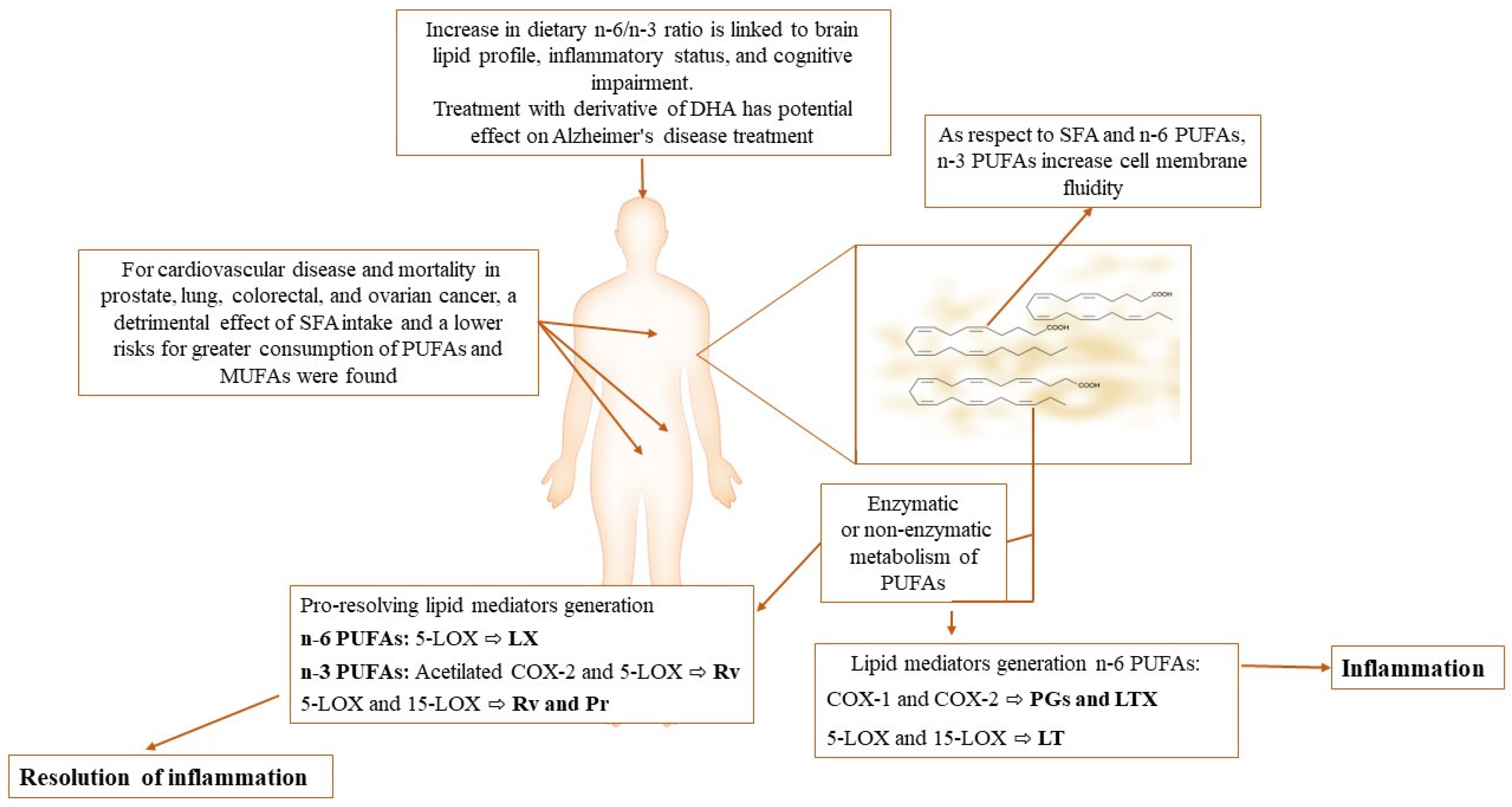
:1. Introduction
2. Fatty Acids in Normal and Altered Spermatogenesis
3. Influence of Dietary FA Supplementation on Sperm Quality and Function
4. Current New Indices of Male Infertility Involving PUFA Oxidation
4.1. F2-Isoprostanes
4.2. What Is the Role of Resolvins?
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Escribá, P.V.; Nicolson, G.L. Membrane structure and function: Relevance of lipid and protein structures in cellular physiology, pathology and therapy. Biochim. Biophys. Acta 2014, 1838, 1449–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichler, H.; Emmerstorfer-Augustin, A. Modification of membrane lipid compositions in single-celled organisms-From basics to applications. Methods 2018, 147, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Devkota, R.; Kaper, D.; Bodhicharla, R.; Henricsson, M.; Borén, J.; Pilon, M. A genetic titration of membrane composition in Caenorhabditis elegans reveals its importance for multiple cellular and physiological traits. Genetics 2021, 219, iyab093. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Song, C. Potential treatment of Parkinson’s disease with omega-3 polyunsaturated fatty acids. Nutr. Neurosci. 2022, 25, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Suburu, J.; Gu, Z.; Chen, H.; Chen, W.; Zhang, H.; Chen, Y.Q. Fatty acid metabolism: Implications for diet, genetic variation, and disease. Food Biosci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Custers, E.M.; Kiliaan, A.J.; Amanda, J. Dietary lipids from body to brain. Prog. Lipid Res. 2022, 85, 101144. [Google Scholar] [CrossRef]
- Brown, D.A. Lipid Rafts. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 741–744. [Google Scholar]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The imbalance between n-6/n-3 polyunsaturated fatty acids and inflammatory bowel disease: A comprehensive review and future therapeutic perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef] [Green Version]
- Kotlyarov, S.; Kotlyarova, A. Anti-Inflammatory function of fatty acids and involvement of their metabolites in the resolution of inflammation in chronic obstructive pulmonary disease. Int. J. Mol. Sci. 2021, 22, 12803. [Google Scholar] [CrossRef]
- Mani, V.; Hollis, J.H.; Gabler, N.K. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Nutr. Metab. 2013, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.R.W.; et al. Evidence that TLR4 is not a receptor for saturated fatty acids but mediates lipid-induced inflammation by reprogramming macrophage metabolism. Cell Metab. 2018, 27, 1096–1110.e5. [Google Scholar] [CrossRef] [Green Version]
- Mallick, R.; Basak, S.; Duttaroy, A.K. Docosahexaenoic acid, 22:6n-3: Its roles in the structure and function of the brain. Int. J. Dev. Neurosci. 2019, 79, 21–31. [Google Scholar] [CrossRef] [PubMed]
- von Schacky, C. Importance of EPA and DHA blood levels in brain structure and function. Nutrients 2021, 13, 1074. [Google Scholar] [CrossRef]
- Guo, X.F.; Li, K.L.; Li, J.M.; Li, D. Effects of EPA and DHA on blood pressure and inflammatory factors: A meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2019, 59, 3380–3393. [Google Scholar] [CrossRef]
- Serhan, C.N.; Petasis, N.A. Resolvins and protectins in inflammation resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory links between high fat diets and diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothapalli, K.S.D.; Park, H.G.; Brenna, J.T. Polyunsaturated fatty acid biosynthesis pathway and genetics. implications for interindividual variability in prothrombotic, inflammatory conditions such as COVID-19. Prostaglandins Leukot. Essent. Fat. Acids 2020, 162, 102183. [Google Scholar] [CrossRef]
- Sala, A.; Proschak, E.; Steinhilber, D.; Rovati, G.E. Two-pronged approach to anti-inflammatory therapy through the modulation of the arachidonic acid cascade. Biochem. Pharmacol. 2018, 158, 161–173. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Ibarguren, M.; López, D.J.; Escribá, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta 2014, 1838, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.; Price, S.L.; Fiol-Deroque, M.A.; Marcilla-Etxenike, A.; Ahyayauch, H.; Barceló-Coblijn, G.; Terés, S.; Katsouri, L.; Ordinas, M.; López, D.J.; et al. Membrane lipid modifications and therapeutic effects mediated by hydroxydocosahexaenoic acid on Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1838, 1680–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordóñez-Gutiérrez, L.; Fábrias, G.; Casas, J.; Wandosell, F. Diets with higher ω-6/ω-3 ratios show differences in ceramides and fatty acid levels accompanied by increased amyloid-beta in the brains of male APP/PS1 transgenic mice. Int. J. Mol. Sci. 2021, 22, 10907. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Xu, X.; Wang, S.; Xia, D. Associations of dietary fat intake with mortality from all causes, cardiovascular disease, and cancer: A prospective study. Front. Nutr. 2021, 8, 701430. [Google Scholar] [CrossRef] [PubMed]
- Nienaber, A.; Ozturk, M.; Dolman, R.; Blaauw, R.; Zandberg, L.L.; van Rensburg, S.; Britz, M.; Hayford, F.E.A.; Brombacher, F.; Loots, D.T.; et al. n-3 long-chain PUFA promote antibacterial and inflammation-resolving effects in Mycobacterium tuberculosis-infected C3HeB/FeJ mice, dependent on fatty acid status. Br. J. Nutr. 2022, 127, 384–397. [Google Scholar] [CrossRef]
- Furland, N.E.; Maldonado, E.N.; Ayuza Aresti, P.; Aveldaño., M.I. Changes in lipids containing long- and very long-chain polyunsaturated fatty acids in cryptorchid rat testes. Biol. Reprod. 2007, 77, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Hurtado de Catalfo, G.E.; de Gómez Dumm, I.N.T. Influence of testosterone on polyunsaturated fatty acid biosynthesis in Sertoli cells in culture. Cell Biochem. Funct. 2005, 23, 175–180. [Google Scholar] [CrossRef]
- Gautier, C.; Scarlet, D.; Ertl, R.; Walter, I.; Wulf, M.; Nagel, C.; Aurich, J.; Aurich, C. Expression of enzymes involved in polyunsaturated fatty acid synthesis in the stallion testis and epididymis. Reprod. Fertil. Dev. 2020, 32, 851–861. [Google Scholar] [CrossRef]
- Castellini, C.; Mattioli, S.; Moretti, E.; Cotozzolo, E.; Perini, F.; Dal Bosco, A.; Signorini, C.; Noto, D.; Belmonte, G.; Lasagna, E.; et al. Expression of genes and localization of enzymes involved in polyunsaturated fatty acid synthesis in rabbit testis and epididymis. Sci. Rep. 2022, 12, 2637. [Google Scholar] [CrossRef]
- Hu, X.; Ge, X.; Liang, W.; Shao, Y.; Jing, J.; Wang, C.; Zeng, R.; Yao, B. Effects of saturated palmitic acid and omega-3 polyunsaturated fatty acids on Sertoli cell apoptosis. Syst. Biol. Reprod. Med. 2018, 64, 368–380. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Kim, Y.J.; Kang, B.H.; Yun, Y.S.; Park, C.K. The relationship between acrosome reaction and polyunsaturated fatty acid composition in boar sperm. Reprod. Domest. Anim. 2020, 55, 624–631. [Google Scholar] [CrossRef]
- Lenzi, A.; Gandini, L.; Maresca, V.; Rago, R.; Sgrò, P.; Dondero, F.; Picardo, M. Fatty acid composition of spermatozoa and immature germ cells. Mol. Hum. Reprod. 2000, 6, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbinati, C.; Caponecchia, L.; Rago, R.; Leoncini, E.; Bottaccioli, A.G.; Ciacciarelli, M.; Pacelli, A.; Salacone, P.; Sebastianelli, A.; Pastore, A.; et al. Fatty acids profiling reveals potential candidate markers of semen quality. Andrology 2016, 4, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, J.C.; Landeras, J.; Gadea, J. Spermatozoa and seminal plasma fatty acids as predictors of cryopreservation success. Andrology 2013, 1, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksoy, Y.; Aksoy, H.; Altinkaynak, K.; Aydin, H.R.; Ozkan, A. Sperm fatty acid composition in subfertile men. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Tavilani, H.; Doosti, M.; Abdi, K.; Vaisiraygani, A.; Joshaghani, H.R. Decreased polyunsaturated and increased saturated fatty acid concentration in spermatozoa from asthenozoospermic males as compared with normozoospermic males. Andrologia 2006, 38, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.X.; Yuan, D.J.; Wang, Q.L.; Jiang, F.; Guo, J.; Tang, Y.G.; Zheng, L.X.; Kang, J.X. Association of decreased spermatozoa omega-3 fatty acid levels and increased oxidative DNA damage with varicocele in infertile men: A case control study. Reprod. Fertil. Dev. 2016, 28, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Moretti, E.; Noto, D.; Iacoponi, F.; Signorini, C. Fatty acid profile and metabolism are related to human sperm parameters and are relevant in idiopathic infertility and varicocele. Mediat. Inflamm. 2020, 2020, 3640450. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Hosseini, S.Y.; Dadkhah, F.; Asgari, M.A. Relationship of omega-3 and omega-6 fatty acids with semen characteristics, and anti-oxidant status of seminal plasma: A comparison between fertile and infertile men. Clin. Nutr. 2010, 29, 100–105. [Google Scholar] [CrossRef]
- Tang, B.; Shang, X.; Qi, H.; Li, J.; Ma, B.; An, G.; Zhang, Q. Metabonomic analysis of fatty acids in seminal plasma between healthy and asthenozoospermic men based on gas chromatography mass spectrometry. Andrologia 2017, 49, e12744. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Fang, Y.; Su, Y.; Xu, J.; Hu, Z.; Zhao, K.; Liu, C.; Zhang, H. Varicocele-mediated male infertility: From the perspective of testicular immunity and inflammation. Front. Immunol. 2021, 12, 729539. [Google Scholar] [CrossRef] [PubMed]
- Collodel, G.; Castellini, C.; Lee, J.C.; Signorini, C. Relevance of fatty acids to sperm maturation and quality. Oxid. Med. Cell. Longev. 2020, 2020, 7038124. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, V.; Shahverdi, A.H.; Moghadasian, M.H.; Alizadeh, A.R. Dietary fatty acids affect semen quality: A review. Andrology 2015, 3, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Heitmann, B.L.; Blomberg Jensen, M.; Halldorsson, T.I.; Andersson, A.M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgård, C.; et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Chavarro, J.E.; Mínguez-Alarcón, L.; Mendiola, J.; Cutillas-Tolín, A.; López-Espín, J.J.; Torres-Cantero, A.M. Trans fatty acid intake is inversely related to total sperm count in young healthy men. Hum. Reprod. 2014, 29, 429–440. [Google Scholar] [CrossRef]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.R.; Baghestani, A.R.; Hekmatdoost, A. Dietary fatty acid intakes and asthenozoospermia: A case-control study. Fertil. Steril. 2015, 103, 190–198. [Google Scholar] [CrossRef]
- Nassan, F.L.; Chavarro, J.E.; Tanrikut, C. Diet and men’s fertility: Does diet affect sperm quality? Fertil. Steril. 2018, 110, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Çekici, H.; Akdevelioğlu, Y. The association between trans fatty acids, infertility and fetal life: A review. Hum. Fertil. 2019, 22, 154–163. [Google Scholar] [CrossRef]
- Ferramosca, A.; Moscatelli, N.; Di Giacomo, M.; Zara, V. Dietary fatty acids influence sperm quality and function. Andrology 2017, 5, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Bunay, J.; Gallardo, L.M.; Torres-Fuentes, J.L.; Aguirre-Arias, M.V.; Orellana, R.; Sepúlveda, N.; Moreno, R.D. A decrease of docosahexaenoic acid in testes of mice fed a high-fat diet is associated with impaired sperm acrosome reaction and fertility. Asian J. Androl. 2021, 23, 306–313. [Google Scholar] [CrossRef]
- Moallem, U.; Neta, N.; Zeron, Y.; Zachut, M.; Roth, Z. Dietary α-linolenic acid from flaxseed oil or eicosapentaenoic and docosahexaenoic acids from fish oil differentially alter fatty acid composition and characteristics of fresh and frozen-thawed bull semen. Theriogenology 2015, 83, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.J. Omega-3 fatty acid supplementation and cardiovascular disease events. JAMA 2013, 309, 27. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.S.; Yoshida, Y.; Erickson, K.L. Do n-3 polyunsaturated fatty acids increase or decrease lipid peroxidation in humans? Metab. Syndr. Relat. Disord. 2014, 12, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Mattioli, S.; Signorini, C.; Cotozzolo, E.; Noto, D.; Moretti, E.; Brecchia, G.; Dal Bosco, A.; Belmonte, G.; Durand, T.; et al. Effect of dietary n-3 source on rabbit male reproduction. Oxid. Med. Cell. Longev. 2019, 2019, 3279670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menchetti, L.; Barbato, O.; Sforna, M.; Vigo, D.; Mattioli, S.; Curone, G.; Tecilla, M.; Riva, F.; Brecchia, G. Effects of diets enriched in linseed and fish oil on the expression pattern of Toll-Like Receptors 4 and proinflammatory cytokines on gonadal axis and reproductive organs in rabbit buck. Oxid. Med. Cell. Longev. 2020, 2020, 4327470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attaman, J.A.; Toth, T.L.; Furtado, J.; Campos, H.; Hauser, R.; Chavarro, J.E. Dietary fat and semen quality among men attending a fertility clinic. Hum. Reprod. 2012, 27, 1466–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karayiannis, D.; Kontogianni, M.D.; Mendorou, C.; Douka, L.; Mastrominas, M.; Yiannakouris, N. Association between adherence to the mediterranean diet and semen quality parameters in male partners of couples attempting fertility. Hum. Reprod. 2017, 32, 215–222. [Google Scholar] [CrossRef]
- Vujkovic, M.; de Vries, J.H.; Dohle, G.R.; Bonsel, G.J.; Lindemans, J.; Macklon, N.S.; van der Spek, P.J.; Steegers, E.A.; Steegers-Theunissen, R.P. Associations between dietary patterns and semen quality in men undergoing IVF/ICSI treatment. Hum. Reprod. 2009, 24, 1304–1312. [Google Scholar] [CrossRef]
- MInguez-Alarcón, L.; Chavarro, J.E.; Mendiola, J.; Roca, M.; Tanrikut, C.; Vioque, J.; Jørgensen, N.; Torres-Cantero, A.M. Fatty acid intake in relation to reproductive hormones and testicular volume among young healthy men. Asian J. Androl. 2017, 19, 184–190. [Google Scholar] [CrossRef]
- Safarinejad, M.R.; Safarinejad, S. The roles of omega-3 and omega-6 fatty acids in idiopathic male infertility. Asian J. Androl. 2012, 14, 514–515. [Google Scholar] [CrossRef] [Green Version]
- Blesbois, E.; Douard, V.; Germain, M.; Boniface, P.; Pellet, F. Effects of n-3 polyunsaturated dietary supplementation on the reproductive capacity of male turkeys. Theriogenology 2004, 61, 537–549. [Google Scholar] [CrossRef]
- Abayasekara, D.R.; Wathes, D.C. Effects of altering dietary fatty acid composition on prostaglandin synthesis and fertility. Prostaglandins Leukot. Essent. Fatty Acids 1999, 61, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Roqueta-Rivera, M.; Stroud, C.K.; Haschek, W.M.; Akare, S.J.; Segre, M.; Brush, R.S.; Agbaga, M.P.; Anderson, R.E.; Hess, R.A.; Nakamura, M.T. Docosahexaenoic acid supplementation fully restores fertility and spermatogenesis in male delta-6 desaturase-null mice. J. Lipid Res. 2010, 51, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, B.; Nourmohamadi, M.; Hajipour, S.; Taghizadeh, M.; Asemi, Z.; Keshavarz, S.A.; Jafarnejad, S. The effect of omega-3 fatty acids, EPA, and/or DHA on male infertility: A systematic review and meta-analysis. J. Diet. Suppl. 2019, 16, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Falsig, A.L.; Gleerup, C.S.; Knudsen, U.B. The influence of omega-3 fatty acids on semen quality markers: A systematic PRISMA review. Andrology 2019, 7, 794–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoracka, K.; Eder, P.; Łykowska-Szuber, L.; Dobrowolska, A.; Krela-Kaźmierczak, I. Diet and nutritional factors in male (in)fertility-underestimated factors. J. Clin. Med. 2020, 9, 1400. [Google Scholar] [CrossRef] [PubMed]
- Walczak-Jedrzejowska, R.; Wolski, J.K.; Slowikowska-Hilczer, J. The role of oxidative stress and antioxidants in male fertility. Cent. Eur. J. Urol. 2013, 66, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review evaluation and management. Arab. J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, T.; Usui, K.; Mori, K.; Asai, T.; Yasuda, K.; Kuroda, S.; Yumura, Y. Oxidative stress and male infertility. Reprod. Med. Biol. 2020, 20, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Galano, J.M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C. Isoprostanes, neuroprostanes and phytoprostanes: An overview of 25 years of research in chemistry and biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef]
- Milne, G.L.; Yin, H.; Hardy, K.D.; Davies, S.S.; Roberts, L.J. Isoprostane generation and function. Chem. Rev. 2011, 111, 5973–5996. [Google Scholar] [CrossRef] [Green Version]
- Milne, G.L.; Dai, Q.; Roberts, L.J. The isoprostanes-25 years later. Biochim. Biophys. Acta 2015, 1851, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Menzel, A.; Samouda, H.; Dohet, F.; Loap, S.; Ellulu, M.S.; Bohn, T. Common and novel markers for measuring inflammation and oxidative stress ex vivo in research and clinical practice-which to use regarding disease outcomes? Antioxidants 2021, 10, 414. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Collodel, G. Role of isoprostanes in human male infertility. Syst. Biol. Reprod. Med. 2020, 66, 291–299. [Google Scholar] [CrossRef]
- van’t Erve, T.J.; Kadiiska, M.B.; London, S.J.; Mason, R.P. Classifying oxidative stress by F2-isoprostane levels across human diseases: A meta-analysis. Redox Biol. 2017, 12, 582–599. [Google Scholar] [CrossRef]
- Zhang, Z.J. Systematic review on the association between F2-isoprostanes and cardiovascular disease. Ann. Clin. Biochem. 2013, 50 Pt 2, 108–114. [Google Scholar] [CrossRef]
- Collodel, G.; Castellini, C.; Iacoponi, F.; Noto, D.; Signorini, C. Cytosolic phospholipase A2 and F2 isoprostanes are involved in semen quality and human infertility-A study on leucocytospermia, varicocele and idiopathic infertility. Andrologia 2020, 52, e13465. [Google Scholar] [CrossRef]
- Longini, M.; Moretti, E.; Signorini, C.; Noto, D.; Iacoponi, F.; Collodel, G. Relevance of seminal F2-dihomo-IsoPs, F2-IsoPs and F4-NeuroPs in idiopathic infertility and varicocele. Prostaglandins Other Lipid Mediat. 2020, 149, 106448. [Google Scholar] [CrossRef]
- Moretti, E.; Signorini, C.; Ferretti, F.; Noto, D.; Collodel, G. A Study to validate the relevance of semen F2-Isoprostanes on human male infertility. Int. J. Environ. Res. Public Health 2022, 19, 1642. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Longobardi, S.; Di Rella, F.; Adiga, S.K.; Talevi, R. Sperm oxidative stress during in vitro manipulation and its effects on sperm function and embryo development. Antioxidants 2021, 10, 1025. [Google Scholar] [CrossRef]
- Noto, D.; Collodel, G.; Cerretani, D.; Signorini, C.; Gambera, L.; Menchiari, A.; Moretti, E. Protective effect of chlorogenic acid on human sperm: In vitro studies and frozen-thawed protocol. Antioxidants 2021, 10, 744. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Da Silva, M.S.; Bilodeau, J.F.; Julien, P.; Rudkowska, I. Dietary fats and F2-isoprostanes: A review of the clinical evidence. Crit. Rev. Food Sci. Nutr. 2017, 57, 3929–3941. [Google Scholar] [CrossRef]
- Meyer, K.A.; Sijtsma, F.P.; Nettleton, J.A.; Steffen, L.M.; Van Horn, L.; Shikany, J.M.; Gross, M.D.; Mursu, J.; Traber, M.G.; Jacobs, D.R., Jr. Dietary patterns are associated with plasma F2-isoprostanes in an observational cohort study of adults. Free Radic. Biol. Med. 2013, 57, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Ribas-Maynou, J.; Yeste, M. Oxidative stress in male infertility: Causes, effects in assisted reproductive techniques, and protective support of antioxidants. Biology 2020, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Marion-Letellier, R.; Déchelotte, P.; Iacucci, M.; Ghosh, S. Dietary modulation of peroxisome proliferator-activated receptor gamma. Gut 2009, 58, 586–593. [Google Scholar] [CrossRef]
- Collodel, G.; Noto, D.; Signorini, C.; Gambera, L.; Stendardi, A.; Mahmutbegovic, A.; Micheli, L.; Menchiari, A.; Moretti, E. Do seminal isoprostanes have a role in assisted reproduction outcome? Life 2021, 11, 675. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Noto, D.; Mattioli, S.; Castellini, C.; Pascarelli, N.A.; Durand, T.; Oger, C.; Galano, J.M.; De Felice, C.; et al. F4-Neuroprostanes: A role in sperm capacitation. Life 2021, 11, 655. [Google Scholar] [CrossRef] [PubMed]
- Ayub, M.; Jin, H.K.; Bae, J.S. Sphingosine kinase-dependent regulation of pro-resolving lipid mediators in Alzheimer’s disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159126. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P. The inflammation theory of disease. The growing realization that chronic inflammation is crucial in many diseases opens new avenues for treatment. EMBO Rep. 2012, 13, 968–970. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Brain, S.D.; Buckley, C.D.; Gilroy, D.W.; Haslett, C.; O’Neill, L.A.; Perretti, M.; Rossi, A.G.; Wallace, J.L. Resolution of inflammation: State of the art, definitions and terms. FASEB J. 2007, 21, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Serhan, C.N. Specialized pro-resolving mediator network: An update on production and actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar] [CrossRef]
- Motrich, R.D.; Salazar, F.C.; Breser, M.L.; Mackern-Oberti, J.P.; Godoy, G.J.; Olivera, C.; Paira, D.A.; Rivero, V.E. Implications of prostate inflammation on male fertility. Andrologia 2018, 50, e13093. [Google Scholar] [CrossRef] [Green Version]
- Haidl, G.; Haidl, F.; Allam, J.P.; Schuppe, H.C. Therapeutic options in male genital tract inflammation. Andrologia 2019, 51, e13207. [Google Scholar] [CrossRef]
- Fijak, M.; Pilatz, A.; Hedger, M.P.; Nicolas, N.; Bhushan, S.; Michel, V.; Tung, K.S.K.; Schuppe, H.C.; Meinhardt, A. Infectious, inflammatory and ‘autoimmune’ male factor infertility: How do rodent models inform clinical practice? Hum. Reprod. Update 2018, 24, 416–441. [Google Scholar] [CrossRef] [Green Version]
- Recchiuti, A. Resolvin D1 and its GPCRs in resolution circuits of inflammation. Prostaglandins Other Lipid Mediat. 2013, 107, 64–76. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Noto, D.; Micheli, L.; Ponchia, R.; Collodel, G. Fatty acid oxidation and pro-resolving lipid mediators are related to male infertility. Antioxidants 2022, 11, 107. [Google Scholar] [CrossRef]
- Elliott, E.; Hanson, C.K.; Anderson-Berry, A.L.; Nordgren, T.M. The role of specialized pro-resolving mediators in maternal-fetal health. Prostaglandins Leukot. Essent. Fat. Acids 2017, 126, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Mark, P.J.; Keelan, J.A.; Barden, A.; Mas, E.; Mori, T.A.; Waddell, B.J. Maternal dietary omega-3 fatty acid intake increases resolvin and protectin levels in the rat placenta. J. Lipid Res. 2013, 54, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regidor, P.A.; de la Rosa, X.; Müller, A.; Mayr, M.; Gonzalez Santos, F.; Gracia Banzo, R.; Rizo, J.M. PCOS: A chronic disease that fails to produce adequately specialized pro-resolving lipid mediators (SPMs). Biomedicines 2022, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Z.; Li, H.; Zhu, M.; Peng, X.; Xin, A.; Qu, R.; He, W.; Fu, J.; Sun, X. Resolvin E1 in Follicular fluid acts as a potential biomarker and improves oocyte developmental competence by optimizing cumulus cells. Front. Endocrinol. 2020, 11, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Zhang, L.; Li, J.; Fang, Z.; Li, S.; Peng, Y.; Zhang, M.; Wang, X. Effect of RvD1/FPR2 on inflammatory response in chorioamnionitis. J. Cell. Mol. Med. 2020, 24, 13397–13407. [Google Scholar] [CrossRef] [PubMed]
- Perucci, L.O.; Santos, T.A.P.; Santos, P.C.; Teixeira, L.C.R.; Alpoim, P.N.; Gomes, K.B.; Sousa, L.P.; Dusse, L.M.S.; Talvani, A. Pre-eclampsia is associated with reduced resolvin D1 and maresin 1 to leukotriene B4 ratios in the plasma. Am. J. Reprod. Immunol. 2020, 83, e13206. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collodel, G.; Moretti, E.; Noto, D.; Corsaro, R.; Signorini, C. Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility. Antioxidants 2022, 11, 1002. https://doi.org/10.3390/antiox11051002
Collodel G, Moretti E, Noto D, Corsaro R, Signorini C. Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility. Antioxidants. 2022; 11(5):1002. https://doi.org/10.3390/antiox11051002
Chicago/Turabian StyleCollodel, Giulia, Elena Moretti, Daria Noto, Roberta Corsaro, and Cinzia Signorini. 2022. "Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility" Antioxidants 11, no. 5: 1002. https://doi.org/10.3390/antiox11051002
APA StyleCollodel, G., Moretti, E., Noto, D., Corsaro, R., & Signorini, C. (2022). Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility. Antioxidants, 11(5), 1002. https://doi.org/10.3390/antiox11051002