
White Wine—Induced Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wine Samples
2.2. Laboratory Animals
2.3. Experiments on Isolated Aortic Rings
2.4. Gallic Acid and Quercetin Supplementation
2.5. Determination of Phenol Content
2.6. Determination of Flavonoid Content
2.7. Determination of Antioxidant Activity Using the DPPH, TBARS and FRAP Methods
2.8. Statistical Analysis
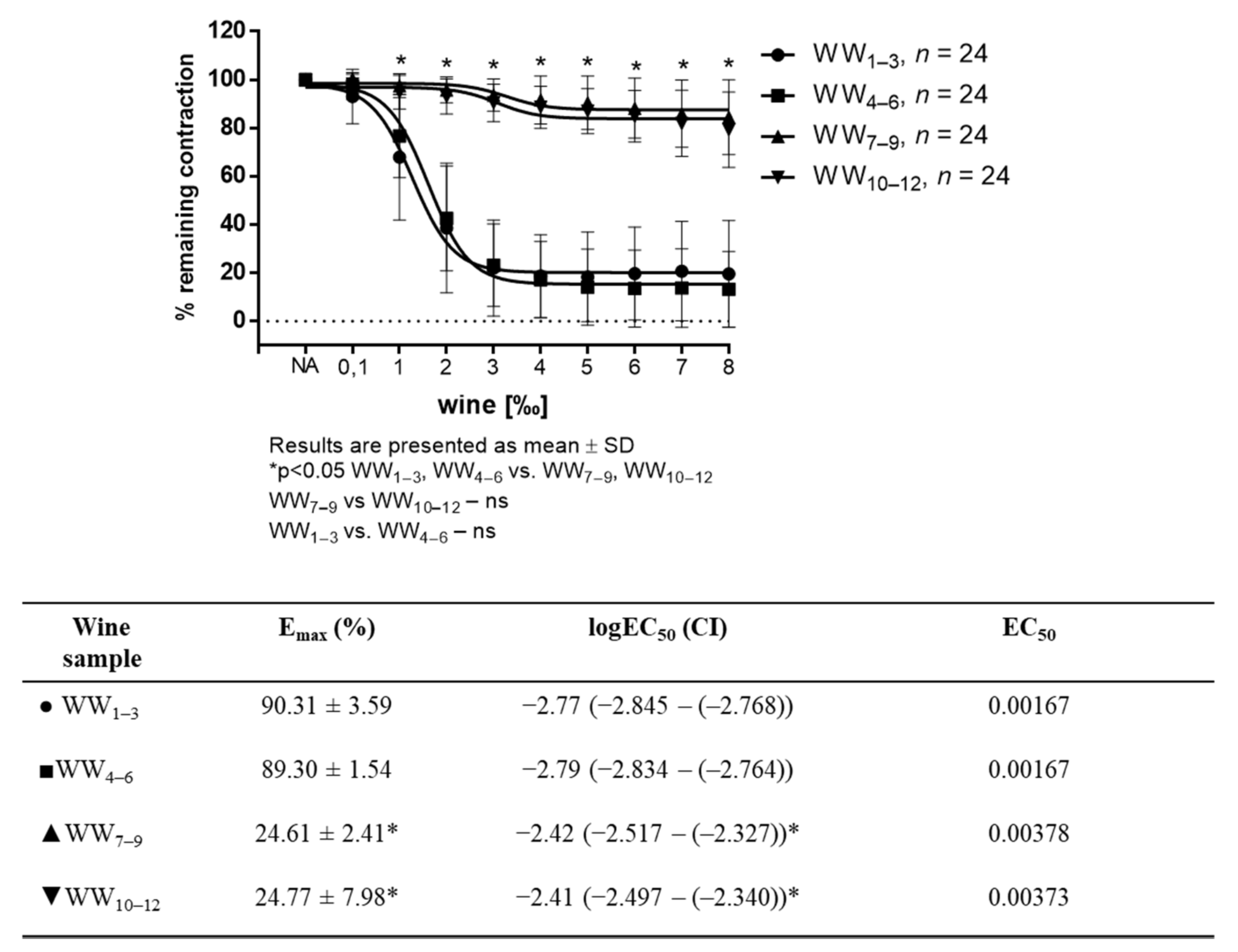
3. Results
3.1. Experiments in Isolated Aortic Rings
3.2. Wine Content Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fornoni, A.; Raij, L. Metabolic syndrome and endothelial dysfunction. Curr. Hypertens. Rep. 2005, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Godo, S.; Shimokawa, H. Endothelial Functions. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e108–e114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousoulis, D.; Kampoli, A.M.; Tentolouris, C.; Papageorgiou, N.; Stefanadis, C. The role of nitric oxide on endothelial function. Curr. Vasc. Pharmacol. 2012, 10, 4–18. [Google Scholar] [CrossRef]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef]
- Suganya, N.; Dornadula, S.; Chatterjee, S.; Mohanram, R.K. Quercetin improves endothelial function in diabetic rats through inhibition of endoplasmic reticulum stress-mediated oxidative stress. Eur. J. Pharmacol. 2018, 819, 80–88. [Google Scholar] [CrossRef]
- Ajay, M.; Achike, F.I.; Mustafa, A.M.; Mustafa, M.R. Effect of quercetin on altered vascular reactivity in aortas isolated from streptozotocin-induced diabetic rats. Diabetes Res. Clin. Pract. 2006, 73, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chis, I.C.; Coseriu, A.; Simedrea, R.; Oros, A.; Nagy, A.L.; Clichici, S. In Vivo Effects of Quercetin in Association with Moderate Exercise Training in Improving Streptozotocin-Induced Aortic Tissue Injuries. Molecules 2015, 20, 21770–21786. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, K.; Tano, I.; Kaneko, N.; Matsumoto, T.; Kobayashi, T. Plant polyphenols Morin and Quercetin rescue nitric oxide production in diabetic mouse aorta through distinct pathways. Biomed. Pharmacother. 2020, 129, 110463. [Google Scholar] [CrossRef]
- Fitzpatrick, D.F.; Hirschfield, S.L.; Coffey, R.G. Endothelium-dependent vasorelaxing activity of wine and other grape products. Am. J. Physiol. 1993, 265 Pt 2, H774–H778. [Google Scholar] [CrossRef]
- Flesch, M.; Schwarz, A.; Böhm, M. Effects of red and white wine on endothelium-dependent vasorelaxation of rat aorta and human coronary arteries. Am. J. Physiol. 1998, 275, H1183–H1190. [Google Scholar] [CrossRef]
- Stoclet, J.C.; Chataigneau, T.; Ndiaye, M.; Oak, M.H.; El Bedoui, J.; Chataigneau, M.; Schini-Kerth, V.B. Vascular protection by dietary polyphenols. Eur. J. Pharmacol. 2004, 500, 299–313. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Spencer, J.P.; Abd El Mohsen, M.M.; Minihane, A.M.; Mathers, J.C. Biomarkers of the intake of dietary polyphenols: Strengths, limitations and application in nutrition research. Br. J. Nutr. 2008, 99, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Galleano, M.; Pechanova, O.; Fraga, C.G. Hypertension, nitric oxide, oxidants, and dietary plant polyphenols. Curr. Pharm. Biotechnol. 2010, 11, 837–848. [Google Scholar] [CrossRef]
- Du, L.; Chen, E.; Wu, T.; Ruan, Y.; Wu, S. Resveratrol attenuates hydrogen peroxide-induced aging through upregulation of autophagy in human umbilical vein endothelial cells. Drug Des. Devel. 2019, 13, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.L.; Lyon, T.; Litwin, S.E.; Rabovsky, A.; Symons, J.D.; Jalili, T. Quercetin reduces blood pressure in hypertensive subjects. J. Nutr. 2007, 137, 2405–2411. [Google Scholar] [CrossRef]
- Xiao, X.; Shi, D.; Liu, L.; Wang, J.; Xie, X.; Kang, T.; Deng, W. Quercetin suppresses cyclooxygenase-2 expression and angiogenesis through inactivation of P300 signaling. PLoS ONE 2011, 6, e22934. [Google Scholar] [CrossRef]
- Tanaka, S.; Trakooncharoenvit, A.; Nishikawa, M.; Ikushiro, S.; Hara, H. Heteroconjugates of quercetin with 4′-O-sulfate selectively accumulate in rat plasma due to limited urinary excretion. Food Funct. 2022, 13, 1459–1471. [Google Scholar] [CrossRef]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Chopra, M.; Fitzsimons, P.E.; Strain, J.J.; Thurnham, D.I.; Howard, A.N. Nonalcoholic red wine extract and quercetin inhibit LDL oxidation without affecting plasma antioxidant vitamin and carotenoid concentrations. Clin. Chem. 2000, 46 Pt 1, 1162–1170. [Google Scholar] [CrossRef]
- Salvamani, S.; Gunasekaran, B.; Shaharuddin, N.A.; Ahmad, S.A.; Shukor, M.Y. Antiartherosclerotic effects of plant flavonoids. Biomed. Res. Int. 2014, 2014, 480258. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yin, O.Q.; Zuo, Z.; Chow, M.S. Pharmacokinetics and modeling of quercetin and metabolites. Pharm. Res. 2005, 22, 892–901. [Google Scholar] [CrossRef]
- García-Mediavilla, V.; Crespo, I.; Collado, P.S.; Esteller, A.; Sánchez-Campos, S.; Tuñón, M.J.; González-Gallego, J. The anti-inflammatory flavones quercetin and kaempferol cause inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor kappaB pathway in Chang Liver cells. Eur. J. Pharmacol. 2007, 557, 221–229. [Google Scholar] [CrossRef]
- Ha, S.K.; Park, H.Y.; Ryu, M.R.; Kim, Y.; Park, Y. Endothelium-Dependent Vasorelaxant Effects of Dealcoholized Wine Powder of Wild Grape (Vitis coignetiae) in the Rat Thoracic Aorta. Evid. Based Complement Alternat. Med. 2016, 2016, 6846084. [Google Scholar] [CrossRef] [Green Version]
- Caimi, G.; Carollo, C.; Lo Presti, R. Wine and endothelial function. Drugs Exp. Clin. Res. 2003, 29, 235–242. [Google Scholar]
- Fuhrman, B.; Volkova, N.; Suraski, A.; Aviram, M. White wine with red wine-like properties: Increased extraction of grape skin polyphenols improves the antioxidant capacity of the derived white wine. J. Agric. Food Chem. 2001, 49, 3164–3168. [Google Scholar] [CrossRef]
- Unfirer, S.; Mihalj, M.; Novak, S.; Kibel, A.; Cavka, A.; Mijalevic, Z.; Gros, M.; Brizic, I.; Budimir, D.; Cosic, A.; et al. Hyperbaric oxygenation affects the mechanisms of acetylcholine-induced relaxation in diabetic rats. Undersea Hyperb. Med. 2016, 43, 787–803. [Google Scholar]
- Kibel, A.; Cavka, A.; Cosic, A.; Falck, J.R.; Drenjancevic, I. Effects of hyperbaric oxygenation on vascular reactivity to angiotensin II and angiotensin-(1-7) in rats. Undersea Hyperb. Med. 2012, 39, 1053–1066. [Google Scholar]
- Kibel, A.; Novak, S.; Cosic, A.; Mihaljevic, Z.; Falck, J.R.; Drenjancevic, I. Hyperbaric oxygenation modulates vascular reactivity to angiotensin-(1-7) in diabetic rats: Potential role of epoxyeicosatrienoic acids. Diabetes Vasc. Dis. Res. 2015, 12, 33–45. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Rašić, L.; Jukić, I.; Drenjančević, I. Acute Hyperbaric Oxygenation, Contrary to Intermittent Hyperbaric Oxygenation, Adversely Affects Vasorelaxation in Healthy Sprague-Dawley Rats due to Increased Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 7406027. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Frkanec, R.; Tavčar, B.; Kelava, V.; Tartaro Bujak, I.; Kolobarić, N.; Kibel, A.; Drenjančević, I. Arachidonic Acid Metabolites of CYP450 Enzymes and HIF-1α Modulate Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats under Acute and Intermittent Hyperbaric Oxygenation. Int. J. Mol. Sci. 2020, 21, 6353. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phospho-molybdic-phosphotungistic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Ordoñez, A.A.L.; Gomez, J.D.; Vattuone, M.A.; Isla, M.I. Antioxidant activities of Sechium edule (Jacq.) Swart extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use Of A Free Radical Method To Evaluate Antioxidant Activity. Lebensm.-Wiss. Technol.-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under NaCl stress. J. Agric. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef]
- Torres, A.; Cachofeiro, V.; Millán, J.; Lahera, V.; Nieto, M.L.; Martín, R.; Bello, E.; Alvarez-Sala, L.A. Red wine intake but not other alcoholic beverages increases total antioxidant capacity and improves pro-inflammatory profile after an oral fat diet in healthy volunteers. Rev. Clin. Esp. 2015, 215, 486–494. [Google Scholar] [CrossRef]
- Sparwel, J.; Vantler, M.; Caglayan, E.; Kappert, K.; Fries, J.W.; Dietrich, H.; Böhm, M.; Erdmann, E.; Rosenkranz, S. Differential effects of red and white wines on inhibition of the platelet-derived growth factor receptor: Impact of the mash fermentation. Cardiovasc. Res. 2009, 81, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Boban, M.; Modun, D.; Music, I.; Vukovic, J.; Brizic, I.; Salamunic, I.; Obad, A.; Palada, I.; Dujic, Z. Red wine induced modulation of vascular function: Separating the role of polyphenols, ethanol, and urates. J. Cardiovasc. Pharmacol. 2006, 47, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Ružić, I.; Škerget, M.; Knez, Ž.; Runje, M. Phenolic content and antioxidant potential of macerated white wines. Eur. Food Res. Technol. 2011, 233, 465–472. [Google Scholar] [CrossRef]
- De Oliveira, L.M.; de Oliveira, T.S.; da Costa, R.M.; de Souza Gil, E.; Costa, E.A.; Passaglia Rde, C.; Filgueira, F.P.; Ghedini, P.C. The vasorelaxant effect of gallic acid involves endothelium-dependent and -independent mechanisms. Vasc. Pharmacol. 2016, 81, 69–74. [Google Scholar] [CrossRef]
- Li, P.G.; Sun, L.; Han, X.; Ling, S.; Gan, W.T.; Xu, J.W. Quercetin induces rapid eNOS phosphorylation and vasodilation by an Akt-independent and PKA-dependent mechanism. Pharmacology 2012, 89, 220–228. [Google Scholar] [CrossRef]
- Mudnic, I.; Modun, D.; Rastija, V.; Vukovic, J.; Brizic, I.; Katalinic, V.; Kozina, B.; Medic-Saric, M.; Boban, M. Antioxidative and vasodilatory effects of phenolic acids in wine. Food Chem. 2010, 119, 1205–1210. [Google Scholar] [CrossRef]
- Roghani, M.; Baluchnejadmojarad, T.; Vaez-Mahdavi, M.R.; Roghani-Dehkordi, F. Mechanisms underlying quercetin-induced vasorelaxation in aorta of subchronic diabetic rats: An in vitro study. Vasc. Pharmacol. 2004, 42, 31–35. [Google Scholar] [CrossRef]
- Kang, K.T. Endothelium-derived Relaxing Factors of Small Resistance Arteries in Hypertension. Toxicol. Res. 2014, 30, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Palmer, R.M.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef]
- Sandoo, A.; van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [Green Version]
- Nong, Z.; Hoylaerts, M.; Van Pelt, N.; Collen, D.; Janssens, S. Nitric oxide inhalation inhibits platelet aggregation and platelet-mediated pulmonary thrombosis in rats. Circ. Res. 1997, 81, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Lefer, D.J.; Jones, S.P.; Girod, W.G.; Baines, A.; Grisham, M.B.; Cockrell, A.S.; Huang, P.L.; Scalia, R. Leukocyte-endothelial cell interactions in nitric oxide synthase-deficient mice. Am. J. Physiol. 1999, 276, H1943–H1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, S.D.; Seggara, G.; Vo, P.A.; Macallister, R.J.; Hobbs, A.J.; Ahluwalia, A. Protection against lipopolysaccharide-induced endothelial dysfunction in resistance and conduit vasculature of iNOS knockout mice. FASEB J. 2003, 17, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.B.; Spiecker, M.; Liao, J.K. Inducible nitric oxide: An autoregulatory feedback inhibitor of vascular inflammation. J. Immunol. 1998, 161, 1970–1976. [Google Scholar] [PubMed]
- Ahluwalia, A.; Foster, P.; Scotland, R.S.; McLean, P.G.; Mathur, A.; Perretti, M.; Moncada, S.; Hobbs, A.J. Antiinflammatory activity of soluble guanylate cyclase: cGMP-dependent down-regulation of P-selectin expression and leukocyte recruitment. Proc. Natl. Acad. Sci. USA 2004, 101, 1386–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollny, T.; Aiello, L.; Di Tommaso, D.; Bellavia, V.; Rotilio, D.; Donati, M.B.; De Gaetano, G.; Iacoviello, L. Modulation of haemostatic function and prevention of experimental thrombosis by red wine in rats: A role for increased nitric oxide production. Br. J. Pharmacol. 1999, 127, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Giglio, R.V.; Patti, A.M.; Cicero, A.F.; Lippi, G.; Rizzo, M.; Toth, P.P.; Banach, M. Polyphenols: Potential Use in the Prevention and Treatment of Cardiovascular Diseases. Curr. Pharm. Des. 2018, 24, 239–258. [Google Scholar] [CrossRef]
- Brizic, I.; Modun, D.; Vukovic, J.; Budimir, D.; Katalinic, V.; Boban, M. Differences in vasodilatory response to red wine in rat and guinea pig aorta. J. Cardiovasc. Pharmacol. 2009, 53, 116–120. [Google Scholar] [CrossRef]
- Krymchantowski, A.V.; da Cunha Jevoux, C. Wine and headache. Headache 2014, 54, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Raposo, R.; Ruiz-Moreno, M.J.; Garde-Cerdán, T.; Puertas, B.; Moreno-Rojas, J.M.; Zafrilla, P.; Gonzalo-Diago, A.; Guerrero, R.F.; Cantos-Villar, E. Replacement of sulfur dioxide by hydroxytyrosol in white wine: Influence on both quality parameters and sensory. LWT-Food Sci. Technol. 2016, 65, 214–221. [Google Scholar] [CrossRef]
- Pravilnik o Izmjenama i Dopunama Pravilnika o Proizvodnji Vina (NN 137/2008). Available online: https://narodne-novine.nn.hr/clanci/sluzbeni/2008_11_137_3851.html (accessed on 3 April 2022).
- Silva, M.; Gama, J.; Pinto, N.; Pivi, G.; Brancal, H.; Carvalho, L.; Loureiro, V.; Patto, M.V. Sulfite concentration and the occurrence of headache in young adults: A prospective study. Eur. J. Clin. Nutr. 2019, 73, 1316–1322. [Google Scholar] [CrossRef]
- Pavić, V.; Kujundžić, T.; Kopić, M.; Jukić, V.; Braun, U.; Schwander, F.; Drenjančević, M. Effects of Defoliation on Phenolic Concentrations, Antioxidant and Antibacterial Activity of Grape Skin Extracts of the Varieties Blaufränkisch and Merlot (Vitis vinifera L.). Molecules 2019, 24, 2444. [Google Scholar] [CrossRef] [Green Version]
- Beltran, G.; Novo, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 2008, 121, 169–177. [Google Scholar] [CrossRef]
- Liu, K.; Liu, Y.; Chen, F. Effect of storage temperature on lipid oxidation and changes in nutrient contents in peanuts. Food Sci. Nutr. 2019, 7, 2280–2290. [Google Scholar] [CrossRef]
- Matute, A.; Tabart, J.; Cheramy-Bien, J.P.; Kevers, C.; Dommes, J.; Defraigne, J.O.; Pincemail, J. Ex Vivo Antioxidant Capacities of Fruit and Vegetable Juices. Potential In Vivo Extrapolation. Antioxidants 2021, 10, 770. [Google Scholar] [CrossRef]
- Shahidi, F.; Peng, H. Bioaccessibility and bioavailability of phenolic compounds. J. Food Bioact. 2018, 4, 11–68. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WW1–3 | WW4–6 | WW7–9 | WW10–12 | |
|---|---|---|---|---|
| Phenol content [mg Gallic acids g−1 FW] | 1.17 ± 0.107 # | 0.81 ± 0.017 * | 0.27 ± 0.011 *# | 0.28 ± 0.004 *# |
| Flavonoid content [µg Quercetine g−1 FW] | 24.07 ± 1.066 | 25.18 ± 1.258 | 16.53 ± 1.478 *# | 14.98 ± 1.434 *# |
| WW1–3 | WW4–6 | WW7–9 | WW10–12 | |
|---|---|---|---|---|
| Alcoholic strength [vol %] | 13.04 ± 0.253 | 13.13 ± 0.006 | 13.71 ± 0.045 *# | 13.72 ± 0.060 *# |
| Total dry extract [g/L] | 23.87 ± 0.568 | 23.57 ± 0.252 | 18.73 ± 0.416 *# | 18.07 ± 0.404 *# |
| Free sulfur dioxide [mg/L] | 12.69 ± 5.143 | 11.73 ± 3.844 | 75.41 ± 4.345 *# | 53.33 ± 4.052 *#† |
| Total sulfur dioxide [mg/L] | 128.00 ± 7.212 | 131.50 ± 6.841 | 126.30 ± 4.541 | 196.40 ± 3.221 *#† |
| Volatile acids [g/L] | 0.70 ± 0.021 | 0.65 ± 0.042 | 0.81 ± 0.026 *# | 0.66 ± 0.017 #† |
| Total acids [g/L] | 4.87 ± 0.130 # | 5.28 ± 0.112 * | 4.80 ± 0.075 *# | 6.20 ± 0.173 *#† |
| pH | 3.73 ± 0.0265 # | 3.60 ± 0.020 * | 3.51 ± 0.045 *# | 2.92 ± 0.030 *#† |
| WW1–3 | WW4–6 | WW7–9 | WW10–12 | |
|---|---|---|---|---|
| FRAP in equivalents Fe(II) [mM/g FW] | 119.30 ± 18.17 # | 74.29 ± 3.003 * | 32.93 ± 1.283 *# | 29.56 ± 1.836 *# |
| TBARS [nmol/g FW] | 13.84 ± 1.307 # | 9.85 ± 1.419 * | 1.47 ± 0.014 *# | 2.79 ± 0.303 *# |
| DPPH 50% EC [mg FW/mL] | 10.65 ± 1.791 | 16.97 ± 0.7992 | 56.93 ± 9.138 *# | 59.25 ± 2.415 *# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihaljević, Z.; Kujundžić, T.; Jukić, V.; Stupin, A.; Drenjančević, M.; Drenjančević, I. White Wine—Induced Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats. Antioxidants 2022, 11, 944. https://doi.org/10.3390/antiox11050944
Mihaljević Z, Kujundžić T, Jukić V, Stupin A, Drenjančević M, Drenjančević I. White Wine—Induced Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats. Antioxidants. 2022; 11(5):944. https://doi.org/10.3390/antiox11050944
Chicago/Turabian StyleMihaljević, Zrinka, Toni Kujundžić, Vladimir Jukić, Ana Stupin, Mato Drenjančević, and Ines Drenjančević. 2022. "White Wine—Induced Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats" Antioxidants 11, no. 5: 944. https://doi.org/10.3390/antiox11050944
APA StyleMihaljević, Z., Kujundžić, T., Jukić, V., Stupin, A., Drenjančević, M., & Drenjančević, I. (2022). White Wine—Induced Endothelium-Dependent Vasorelaxation in Sprague-Dawley Rats. Antioxidants, 11(5), 944. https://doi.org/10.3390/antiox11050944