
Morin Disrupts Cytoskeleton Reorganization in Osteoclasts through an ROS/SHP1/c-Src Axis and Grants Protection from LPS-Induced Bone Loss

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents and Antibodies
2.3. Animals and Study Design
2.4. OC Formation
2.5. Cell Viability
2.6. RNA Isolation and Quantitative Polymerase Chain Reaction (qPCR)
2.7. Bone Resorption
2.8. Actin Cytoskeleton
2.9. Western Blot Analysis
2.10. Determination of Intracellular and Mitochondrial Reactive Oxygen Species (ROS)
2.11. Detection of Oxidized SHP1 by Carboxymethylation
2.12. Rac1 Activity Assay (Pulldown Assay)
2.13. Transfection of siRNA
2.14. Statistical Analyses
3. Results
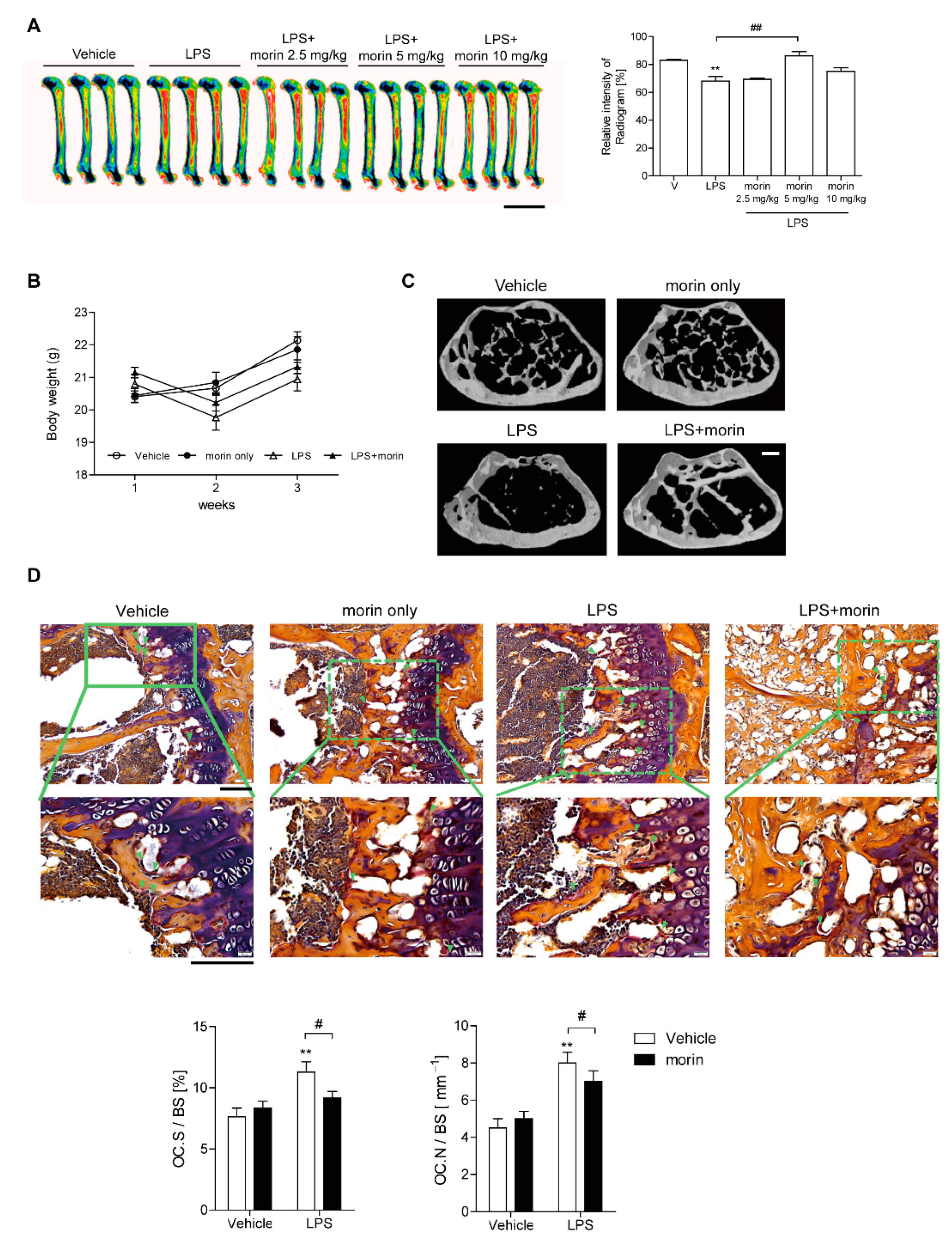
3.1. Morin Protects Mice from LPS-Induced Bone Loss
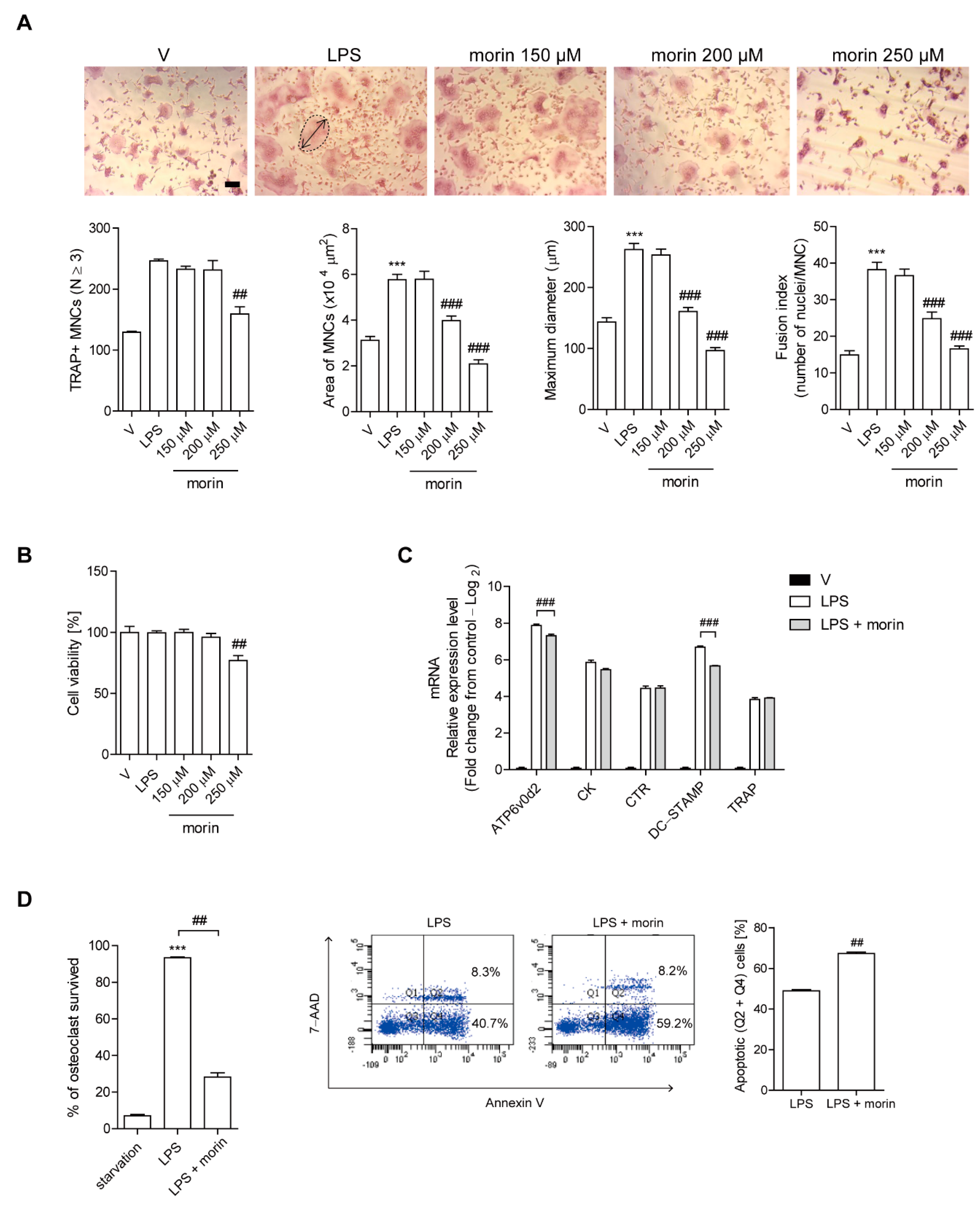
3.2. Morin Reduces the Number of OCs upon LPS Stimulation In Vitro
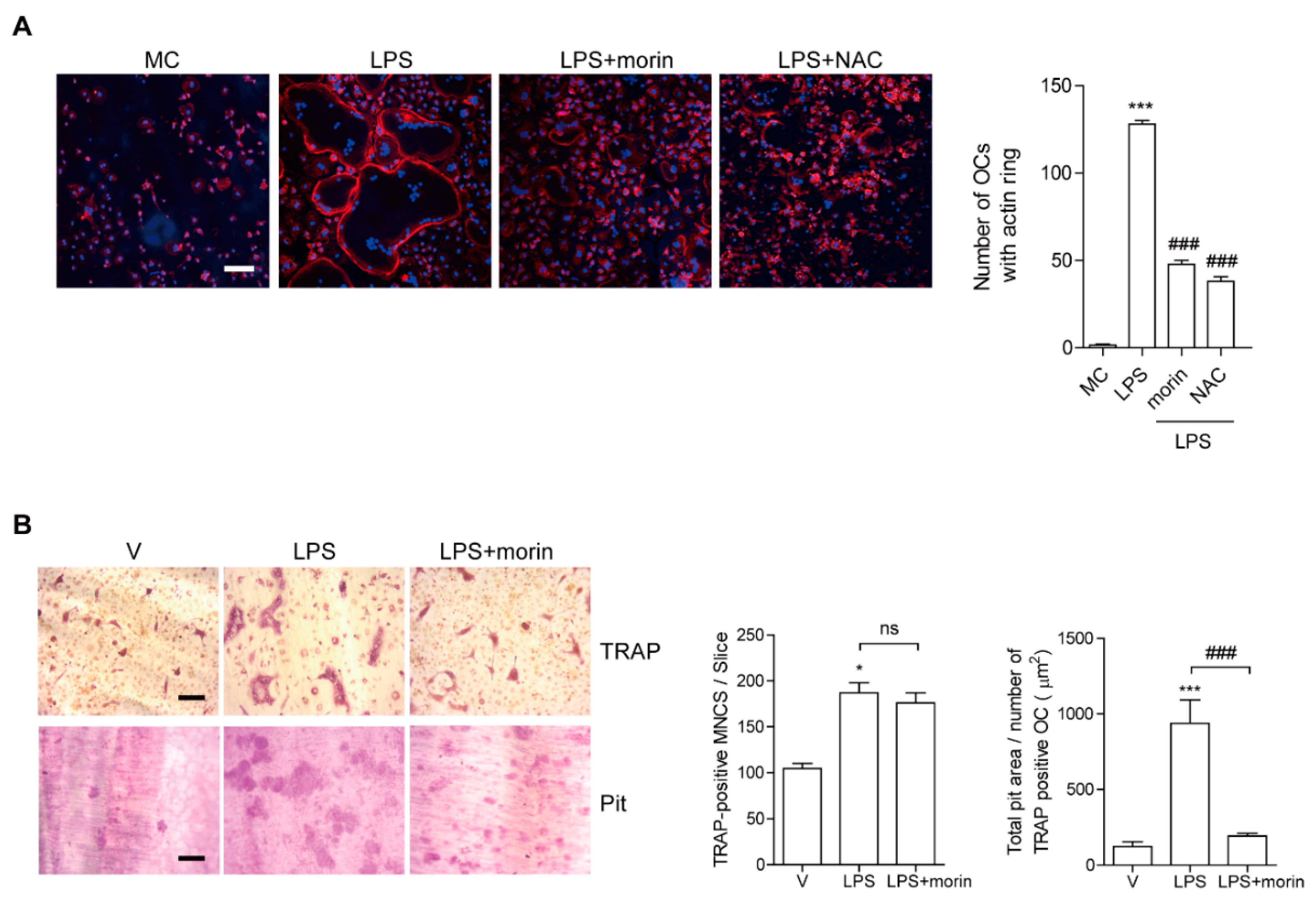
3.3. Morin Inhibits LPS-Stimulated Actin Ring Formation and OC Activity
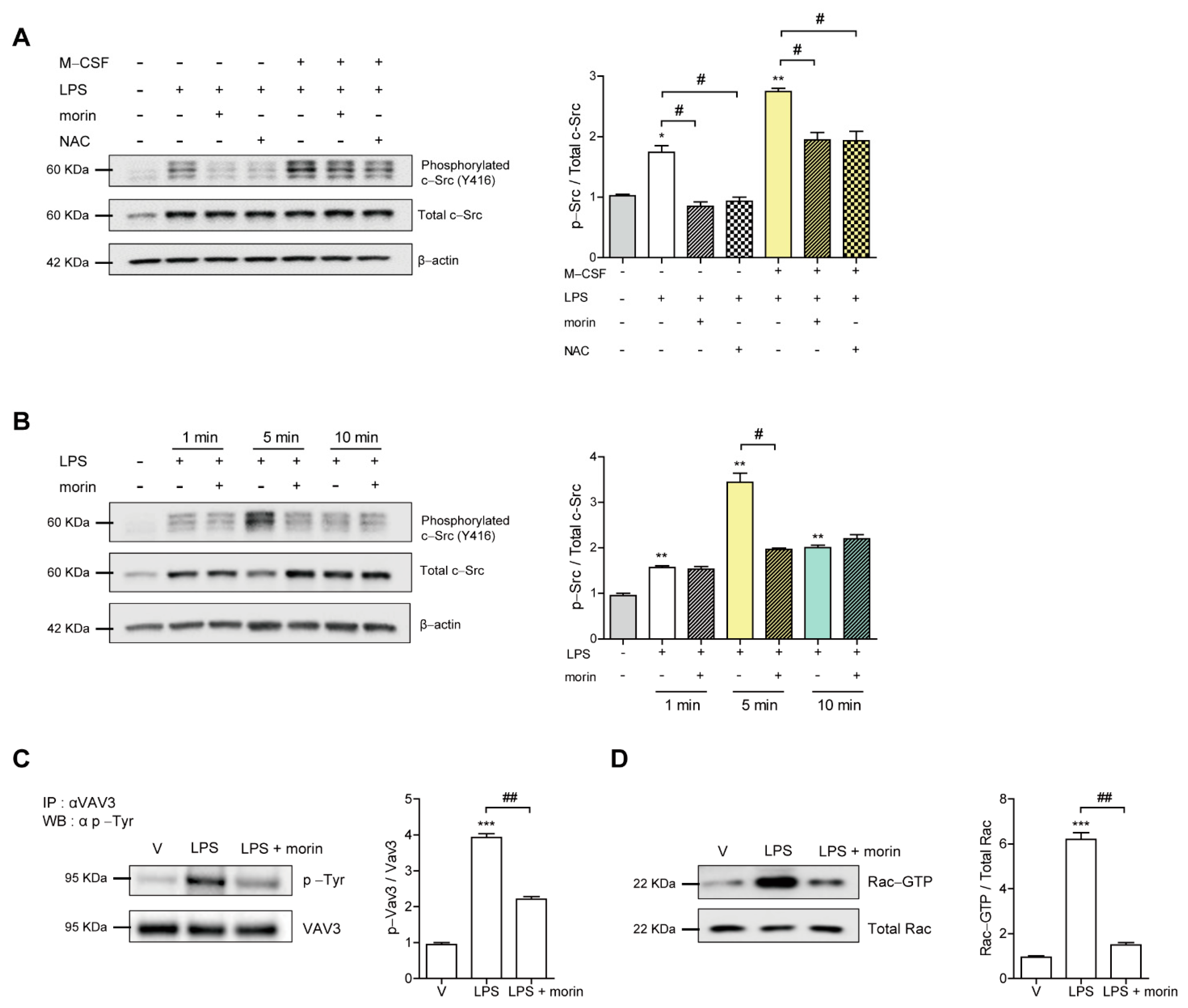
3.4. Morin Decreases LPS-Induced Cytoskeletal Reorganization through a c-Src/Vav3/Rac1 Signaling Pathway in OCs
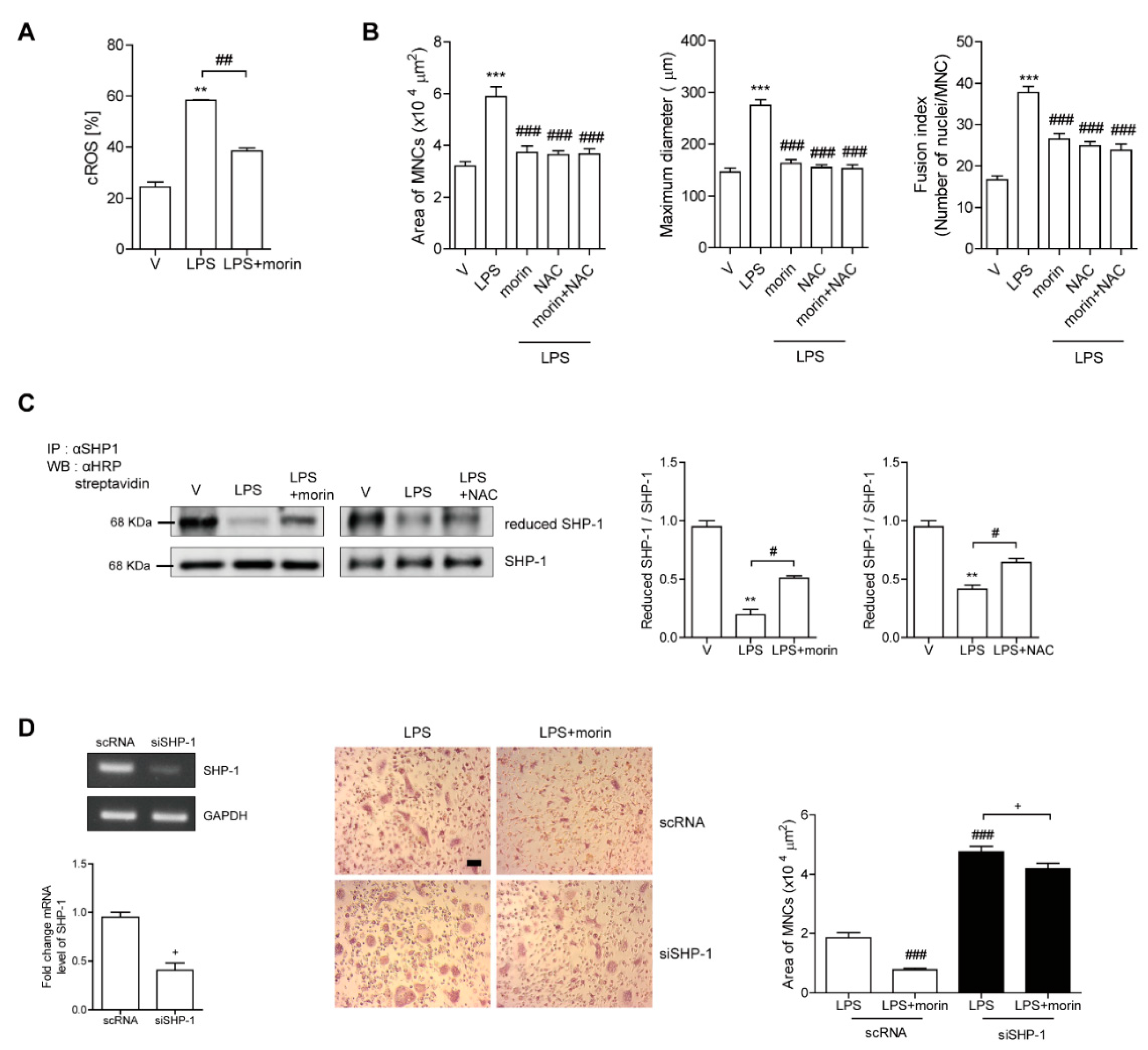
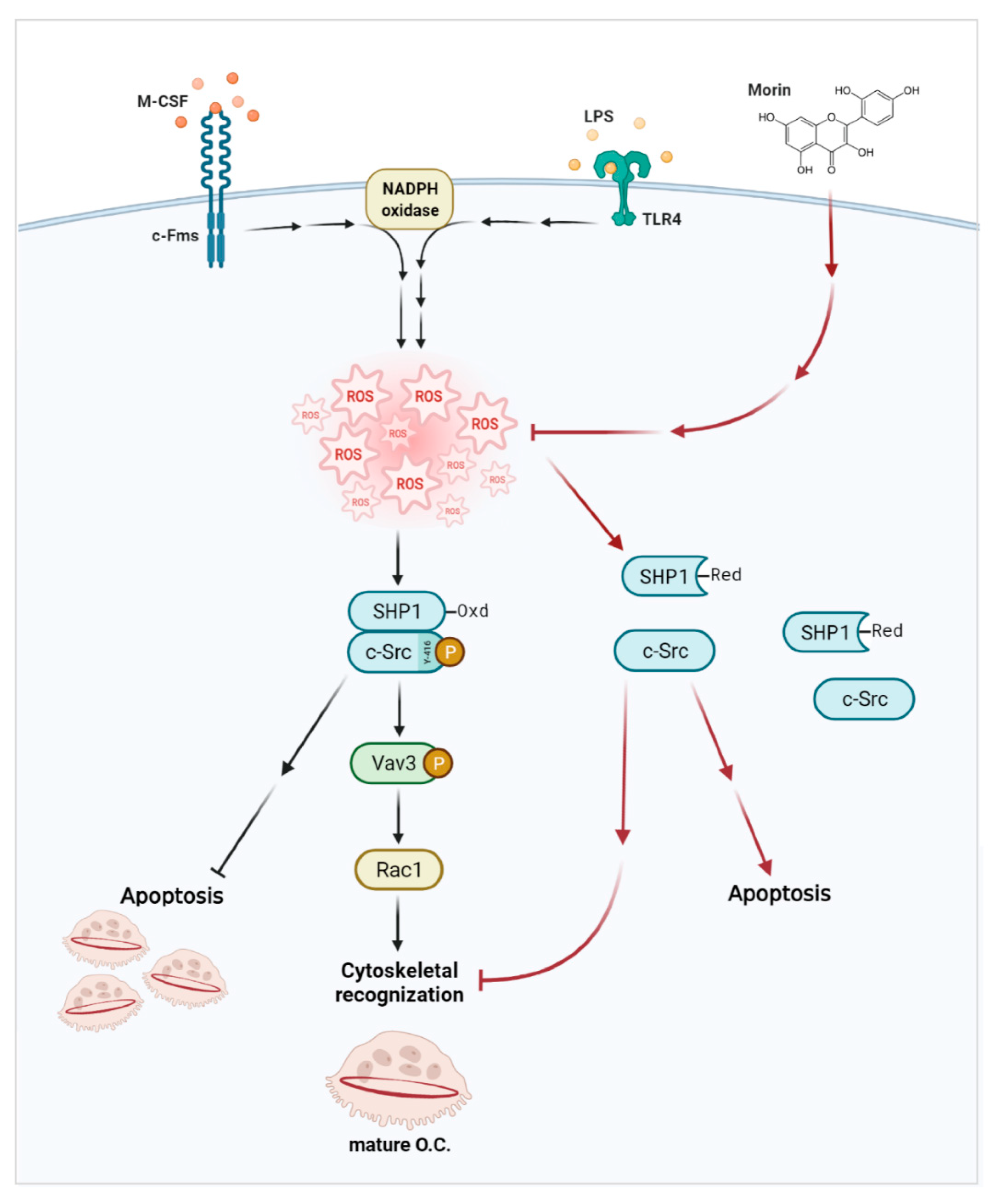
3.5. Morin Decreases LPS-Induced Oxidation of SHP-1 to Attenuate c-Src Activation by Decreasing ROS Levels in OCs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Staa, T.V.; Geusens, P.; Bijlsma, J.W.J.; Leufkens, H.G.M.; Cooper, C. Clinical assessment of the long-term risk of fracture in patients with rheumatoid arthritis. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2006, 54, 3104–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocijan, R.; Englbrecht, M.; Haschka, J.; Simon, D.; Kleyer, A.; Finzel, S.; Kraus, S.; Resch, H.; Muschitz, C.; Engelke, K.; et al. Quantitative and qualitative changes of bone in psoriasis and psoriatic arthritis patients. J. Bone Miner. Res. 2015, 30, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Haschka, J.; Hirschmann, S.; Kleyer, A.; Englbrecht, M.; Faustini, F.; Simon, D.; Figueiredo, C.P.; Schuster, L.; Muschitz, C.; Kocijan, R.; et al. High-resolution quantitative computed tomography demonstrates structural defects in cortical and trabecular bone in IBD patients. J. Crohn’s Colitis. 2016, 10, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Pasco, J.A.; Kotowicz, M.A.; Henry, H.J.; Nicholson, G.C.; Spilsbury, H.J.; Nicholson, G.C.; Spilsbury, H.J.; Box, J.D.; Schneider, H.G. High-sensitivity C-reactive protein and fracture risk in elderly women. JAMA 2006, 296, 1353–1355. [Google Scholar] [CrossRef]
- Gravallese, E.M.; Harada, Y.; Wang, J.T.; Gorn, A.H.; Thornhill, T.S.; Goldring, S.R. Identification of cell types responsible for bone resorption in rheumatoid arthritis and juvenile rheumatoid arthritis. Am. J. Pathol. 1998, 152, 943–951. [Google Scholar]
- Dewhirst, F.E.; Stashenko, P.P.; Mole, J.E.; Tsurumachi, T. Purification and partial sequence of human osteoclast-activating factor: Identity with interleukin 1 beta. J. Immunol. 1985, 135, 2562–2568. [Google Scholar]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]
- Orcel, P.; Feuga, M.; Bielakoff, J.; De Vemejoul, M.C. Local bone injections of LPS and M-CSF increase bone resorption by different pathways in vivo in rats. Am. J. Physiol. 1993, 264, 391–397. [Google Scholar] [CrossRef]
- Park, H.J.; Son, H.J.; Sul, O.J.; Suh, J.H.; Choi, H.S. 4-Phenylbutyric acid protects against lipopolysaccharide-induced bone loss by modulating autophagy in osteoclasts. Biochem. Pharmacol. 2018, 151, 9–17. [Google Scholar] [CrossRef]
- Park, H.J.; Gholam-Zadeh, M.; Suh, J.H.; Choi, H.S. Lycorine attenuates autophagy in osteoclasts via an axis of mROS/TRPML1/TFEB to reduce LPS-induced bone loss. Oxid. Med. Cell. Longev. 2019, 2019, 8982147. [Google Scholar] [CrossRef]
- Park, H.J.; Noh, A.L.; Kang, J.H.; Sim, J.S.; Lee, D.S.; Yim, M. Peroxiredoxin II negatively regulates lipopolysaccharide-induced osteoclast formation and bone loss via JNK and STAT3. Antioxid. Redox Signal. 2015, 22, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sul, O.J.; Park, H.J.; Son, H.J.; Choi, H.S. Lipopolysaccharide (LPS)-induced autophagy is responsible for enhanced osteoclastogenesis. Mol. Cells 2017, 40, 880–887. [Google Scholar]
- Sul, O.J.; Rajasekaran, M.; Park, H.J.; Suh, J.H.; Choi, H.S. MicroRNA-29b enhances osteoclast survival by targeting Bcl2-modifying factor after lipopolysaccharide stimulation. Oxid. Med. Cell. Longev. 2019, 2019, 6018180. [Google Scholar] [CrossRef] [PubMed]
- Sul, O.J.; Sung, Y.B.; Rajasekaran, M.; Ke, K.; Yu, R.; Back, S.Y.; Choi, H.S. MicroRNA-155 induces autophagy in osteoclasts by targeting transforming growth factor β-activated kinase 1-binding protein 2 upon lipopolysaccharide stimulation. Bone 2018, 116, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Blangy, A.; Bompard, G.; Guerit, D.; Marie, P.; Maurin, J.; Morel, A.; Vives, V. The osteoclast cytoskeleton—Current understanding and therapeutic perspectives for osteoporosis. J. Cell Sci. 2020, 133, 244798. [Google Scholar] [CrossRef] [PubMed]
- McHugh, K.P.; Dilke, K.H.; Zheng, M.H.; Namba, N.; Lam, J.; Novack, D.; Feng, X.; Ross, F.P.; Hynes, R.O.; Teitelbaum, S.L. Mice lacking beta3 integrins are osteosclerotic because of dysfunctional osteoclasts. J. Clin. Invest 2000, 105, 433–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Destaing, O.; Sanjay, A.; Itzstein, C.; Horne, W.C.; Toomre, D.; Camilli, P.D.; Baron, R. The tyrosine kinase activity of c-Src regulates actin dynamics and organization of podosomes in osteoclasts. Mol. Biol. Cell 2008, 19, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Soriano, P.; Montgomery, C.; Geske, R.; Bradley, A. Targeted disruption of the c-src proto-oncogene leads to osteopetrosis in mice. Cell 1991, 64, 693–702. [Google Scholar] [CrossRef]
- Faccio, R.; Novack, D.V.; Zallone, A.; Ross, F.P.; Teitelbaum, S.L. Dynamic changes in the osteoclast cytoskeleton in response to growth factors and cell attachment are controlled by beta3 integrin. J. Cell Biol. 2003, 162, 499–509. [Google Scholar] [CrossRef]
- Croke, M.; Ross, F.P.; Korhonen, M.; Williams, D.A.; Zou, W.; Teitelbaum, S.L. Rac deletion in osteoclasts causes severe osteopetrosis. J. Cell Sci. 2011, 124, 3811–3821. [Google Scholar] [CrossRef] [Green Version]
- Faccio, R.; Teitelbaum, S.L.; Fujikawa, K.; Chappel, J.; Zallone, A.; Tybulewicz, V.L.; Ross, F.P.; Swat, W. Vav3 regulates osteoclast function and bone mass. Nat. Med. 2005, 11, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Caselli, A.; Cirri, P.; Santi, A.; Paoli, P. Morin: A promising natural drug. Curr. Med. Chem. 2016, 23, 774–791. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, S.V.; Simic, M.G. Antioxidants in nutrition. Ann. N. Y. Acad. Sci. 2000, 899, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Q.; Huang, J.; Jin, Q.; Xu, B.; Chen, F.; Tu, C. Morin hydrate inhibits TREM-1/TLR4-mediated inflammatory response in macrophages and protects against carbon tetrachloride-induced acute liver injury in mice. Front. Pharmacol. 2019, 10, 1089. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Wang, X.M.; Xu, D.Y.; Sang, L.X.; Han, Y.; Jiang, L.Y. Morin enhances hepatic Nrf2 expression in a liver fibrosis rat model. World J. Gastroenterol. 2017, 23, 8334–8344. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.K.; Malik, S.; Mutneja, E.; Sahu, A.K.; Bhatia, J.; Arya, D.S. Attenuation of ROS-mediated myocardial ischemia-reperfusion injury by morin via regulation of RISK/SAPK pathways. Pharmacol. Rep. 2020, 72, 877–889. [Google Scholar] [CrossRef]
- Wang, C.; Wan, X.; Li, Y.; Zhang, H.; Zhang, L. Morin protects glucocorticoid-induced osteoporosis through regulating the mitogen-activated protein kinase signaling pathway. J. Nat. Med. 2018, 72, 929–936. [Google Scholar] [CrossRef]
- Gao, Y.; Grassi, F.; Ryan, M.R.; Terauchi, M.; Page, K.; Yang, X.; Weitzmann, M.N.; Pacific, R. IFN-gamma stimulates osteoclast formation and bone loss in vivo via antigen-driven T cell activation. J. Clin. Invest 2007, 117, 122–132. [Google Scholar] [CrossRef]
- Phan, T.V.; Sul, O.J.; Ke, K.; Lee, M.H.; Kim, W.K.; Cho, Y.S.; Kim, H.J.; Kim, S.Y.; Chung, H.T.; Choi, H.S. Carbon monoxide protects against ovariectomy-induced bone loss by inhibiting osteoclastogenesis. Biochem. Pharmacol. 2013, 85, 1145–1152. [Google Scholar] [CrossRef]
- Okayasu, M.; Nakayachi, M.; Hayashida, C.; Ito, J.; Kaneda, T.; Masuhara, M.; Suda, N.; Sato, T.; Hakeda, Y. Low-density lipoprotein receptor deficiency causes impaired osteoclastogenesis and increased bone mass in mice because of defect in osteoclastic cell-cell fusion. J. Biol. Chem. 2012, 287, 19229–19241. [Google Scholar] [CrossRef] [Green Version]
- Jimi, E.; Akiyama, S.; Tsurukai, T.; Okahashi, N.; Kobayashi, K.; Udagawa, N.; Nishihara, T.; Takahashi, N.; Suda, T. Osteoclast differentiation factor acts as a multifunctional regulator in murine osteoclast differentiation and function. J. Immunol. 1999, 163, 434–442. [Google Scholar] [PubMed]
- Kim, H.J.; Zhao, H.; Kitaura, H.; Bhattacharyya, S.; Brewer, J.A.; Muglia, L.J.; Ross, F.P.; Teitelbaum, S.L. Glucocorticoids suppress bone formation via the osteoclast. J. Clin. Invest 2006, 116, 2152–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannoni, E.; Buricchi, F.; Raugei, G.; Ramponi, G.; Chiarugi, P. Intracellular reactive oxygen species activate Src tyrosine kinase during cell adhesion and anchorage-dependent cell growth. Mol. Cell. Biol. 2005, 25, 6391–6403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitelbaum, S.L. The osteoclast and its unique cytoskeleton. Ann. N. Y. Acad. Sci. 2011, 1240, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Shalev, M.; Elson, A. The roles of protein tyrosine phosphatases in bone-resorbing osteoclasts. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 114–123. [Google Scholar] [CrossRef]
- Ke, K.; Sul, O.J.; Choi, E.K.; Safdar, A.M.; Kim, E.S.; Choi, H.S. Reactive oxygen species induce the association of SHP-1 with -Src and the oxidation of both to enhance osteoclast survival. Am. J. Physiol. Endocrinol. Metab. 2014, 307, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Ye, L.; Shen, S.; Qian, T.; Pan, Y.; Jiang, Y.; Lin, J.; Liu, C.; Wu, Y.; Wang, X.; et al. Morin attenuates osteoclast formation and function by suppressing the NF-κB, MAPK and calcium signalling pathways. Phytother. Res. 2021, 35, 5694–5707. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y.; Lu, L.; Ma, Q.; Zhang, J. Preparation, characterization and systemic application of self-assembled hydroxyethyl starch nanoparticles-loaded flavonoid Morin for hyperuricemia therapy. Int. J. Nanomed. 2018, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Faccico, R.; Takeshita, S.; Zallone, A.; Ross, F.P.; Teitelbaum, S.L. c-Fms and the alphavbeta3 integrin collaborate during osteoclast differentiation. J. Clin. Invest 2003, 111, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Celik, H.; Kucukler, S.; Comaklı, S.; Özdemir, S.; Caglayan, C.; Yardım, A.; Kandemir, F.M. Morin attenuates ifosfamide-induced neurotoxicity in rats via suppression of oxidative stress, neuroinflammation and neuronal apoptosis. Neurotoxicology 2020, 76, 126–137. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Vehicle | Morin Only | LPS | LPS + Morin |
|---|---|---|---|---|
| BMD (mg/cm3) | 196.7 ± 8.47 | 189.0 ± 12.86 | 150.1 ± 7.33 a′ | 213.5 ± 11.39 b′′ |
| BV/TV (%) | 16.18 ± 0.86 | 15.96 ± 1.21 | 12.07 ± 0.58 a′ | 18.48 ± 1.21 b′′ |
| Tb.Th (μm) | 70.45 ± 3.69 | 68.41 ± 5.04 | 57.88 ± 2.42 a | 77.39 ± 3.16 b′′ |
| Tb.Sp (μm) | 348.5 ± 17.40 | 338.7 ± 12.19 | 450.9 ± 30.89 a | 342.5 ± 10.45 b′ |
| ALP (U/L) | 48.14 ± 1.00 | 45.97 ± 2.28 | 49.54 ± 1.33 | 48.45 ± 1.33 |
| OCN (ng/mL) | 25.90 ± 1.18 | 27.85 ± 1.48 | 27.43 ± 0.62 | 27.32 ± 1.95 |
| CTX-1 (ng/mL) | 25.55 ± 1.72 | 24.98 ± 1.41 | 48.67 ± 1.81 a′′ | 37.18 ± 2.70 b′ |
| MCP-1 (pg/mL) | 143.6 ± 16.59 | 140.1 ± 20.55 | 295.9 ± 13.91 a′′ | 209.3 ± 11.66 b′′ |
| H2O2 (μM) | 42.69 ± 2.50 | 42.08 ± 2.37 | 50.31 ± 1.87 a | 42.36 ± 1.09 b′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-J.; Park, J.-N.; Yoon, S.-Y.; Yu, R.; Suh, J.-H.; Choi, H.-S. Morin Disrupts Cytoskeleton Reorganization in Osteoclasts through an ROS/SHP1/c-Src Axis and Grants Protection from LPS-Induced Bone Loss. Antioxidants 2022, 11, 963. https://doi.org/10.3390/antiox11050963
Park H-J, Park J-N, Yoon S-Y, Yu R, Suh J-H, Choi H-S. Morin Disrupts Cytoskeleton Reorganization in Osteoclasts through an ROS/SHP1/c-Src Axis and Grants Protection from LPS-Induced Bone Loss. Antioxidants. 2022; 11(5):963. https://doi.org/10.3390/antiox11050963
Chicago/Turabian StylePark, Hyun-Jung, Jung-Nam Park, Sun-Young Yoon, Rina Yu, Jae-Hee Suh, and Hye-Seon Choi. 2022. "Morin Disrupts Cytoskeleton Reorganization in Osteoclasts through an ROS/SHP1/c-Src Axis and Grants Protection from LPS-Induced Bone Loss" Antioxidants 11, no. 5: 963. https://doi.org/10.3390/antiox11050963
APA StylePark, H. -J., Park, J. -N., Yoon, S. -Y., Yu, R., Suh, J. -H., & Choi, H. -S. (2022). Morin Disrupts Cytoskeleton Reorganization in Osteoclasts through an ROS/SHP1/c-Src Axis and Grants Protection from LPS-Induced Bone Loss. Antioxidants, 11(5), 963. https://doi.org/10.3390/antiox11050963