
Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions, and Reagents
2.2. Sample Preparations for iSS Detection
2.3. The Optimized iSS Detection Method
2.4. The Effect of Growth Conditions and Reagents on E. coli iSS
3. Results
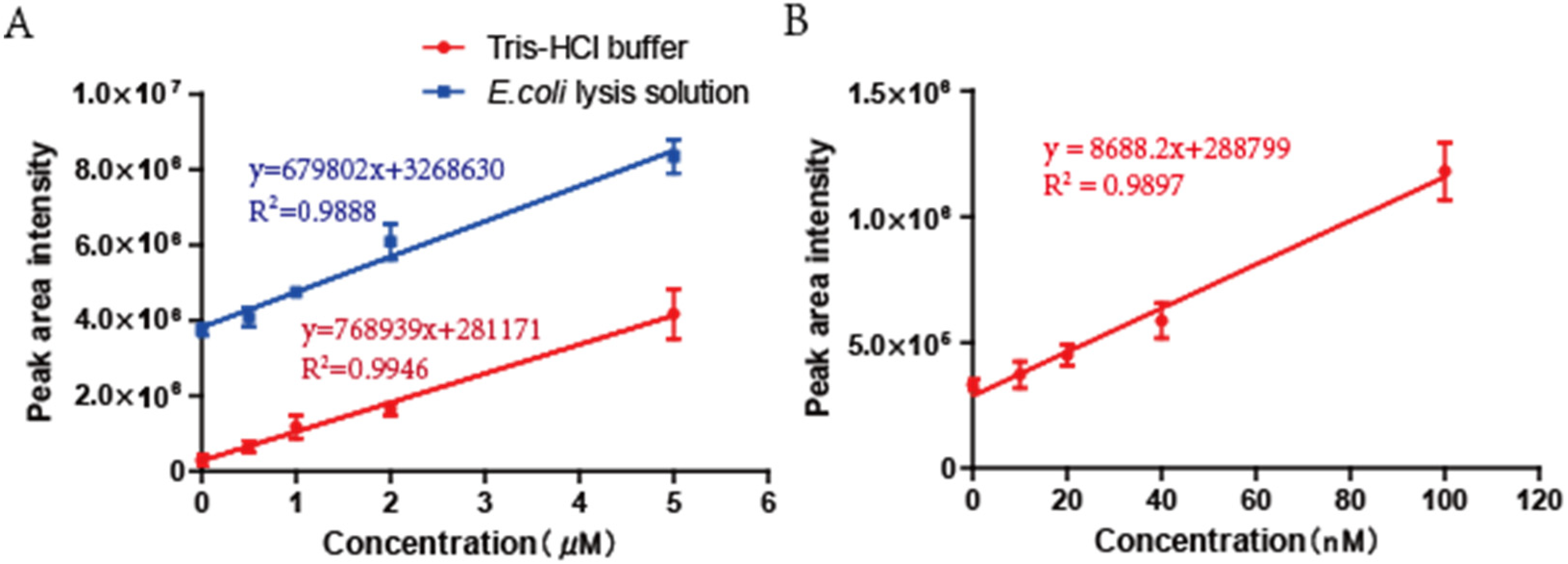
3.1. The Optimization of Bacterial Samples Preparation
3.2. The Effects of Different Stress Factors on iSS Content of E. coli
3.3. Effects of Cellular Redox Balance on E. coli iSS
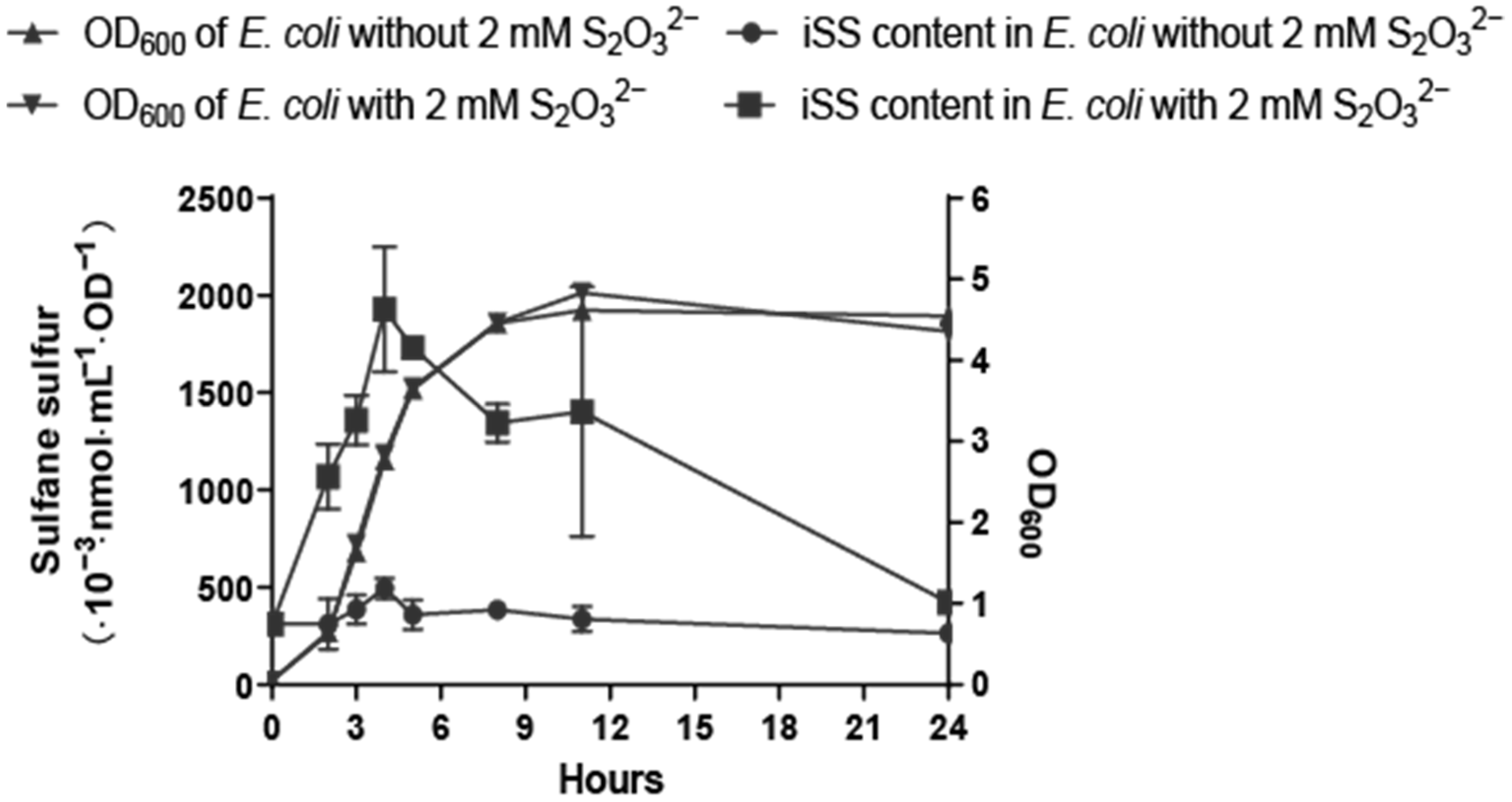
3.4. Effects of Exogenous Sulfur-Bearing Compounds on E. coli iSS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mishanina, A.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Shibuya, N.; Kimura, Y. Hydrogen Sulfide Is a Signaling Molecule and a Cytoprotectant. Antioxid. Redox Signal. 2012, 17, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 377–461. [Google Scholar] [CrossRef]
- Kimura, H. Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years. Biomolecules 2021, 11, 896. [Google Scholar] [CrossRef]
- Nagy, P.; Doka, E.; Ida, T.; Akaike, T. Measuring Reactive Sulfur Species and Thiol Oxidation States: Challenges and Cautions in Relation to Alkylation-Based Protocols. Antioxid. Redox Signal. 2020, 33, 1174–1189. [Google Scholar] [CrossRef]
- Castelblanco, M.; Nasi, S.; Pasch, A.; So, A.; Busso, N. The role of the gasotransmitter hydrogen sulfide in pathological calcification. Brit. J. Pharmacol. 2020, 177, 778–792. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, A.P.; Wei, W.L.; Gauld, J.W.; Auld, J.; Ozcan, F.; Aslan, M.; Mutus, B. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is inactivated by S-sulfuration in vitro. Free Radic. Bio. Med. 2015, 89, 512–521. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, Y.X.; Palmer, L.D.; Kehl-Fie, T.E.; Skaar, E.P.; Trinidad, J.C.; Giedroc, D.P. Hydrogen Sulfide and Reactive Sulfur Species Impact Proteome S-Sulfhydration and Global Virulence Regulation in Staphylococcus aureus. ACS Infect. Dis. 2017, 3, 744–755. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.T.; Gazi, S.K.; Barrow, R.K.; Yang, G.D.; Wang, R.; Snyder, S.H. H2S Signals Through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [Green Version]
- Xuan, G.; Lü, C.; Xu, H.; Chen, Z.; Li, K.; Liu, H.; Liu, H.; Xia, Y.; Xun, L. Sulfane Sulfur is an intrinsic signal activating MexR-regulated antibiotic resistance in Pseudomonas aeruginosa. Mol. Microbiol. 2020, 114, 1038–1048. [Google Scholar] [CrossRef]
- Xuan, G.; Lv, C.; Xu, H.; Li, K.; Liu, H.; Xia, Y.; Xun, L. Sulfane Sulfur Regulates LasR-Mediated Quorum Sensing and Virulence in Pseudomonas aeruginosa PAO1. Antioxidants 2021, 10, 1498. [Google Scholar] [CrossRef]
- Xu, H.; Xuan, G.; Liu, H.; Xia, Y.; Xun, L. Sulfane Sulfur Is a Strong Inducer of the Multiple Antibiotic Resistance Regulator MarR in Escherichia coli. Antioxidants 2021, 10, 1778. [Google Scholar] [CrossRef]
- Hou, N.K.; Yan, Z.Z.; Fan, K.L.; Li, H.J.; Zhao, R.; Xia, Y.Z.; Xun, L.Y.; Liu, H.W. OxyR senses sulfane sulfur and activates the genes for its removal in Escherichia coli. Redox Biol. 2019, 26, 101293. [Google Scholar] [CrossRef]
- Ni, X.; Kelly, S.S.; Xu, S.; Xian, M. The Path to Controlled Delivery of Reactive Sulfur Species. Acc. Chem. Res. 2021, 54, 3968–3978. [Google Scholar] [CrossRef]
- Xu, S.; Hamsath, A.; Neill, D.L.; Wang, Y.Y.; Yang, C.T.; Xian, M. Strategies for the Design of Donors and Precursors of Reactive Sulfur Species. Chem. Eur. J. 2019, 25, 4005–4016. [Google Scholar] [CrossRef] [PubMed]
- Gregory, G.J.; Boyd, E.F. Stressed out: Bacterial response to high salinity using compatible solute biosynthesis and uptake systems, lessons from Vibrionaceae. Comput. Struct. Biotec. 2021, 19, 1014–1027. [Google Scholar] [CrossRef]
- Fang, F.C.; Frawley, E.R.; Tapscott, T.; Vazquez-Torres, A. Bacterial Stress Responses during Host Infection. Cell Host Microbe 2016, 20, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reniere, M.L. Reduce, Induce, Thrive: Bacterial Redox Sensing during Pathogenesis. J. Bacteriol. 2018, 200, e00128-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Imlay, J.A. When anaerobes encounter oxygen: Mechanisms of oxygen toxicity, tolerance and defence. Nat. Rev. Microbiol. 2021, 19, 774–785. [Google Scholar] [CrossRef]
- Khademian, M.; Imlay, J.A. How Microbes Evolved to Tolerate Oxygen. Trends Microbiol. 2021, 29, 428–440. [Google Scholar] [CrossRef]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a Healthy Proteome during Oxidative Stress. Mol. Cell 2018, 69, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabel, M.; Nackenoff, A.; Kirsch, W.M.; Harrison, F.E.; Perry, G.; Schrag, M. Markers of oxidative damage to lipids, nucleic acids and proteins and antioxidant enzymes activities in Alzheimer’s disease brain: A meta-analysis in human pathological specimens. Free Radic. Biol. Med. 2018, 115, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, K.; Dec, K.; Kaldunska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species—Sources, functions, oxidative damage. Pol. Merkur Lek. 2020, 48, 124–127. [Google Scholar]
- Ashby, L.V.; Springer, R.; Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Evaluating the bactericidal action of hypochlorous acid in culture media. Free Radic. Biol. Med. 2020, 159, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Calleja, L.F.; Yoval-Sanchez, B.; Hernandez-Esquivel, L.; Gallardo-Perez, J.C.; Sosa-Garrocho, M.; Marin-Hernandez, A.; Jasso-Chavez, R.; Macias-Silva, M.; Rodriguez-Zavala, J.S. Activation of ALDH1A1 by omeprazole reduces cell oxidative stress damage. FEBS J. 2021, 288, 4064–4080. [Google Scholar] [CrossRef] [PubMed]
- Corona, F.; Martinez, J.L.; Nikel, P.I. The global regulator Crc orchestrates the metabolic robustness underlying oxidative stress resistance in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 898–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Simplicio, P.; Lupis, E.; Rossi, R. Different mechanisms of formation of glutathione-protein mixed disulfides of diamide and tert-butyl hydroperoxide in rat blood. Biochim. Biophys. Acta 1996, 1289, 252–260. [Google Scholar] [CrossRef]
- Li, K.; Xin, Y.; Xuan, G.; Zhao, R.; Liu, H.; Xia, Y.; Xun, L. Escherichia coli Uses Separate Enzymes to Produce H2S and Reactive Sulfane Sulfur From L-cysteine. Front. Microbiol. 2019, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Pester, M.; Knorr, K.H.; Friedrich, M.W.; Wagner, M.; Loy, A. Sulfate-reducing microorganisms in wetlands—Fameless actors in carbon cycling and climate change. Front. Microbiol. 2012, 3, 72. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R. H2S and polysulfide metabolism: Conventional and unconventional pathways. Biochem. Pharmacol. 2018, 149, 77–90. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, Y.; Chen, Z.; Xia, Y.; Xun, L.; Liu, H. Sulfide-quinone oxidoreductase is required for cysteine synthesis and indispensable to mitochondrial health. Redox Biol. 2021, 47, 102169. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.H.; Jen, L.N.; Su, H.Y.; Lii, C.K.; Sheen, L.Y.; Liu, C.T. Effects of garlic oil and two of its major organosulfur compounds, diallyl disulfide and diallyl trisulfide, on intestinal damage in rats injected with endotoxin. Toxicol. Appl. Pharm. 2006, 213, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.; Wang, T.; Shao, M.; Chen, Z.; Liu, H.; Xia, Y.; Xun, L. Sensitive Method for Reliable Quantification of Sulfane Sulfur in Biological Samples. Anal. Chem. 2019, 91, 11981–11986. [Google Scholar] [CrossRef] [PubMed]
- Kruithof, P.D.; Lunev, S.; Lozano, S.P.A.; Batista, F.D.; Al-dahmani, Z.M.; Joles, J.A.; Dolga, A.M.; Groves, M.R.; van Goor, H. Unraveling the role of thiosulfate sulfurtransferase in metabolic diseases. BBA Mol. Basis Dis. 2020, 1866, 165716. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xia, Y.; Liu, H.; Liu, H.; Xun, L. The Mechanisms of Thiosulfate Toxicity against Saccharomyces cerevisiae. Antioxidants 2021, 10, 646. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Cao, Q.; Pang, X.; Xia, Y.; Xun, L.; Liu, H. Sulfane sulfur-activated actinorhodin production and sporulation is maintained by a natural gene circuit in Streptomyces coelicolor. Microb. Biotechnol. 2020, 13, 1917–1932. [Google Scholar] [CrossRef]
- Müller, N.; Rauhut, D.; Tarasov, A. Sulfane Sulfur Compounds as Source of Reappearance of Reductive Off-Odors in Wine. Fermentation 2022, 8, 53. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Redox Biochemistry of Hydrogen Sulfide. J. Biol. Chem. 2010, 285, 21903–21907. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Choi, S.E.; Koh, H.C. PGAM5 regulates PINK1/Parkin-mediated mitophagy via DRP1 in CCCP-induced mitochondrial dysfunction. Toxicol. Lett. 2018, 284, 120–128. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.; Zhang, X.; Xin, Y.; Xia, Y.; Xun, L.; Liu, H. Rhodanese Rdl2 produces reactive sulfur species to protect mitochondria from reactive oxygen species. Free Radic. Biol Med. 2021, 177, 287–298. [Google Scholar] [CrossRef]
- Huang, B.; Zhao, Z.; Huang, C.; Zhao, M.; Zhang, Y.; Liu, Y.; Liao, X.; Huang, S.; Zhao, Y. Role of metal cations and oxyanions in the regulation of protein arginine phosphatase activity of Yw1E from Bacillus subtilis. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129698. [Google Scholar] [CrossRef]
- Panmanee, W.; Charoenlap, N.; Atichartpongkul, S.; Mahavihakanont, A.; Whiteside, M.D.; Winsor, G.; Brinkman, F.S.L.; Mongkolsuk, S.; Hassett, D.J. The OxyR-regulated phnW gene encoding 2-aminoethylphosphonate: Pyruvate aminotransferase helps protect Pseudomonas aeruginosa from tert-butyl hydroperoxide. PLoS ONE 2017, 12, e0189066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imlay, J.A. Where in the world do bacteria experience oxidative stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.W.; Schwessinger, E.A.; Jakob, U.; Gray, M.J. The RclR Protein Is a Reactive Chlorine-specific Transcription Factor in Escherichia coli. J. Biol. Chem. 2013, 288, 32574–32584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakovic, J.; Yu, B.Y.K.; Silva, D.; Baczynska, M.; Peak-Chew, S.Y.; Switzer, A.; Burchell, L.; Wigneshweraraj, S.; Vandanashree, M.; Gopal, B.; et al. Redox Regulation of the Quorum-sensing Transcription Factor AgrA by Coenzyme, A. Antioxidants 2021, 10, 841. [Google Scholar] [CrossRef] [PubMed]
- Spear, N.; Aust, S.D. Effects of glutathione on Fenton reagent-dependent radical production and DNA oxidation. Arch. Biochem. Biophys. 1995, 324, 111–116. [Google Scholar] [CrossRef]
- Miyamoto, R.; Koike, S.; Takano, Y.; Shibuya, N.; Kimura, Y.; Hanaoka, K.; Urano, Y.; Ogasawara, Y.; Kimura, H. Polysulfides (H2Sn) produced from the interaction of hydrogen sulfide (H2S) and nitric oxide (NO) activate TRPA1 channels. Sci. Rep. 2017, 7, 45995. [Google Scholar] [CrossRef] [Green Version]
- Stevens, R.; Stevens, L.; Price, N.C. The Stabilities of Various Thiol Compounds Used in Protein Purifications. Biochem. Educ. 1983, 11, 70. [Google Scholar] [CrossRef]
- Olson, K.R. Hydrogen sulfide, reactive sulfur species and coping with reactive oxygen species. Free Radical Bio. Med. 2019, 140, 74–83. [Google Scholar] [CrossRef]
- Hancock, J.T.; Whiteman, M. Hydrogen sulfide signaling: Interactions with nitric oxide and reactive oxygen species. Ann. N. Y. Acad. Sci. 2016, 1365, 5–14. [Google Scholar] [CrossRef]
- Fukuto, J.M.; Ignarro, L.J.; Nagy, P.; Wink, D.A.; Kevil, C.G.; Feelisch, M.; Cortese-Krott, M.M.; Bianco, C.L.; Kumagai, Y.; Hobbs, A.J.; et al. Biological hydropersulfides and related polysulfides—A new concept and perspective in redox biology. FEBS Lett. 2018, 592, 2140–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Predmore, B.L.; Lefer, D.J.; Gojon, G. Hydrogen Sulfide in Biochemistry and Medicine. Antioxid. Redox Signal. 2012, 17, 119–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iciek, M.; Bilska-Wilkosz, A.; Gorny, M. Sulfane sulfur—New findings on an old topic. Acta Biochim. Pol. 2019, 66, 533–544. [Google Scholar] [CrossRef]
- Bruska, M.K.; Stiebritz, M.T.; Reiher, M. Binding of Reactive Oxygen Species at Fe-S Cubane Clusters. Chem. A Eur. J. 2015, 21, 19081–19089. [Google Scholar] [CrossRef]
- Lu, Z.; Imlay, J.A. A conserved motif liganding the [4Fe-4S] cluster in [4Fe-4S] fumarases prevents irreversible inactivation of the enzyme during hydrogen peroxide stress. Redox Biol. 2019, 26, 101296. [Google Scholar] [CrossRef]
- Rydz, L.; Wrobel, M.; Jurkowska, H. Sulfur Administration in Fe-S Cluster Homeostasis. Antioxidants 2021, 10, 1738. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.S.; Olson, K.R.; Zhu, Y.C. Hydrogen Sulfide: Biogenesis, Physiology, and Pathology. Oxid. Med. Cell. Longev. 2016, 2016, 6549625. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Imlay, J. Why do bacteria use so many enzymes to scavenge hydrogen peroxide? Arch. Biochem. Biophys 2012, 525, 145–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, D.H.; Tuminello, J.F.; Emptage, M.H. The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J. Biol. Chem. 1993, 268, 22369–22376. [Google Scholar] [CrossRef]
- Gray, M.J.; Wholey, W.Y.; Jakob, U. Bacterial Responses to Reactive Chlorine Species. Annu. Rev. Microbiol. 2013, 67, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Sevilla, E.; Bes, M.T.; Gonzalez, A.; Peleato, M.L.; Fillat, M.F. Redox-Based Transcriptional Regulation in Prokaryotes: Revisiting Model Mechanisms. Antioxid. Redox Signal. 2019, 30, 1651–1696. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.G.; Lee, B.J. How Bacterial Redox Sensors Transmit Redox Signals via Structural Changes. Antioxidants 2021, 10, 502. [Google Scholar] [CrossRef]
- Unden, G.; Bongaerts, J. Alternative respiratory pathways of Escherichia coli: Energetics and transcriptional regulation in response to electron acceptors. BBA Bioenergetics 1997, 1320, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Kawano, Y.; Onishi, F.; Shiroyama, M.; Miura, M.; Tanaka, N.; Oshiro, S.; Nonaka, G.; Nakanishi, T.; Ohtsu, I. Improved fermentative L-cysteine overproduction by enhancing a newly identified thiosulfate assimilation pathway in Escherichia coli. Appl. Microbiol. Biot. 2017, 101, 6879–6889. [Google Scholar] [CrossRef]
- Sirko, A.; Hryniewicz, M.; Hulanicka, D.; Bock, A. Sulfate and Thiosulfate Transport in Escherichia-Coli K-12—Nucleotide-Sequence and Expression of the Cystwam Gene-Cluster. J. Bacteriol. 1990, 172, 3351–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Donahue, J.L.; Battle, S.E.; Ray, W.K.; Larson, T.J. Biochemical and Genetic Characterization of PspE and GlpE, Two Single-domain Sulfurtransferases of Escherichia coli. Open Microbiol. J. 2008, 2, 18–28. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Q.; Ran, M.; Yang, Y.; Liu, H.; Xun, L.; Xia, Y. Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli. Antioxidants 2022, 11, 1292. https://doi.org/10.3390/antiox11071292
Yu Q, Ran M, Yang Y, Liu H, Xun L, Xia Y. Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli. Antioxidants. 2022; 11(7):1292. https://doi.org/10.3390/antiox11071292
Chicago/Turabian StyleYu, Qiaoli, Mingxue Ran, Yuqing Yang, Huaiwei Liu, Luying Xun, and Yongzhen Xia. 2022. "Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli" Antioxidants 11, no. 7: 1292. https://doi.org/10.3390/antiox11071292
APA StyleYu, Q., Ran, M., Yang, Y., Liu, H., Xun, L., & Xia, Y. (2022). Optimization of a Method for Detecting Intracellular Sulfane Sulfur Levels and Evaluation of Reagents That Affect the Levels in Escherichia coli. Antioxidants, 11(7), 1292. https://doi.org/10.3390/antiox11071292