
Addition of oh8dG to Cardioplegia Attenuated Myocardial Oxidative Injury through the Inhibition of Sodium Bicarbonate Cotransporter Activity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
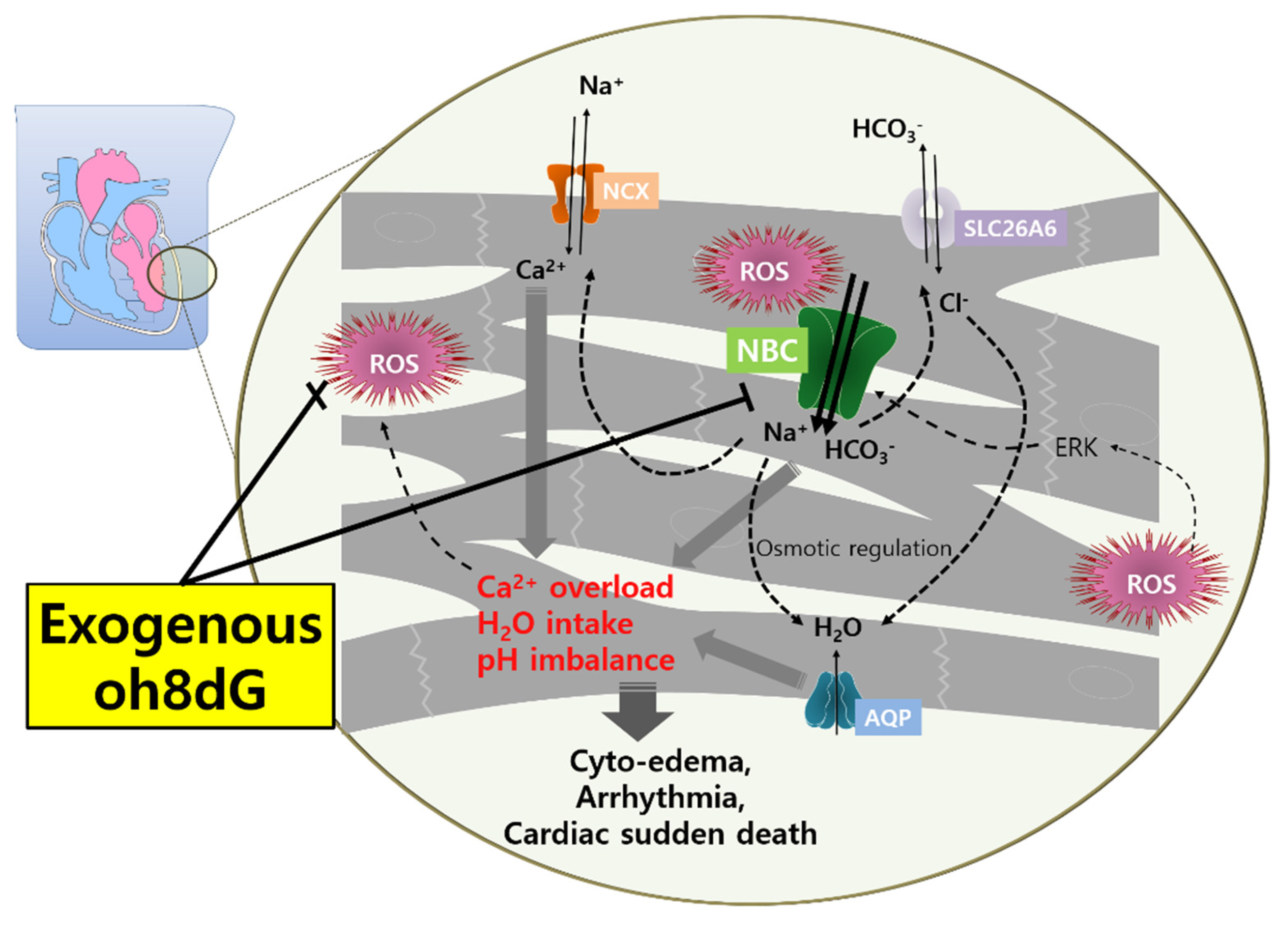
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cardioplegia Infusion and Cardiac Strip Preparation
2.3. Isolation of Single Cardiomyocytes
2.4. ROS Staining
2.5. Cell Culture and DNA Transfection
2.6. Measurement of Na+-HCO3− Cotransporter (NBC) Activity
2.7. Western Blotting
2.8. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.9. Transferase (TdT)-Mediated dUTP Nick End Labeling (TUNEL) Assay
2.10. Immunostaining
2.11. MTT Assay
2.12. Statistical Analysis
3. Results
3.1. Treatment of oh8dG Attenuated HTK-Mediated ROS Level in Cardiac Strips and Isolated Cardiomyocytes
3.2. The Treatment of oh8dG Attenuated ROS-Mediated NBC Activities
3.3. HTK-Exposed Cardiac Tissues Enhanced NBC Activity, Attenuated by the Treatment of oh8dG
3.4. Treatment of oh8dG Attenuated DNA Fragmentation and Expression of the Apoptotic Marker in HTK-Exposed Cardiac Tissues
3.5. Treatment of oh8dG Attenuated HTK-Mediated Cardiac Destructive Signals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, M.J.; Baicu, S.C. Current state of hypothermic machine perfusion preservation of organs: The clinical perspective. Cryobiology 2010, 60, S20–S35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, M.G.; Soares, A.F.; Carvalho, R.A.; Oliveira, P.J. Sodium hydrosulfide improves the protective potential of the cardioplegic histidine buffer solution. Eur. J. Pharmacol. 2011, 654, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Gomez, L.; Li, B.; Mewton, N.; Sanchez, I.; Piot, C.; Elbaz, M.; Ovize, M. Inhibition of mitochondrial permeability transition pore opening: Translation to patients. Cardiovasc. Res. 2009, 83, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Dorado, D.; Ruiz-Meana, M.; Inserte, J.; Rodriguez-Sinovas, A.; Piper, H.M. Calcium-mediated cell death during myocardial reperfusion. Cardiovasc. Res. 2012, 94, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Sanada, S.; Komuro, I.; Kitakaze, M. Pathophysiology of myocardial reperfusion injury: Preconditioning, postconditioning, and translational aspects of protective measures. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1723–H1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.W.; Sun, X.D.; Guo, L.; Su, C.H.; Sun, X.J.; Xu, Z.Y. Hydrogen as additive of HTK solution fortifies myocardial preservation in grafts with prolonged cold ischemia. Int. J. Cardiol. 2013, 167, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Fridell, J.A.; Mangus, R.S.; Tector, A.J. Clinical experience with histidine-tryptophan-ketoglutarate solution in abdominal organ preservation: A review of recent literature. Clin. Transplant. 2009, 23, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Latchana, N.; Peck, J.R.; Whitson, B.A.; Henry, M.L.; Elkhammas, E.A.; Black, S.M. Preservation solutions used during abdominal transplantation: Current status and outcomes. World J. Transplant. 2015, 5, 154–164. [Google Scholar] [CrossRef]
- Janssen, H.; Janssen, P.H.; Broelsch, C.E. Celsior solution compared with University of Wisconsin solution (UW) and histidine-tryptophan-ketoglutarate solution (HTK) in the protection of human hepatocytes against ischemia-reperfusion injury. Transpl. Int. 2003, 16, 515–522. [Google Scholar] [CrossRef]
- Semmelmann, A.; Neeff, H.; Sommer, O.; Thomusch, O.; Hopt, U.T.; von Dobschuetz, E. Evaluation of preservation solutions by ESR-spectroscopy: Superior effects of University of Wisconsin over Histidine-Tryptophan-Ketoglutarate in reducing renal reactive oxygen species. Kidney Int. 2007, 71, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Rauen, U.; Klempt, S.; de Groot, H. Histidine-induced injury to cultured liver cells, effects of histidine derivatives and of iron chelators. Cell. Mol. Life Sci. 2007, 64, 192–205. [Google Scholar] [CrossRef]
- Lee, E.H.; Park, S.R.; Paik, K.S.; Suh, C.K. Intracellular acidosis decreases the outward Na+-Ca2+ exchange current in guinea pig ventricular myocytes. Yonsei Med. J. 1995, 36, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Inserte, J.; Garcia-Dorado, D.; Ruiz-Meana, M.; Padilla, F.; Barrabes, J.A.; Pina, P.; Agullo, L.; Piper, H.M.; Soler-Soler, J. Effect of inhibition of Na+/Ca2+ exchanger at the time of myocardial reperfusion on hypercontracture and cell death. Cardiovasc. Res. 2002, 55, 739–748. [Google Scholar] [CrossRef] [Green Version]
- White, C.W.; Messer, S.J.; Large, S.R.; Conway, J.; Kim, D.H.; Kutsogiannis, D.J.; Nagendran, J.; Freed, D.H. Transplantation of Hearts Donated after Circulatory Death. Front. Cardiovasc. Med. 2018, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Leem, C.H.; Lagadic-Gossmann, D.; Vaughan-Jones, R.D. Characterization of intracellular pH regulation in the guinea-pig ventricular myocyte. J. Physiol. 1999, 517, 159–180. [Google Scholar] [CrossRef]
- Khandoudi, N.; Albadine, J.; Robert, P.; Krief, S.; Berrebi-Bertrand, I.; Martin, X.; Bevensee, M.O.; Boron, W.F.; Bril, A. Inhibition of the cardiac electrogenic sodium bicarbonate cotransporter reduces ischemic injury. Cardiovasc. Res. 2001, 52, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Kohout, T.A.; Rogers, T.B. Angiotensin-Ii Activates the Na+/HCO3− Symport through a Phosphoinositide-Independent Mechanism in Cardiac-Cells. J. Biol. Chem. 1995, 270, 20432–20438. [Google Scholar] [CrossRef] [Green Version]
- De Giusti, V.C.; Garciarena, C.D.; Aiello, E.A. Role of reactive oxygen species (ROS) in angiotensin II-induced stimulation of the cardiac Na+/HCO3− cotransport. J. Mol. Cell. Cardiol. 2009, 47, 716–722. [Google Scholar] [CrossRef]
- De Giusti, V.C.; Orlowski, A.; Aiello, E.A. Angiotensin II inhibits the electrogenic Na+/HCO3− cotransport of cat cardiac myocytes. J. Mol. Cell. Cardiol. 2010, 49, 812–818. [Google Scholar] [CrossRef]
- Ames, B.N. Endogenous oxidative DNA damage, aging, and cancer. Free Radic. Res. Commun. 1989, 7, 121–128. [Google Scholar] [CrossRef]
- Choi, S.; Choi, H.H.; Lee, S.H.; Ko, S.H.; You, H.J.; Ye, S.K.; Chung, M.H. Anti-inflammatory effects of 8-hydroxy-2′-deoxyguanosine on lipopolysaccharide-induced inflammation via Rac suppression in Balb/c mice. Free Radic. Biol. Med. 2007, 43, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Cho, I.H.; Kim, H.S.; Jung, J.E.; Kim, J.E.; Lee, K.H.; Park, T.; Yang, Y.M.; Seong, S.Y.; Ye, S.K.; et al. Anti-inflammatory effects of 8-hydroxydeoxyguanosine in LPS-induced microglia activation: Suppression of STAT3-mediated intercellular adhesion molecule-1 expression. Exp. Mol. Med. 2006, 38, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Taek Han, S.; Choi, S.W.; Sung, S.Y.; You, H.J.; Ye, S.K.; Chung, M.H. Inhibition of Rac and Rac-linked functions by 8-oxo-2′-deoxyguanosine in murine macrophages. Free Radic. Res. 2009, 43, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Ko, S.H.; Ye, S.K.; Chung, M.H. 8-Oxo-2′-deoxyguanosine ameliorates UVB-induced skin damage in hairless mice by scavenging reactive oxygen species and inhibiting MMP expression. J. Dermatol. Sci. 2013, 70, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Kroese, L.J.; Scheffer, P.G. 8-hydroxy-2′-deoxyguanosine and cardiovascular disease: A systematic review. Curr. Atheroscler. Rep. 2014, 16, 452. [Google Scholar] [CrossRef]
- Shin, S.K.; Kim, K.O.; Kim, S.H.; Kwon, O.S.; Choi, C.S.; Jeong, S.H.; Kim, Y.S.; Kim, J.H.; Chung, M.H. Exogenous 8-hydroxydeoxyguanosine ameliorates liver fibrosis through the inhibition of Rac1-NADPH oxidase signaling. J. Gastroenterol. Hepatol. 2020, 35, 1078–1087. [Google Scholar] [CrossRef]
- Urbaniak, S.K.; Boguszewska, K.; Szewczuk, M.; Kazmierczak-Baranska, J.; Karwowski, B.T. 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development. Molecules 2020, 25, 202. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Im, S.T.; Yoon, J.Y.; Kim, S.; Kim, M.K.; Chung, M.H.; Park, C.K. Comparison of therapeutic effects between topical 8-oxo-2′-deoxyguanosine and corticosteroid in ocular alkali burn model. Sci. Rep. 2021, 11, 6909. [Google Scholar] [CrossRef]
- Kim, H.S.; Ye, S.K.; Cho, I.H.; Jung, J.E.; Kim, D.H.; Choi, S.; Kim, Y.S.; Park, C.G.; Kim, T.Y.; Lee, J.W.; et al. 8-hydroxydeoxyguanosine suppresses NO production and COX-2 activity via Rac1/STATs signaling in LPS-induced brain microglia. Free Radical. Bio. Med. 2006, 41, 1392–1403. [Google Scholar] [CrossRef]
- Ro, J.Y.; Kim, D.Y.; Lee, S.H.; Park, J.W.; Chung, M.H. Effects of 7,8-dihydro-8-oxo-deoxyguanosine on antigen challenge in ovalbumin-sensitized mice may be mediated by suppression of Rac. Br. J. Pharmacol. 2009, 158, 1743–1752. [Google Scholar] [CrossRef] [Green Version]
- Hajas, G.; Bacsi, A.; Aguilera-Aguirre, L.; Hegde, M.L.; Tapas, K.H.; Sur, S.; Radak, Z.; Ba, X.; Boldogh, I. 8-Oxoguanine DNA glycosylase-1 links DNA repair to cellular signaling via the activation of the small GTPase Rac1. Free Radic. Biol. Med. 2013, 61, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernikov, A.V.; Gudkov, S.V.; Usacheva, A.M.; Bruskov, V.I. Exogenous 8-Oxo-7,8-dihydro-2′-deoxyguanosine: Biomedical Properties, Mechanisms of Action, and Therapeutic Potential. Biochemistry 2017, 82, 1686–1701. [Google Scholar] [CrossRef] [PubMed]
- Im, S.T.; Kim, H.Y.; Yoon, J.Y.; Oh, J.Y.; Kim, M.K.; Chung, M.H.; Paik, H.J.; Kim, D.H. Therapeutic Effects of Topical 8-Oxo-2′-deoxyguanosine on Ethanol-Induced Ocular Chemical Injury Models. Cornea 2018, 37, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Ock, C.Y.; Hong, K.S.; Choi, K.S.; Chung, M.H.; Kim, Y.; Kim, J.H.; Hahm, K.B. A novel approach for stress-induced gastritis based on paradoxical anti-oxidative and anti-inflammatory action of exogenous 8-hydroxydeoxyguanosine. Biochem. Pharmacol. 2011, 81, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.H.; Lee, J.K.; Lee, H.J.; Ye, S.K.; Kim, H.S.; Chung, M.H. 8-Oxo-2′-deoxyguanosine ameliorates features of metabolic syndrome in obese mice. Biochem. Biophys. Res. Commun. 2014, 443, 610–616. [Google Scholar] [CrossRef]
- Kim, J.E.; Hyun, J.W.; Hayakawa, H.; Choi, S.; Choi, J.; Chung, M.H. Exogenous 8-oxo-dG is not utilized for nucleotide synthesis but enhances the accumulation of 8-oxo-Gua in DNA through error-prone DNA synthesis. Mutat. Res. 2006, 596, 128–136. [Google Scholar] [CrossRef]
- Kim, J.E.; Chung, M.H. 8-Oxo-7,8-dihydro-2′-deoxyguanosine is not salvaged for DNA synthesis in human leukemic U937 cells. Free Radic. Res. 2006, 40, 461–466. [Google Scholar] [CrossRef]
- Ji, M.; In Lee, S.; Lee, S.A.; Son, K.H.; Hong, J.H. Enhanced Activity by NKCC1 and Slc26a6 Mediates Acidic pH and Cl− Movement after Cardioplegia-Induced Arrest of db/db Diabetic Heart. Mediat. Inflamm. 2019, 2019, 7583760. [Google Scholar] [CrossRef] [Green Version]
- Ackers-Johnson, M.; Li, P.Y.; Holmes, A.P.; O’Brien, S.M.; Pavlovic, D.; Foo, R.S. A Simplified, Langendorff-Free Method for Concomitant Isolation of Viable Cardiac Myocytes and Nonmyocytes From the Adult Mouse Heart. Circ. Res. 2016, 119, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Lee, H.S.; Park, M.K.; Hwang, E.S.; Park, E.M.; Kasai, H.; Chung, M.H. Identification of 8-Hydroxyguanine Glycosylase Activity in Mammalian-Tissues Using 8-Hydroxyguanine Specific Monoclonal-Antibody. Biochem. Biophys. Res. Commun. 1993, 196, 1545–1551. [Google Scholar] [CrossRef]
- Borradaile, N.M.; Han, X.; Harp, J.D.; Gale, S.E.; Ory, D.S.; Schaffer, J.E. Disruption of endoplasmic reticulum structure and integrity in lipotoxic cell death. J. Lipid Res. 2006, 47, 2726–2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.H.; Chen, T.P.; Wang, Y.C.; Lin, Y.M.; Fang, S.W. AMP-activated protein kinase activation during cardioplegia-induced hypoxia/reoxygenation injury attenuates cardiomyocytic apoptosis via reduction of endoplasmic reticulum stress. Mediat. Inflamm. 2010, 2010, 130636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Bianchi, C.; Li, J.; Sellke, F.W. Improved profile of bad phosphorylation and caspase 3 activation after blood versus crystalloid cardioplegia. Ann. Thorac. Surg. 2004, 77, 1384–1389; discussion 1389–1390. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Lee, J.W.; Kim, S.K.; Yu, W.J.; Lee, S.J.; Kim, D.; Kim, K.W.; Jung, J.W.; Hong, I.S. The impact of fine particulate matter (PM) on various beneficial functions of human endometrial stem cells through its key regulator SERPINB2. Exp. Mol. Med. 2021, 53, 1850–1865. [Google Scholar] [CrossRef]
- Chen, M.M.; Lam, A.; Abraham, J.A.; Schreiner, G.F.; Joly, A.H. CTGF expression is induced by TGF-beta in cardiac fibroblasts and cardiac myocytes: A potential role in heart fibrosis. J. Mol. Cell. Cardiol. 2000, 32, 1805–1819. [Google Scholar] [CrossRef]
- Accornero, F.; van Berlo, J.H.; Correll, R.N.; Elrod, J.W.; Sargent, M.A.; York, A.; Rabinowitz, J.E.; Leask, A.; Molkentin, J.D. Genetic Analysis of Connective Tissue Growth Factor as an Effector of Transforming Growth Factor beta Signaling and Cardiac Remodeling. Mol. Cell. Biol. 2015, 35, 2154–2164. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Kim, S.C.; Wang, Y.; Gupta, S.; Davis, B.; Simon, S.I.; Torre-Amione, G.; Knowlton, A.A. HSP60 in heart failure: Abnormal distribution and role in cardiac myocyte apoptosis. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2238–H2247. [Google Scholar] [CrossRef] [Green Version]
- Aredia, F.; Scovassi, A.I. Poly(ADP-ribose): A signaling molecule in different paradigms of cell death. Biochem. Pharmacol. 2014, 92, 157–163. [Google Scholar] [CrossRef]
- Ji, M.; Ryu, H.J.; Baek, H.M.; Shin, D.M.; Hong, J.H. Dynamic synovial fibroblasts are modulated by NBCn1 as a potential target in rheumatoid arthritis. Exp. Mol. Med. 2022, 54, 503–517. [Google Scholar] [CrossRef]
- McDonald, P.C.; Swayampakula, M.; Dedhar, S. Coordinated Regulation of Metabolic Transporters and Migration/Invasion by Carbonic Anhydrase IX. Metabolites 2018, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- De Giusti, V.C.; Ciancio, M.C.; Orlowski, A.; Aiello, E.A. Modulation of the cardiac sodium/bicarbonate cotransporter by the renin angiotensin aldosterone system: Pathophysiological consequences. Front. Physiol. 2013, 4, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zima, A.V.; Blatter, L.A. Redox regulation of cardiac calcium channels and transporters. Cardiovasc. Res. 2006, 71, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.W.; Yoon, S.H.; Yu, Y.; Han, C.S.; Park, J.S.; Kim, H.S.; Lee, S.J.; Lee, Y.S.; You, H.J.; Chung, M.H. Oh8dG induces G1 arrest in a human acute leukemia cell line by upregulating P21 and blocking the RAS to ERK signaling pathway. Int. J. Cancer 2006, 118, 302–309. [Google Scholar] [CrossRef]
- Eller-Borges, R.; Batista, W.L.; da Costa, P.E.; Tokikawa, R.; Curcio, M.F.; Strumillo, S.T.; Sartori, A.; Moraes, M.S.; de Oliveira, G.A.; Taha, M.O.; et al. Ras, Rac1, and phosphatidylinositol-3-kinase (PI3K) signaling in nitric oxide induced endothelial cell migration. Nitric Oxide 2015, 47, 40–51. [Google Scholar] [CrossRef]
- Yu, M.H.; Lin, M.C.; Huang, C.N.; Chan, K.C.; Wang, C.J. Acarbose inhibits the proliferation and migration of vascular smooth muscle cells via targeting Ras signaling. Vasc. Pharmacol. 2018, 103–105, 8–15. [Google Scholar] [CrossRef]
- Zhan, H.; Bhattacharya, S.; Cai, H.; Iglesias, P.A.; Huang, C.H.; Devreotes, P.N. An Excitable Ras/PI3K/ERK Signaling Network Controls Migration and Oncogenic Transformation in Epithelial Cells. Dev. Cell 2020, 54, 608–623.e5. [Google Scholar] [CrossRef]
- Pan, Y.H.; Chen, J.; Sun, C.; Ma, J.F.; Li, X. Effect of Ras-guanine nucleotide release factor 1-mediated H-Ras/ERK signaling pathway on glioma. Brain Res. 2021, 1754, 147247. [Google Scholar] [CrossRef]
- Boedtkjer, E.; Bentzon, J.F.; Dam, V.S.; Aalkjaer, C. Na+, HCO3−—Cotransporter NBCn1 increases pHi gradients, filopodia, and migration of smooth muscle cells and promotes arterial remodelling. Cardiovasc. Res. 2016, 111, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Ng, F.L.; Boedtkjer, E.; Witkowska, K.; Ren, M.; Zhang, R.; Tucker, A.; Aalkjaer, C.; Caulfield, M.J.; Ye, S. Increased NBCn1 expression, Na+/HCO3− co-transport and intracellular pH in human vascular smooth muscle cells with a risk allele for hypertension. Hum. Mol. Genet. 2017, 26, 989–1002. [Google Scholar] [CrossRef] [Green Version]
- Takago, S.; Matsumoto, I.; Kato, H.; Saito, N.; Ueda, H.; Iino, K.; Kimura, K.; Takemura, H. Hypothermic Preservation of Rat Hearts Using Antifreeze Glycoprotein. Physiol. Res. 2020, 69, 1029–1038. [Google Scholar] [CrossRef]
- Gu, K.; Kin, S.; Saitoh, Y.; Nosaka, S.; Sasaki, T.; Yamauchi, M.; Nakayama, K. Cardioprotective effect of nicorandil in histidine-tryptophan-ketoglurate solution during the cold storage of isolated hearts. Transplantation 1996, 61, 1572–1575. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, Y.; Hashimoto, M.; Ku, K.; Kin, S.; Nosaka, S.; Masumura, S.; Nakayama, K. Heart preservation in HTK solution: Role of coronary vasculature in recovery of cardiac function. Ann. Thorac. Surg. 2000, 69, 107–112. [Google Scholar] [CrossRef]
- Mohr, A.; Brockmann, J.G.; Becker, F. HTK-N: Modified Histidine-Tryptophan-Ketoglutarate Solution-A Promising New Tool in Solid Organ Preservation. Int. J. Mol. Sci. 2020, 21, 6468. [Google Scholar] [CrossRef]
- Penna, C.; Perrelli, M.G.; Pagliaro, P. Mitochondrial pathways, permeability transition pore, and redox signaling in cardioprotection: Therapeutic implications. Antioxid. Redox Signal. 2013, 18, 556–599. [Google Scholar] [CrossRef]
- Pagliaro, P.; Moro, F.; Tullio, F.; Perrelli, M.G.; Penna, C. Cardioprotective pathways during reperfusion: Focus on redox signaling and other modalities of cell signaling. Antioxid. Redox Signal. 2011, 14, 833–850. [Google Scholar] [CrossRef] [Green Version]
- Penna, C.; Mancardi, D.; Rastaldo, R.; Losano, G.; Pagliaro, P. Intermittent activation of bradykinin B2 receptors and mitochondrial KATP channels trigger cardiac postconditioning through redox signaling. Cardiovasc. Res. 2007, 75, 168–177. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, M.J.; Son, K.H.; Hong, J.H. Addition of oh8dG to Cardioplegia Attenuated Myocardial Oxidative Injury through the Inhibition of Sodium Bicarbonate Cotransporter Activity. Antioxidants 2022, 11, 1641. https://doi.org/10.3390/antiox11091641
Ji MJ, Son KH, Hong JH. Addition of oh8dG to Cardioplegia Attenuated Myocardial Oxidative Injury through the Inhibition of Sodium Bicarbonate Cotransporter Activity. Antioxidants. 2022; 11(9):1641. https://doi.org/10.3390/antiox11091641
Chicago/Turabian StyleJi, Min Jeong, Kuk Hui Son, and Jeong Hee Hong. 2022. "Addition of oh8dG to Cardioplegia Attenuated Myocardial Oxidative Injury through the Inhibition of Sodium Bicarbonate Cotransporter Activity" Antioxidants 11, no. 9: 1641. https://doi.org/10.3390/antiox11091641
APA StyleJi, M. J., Son, K. H., & Hong, J. H. (2022). Addition of oh8dG to Cardioplegia Attenuated Myocardial Oxidative Injury through the Inhibition of Sodium Bicarbonate Cotransporter Activity. Antioxidants, 11(9), 1641. https://doi.org/10.3390/antiox11091641