
Functional Characterization of Sugar Beet M14 Antioxidant Enzymes in Plant Salt Stress Tolerance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, and Growth Conditions
2.2. Isolation and Sequence Analysis of Genes
2.3. Quantitative Real-Time PCR Analysis
2.4. Generation and Phenotypic Analyses of Transgenic A. thaliana
2.5. Quantification of Biomass, MDA Content, H2O2 Content, Na+ and K+ Content, Morphological Index, and Physiological Indicators
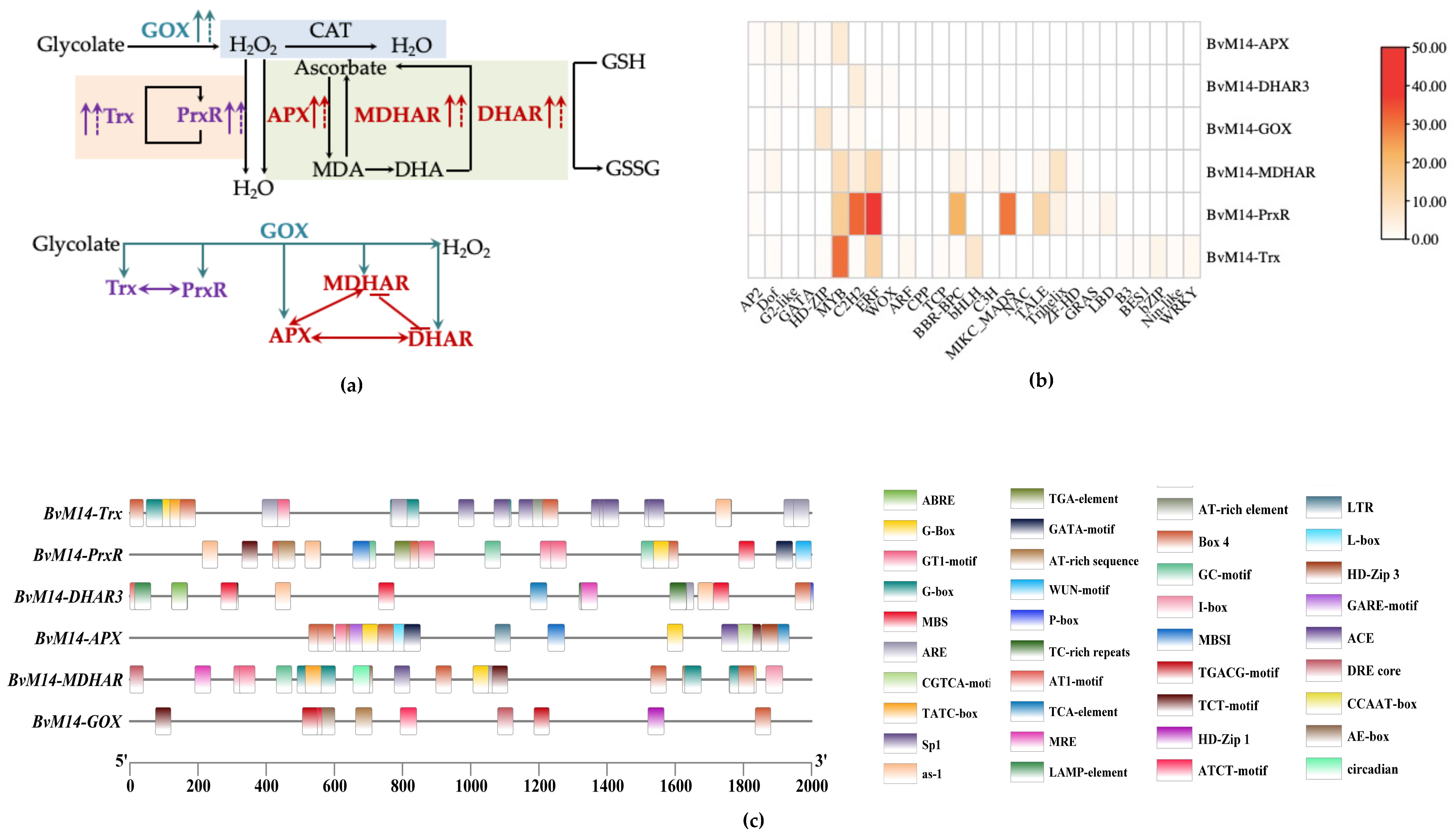
2.6. Cis-Regulatory Elements(CREs) Analysis
2.7. Statistical Analysis
3. Results
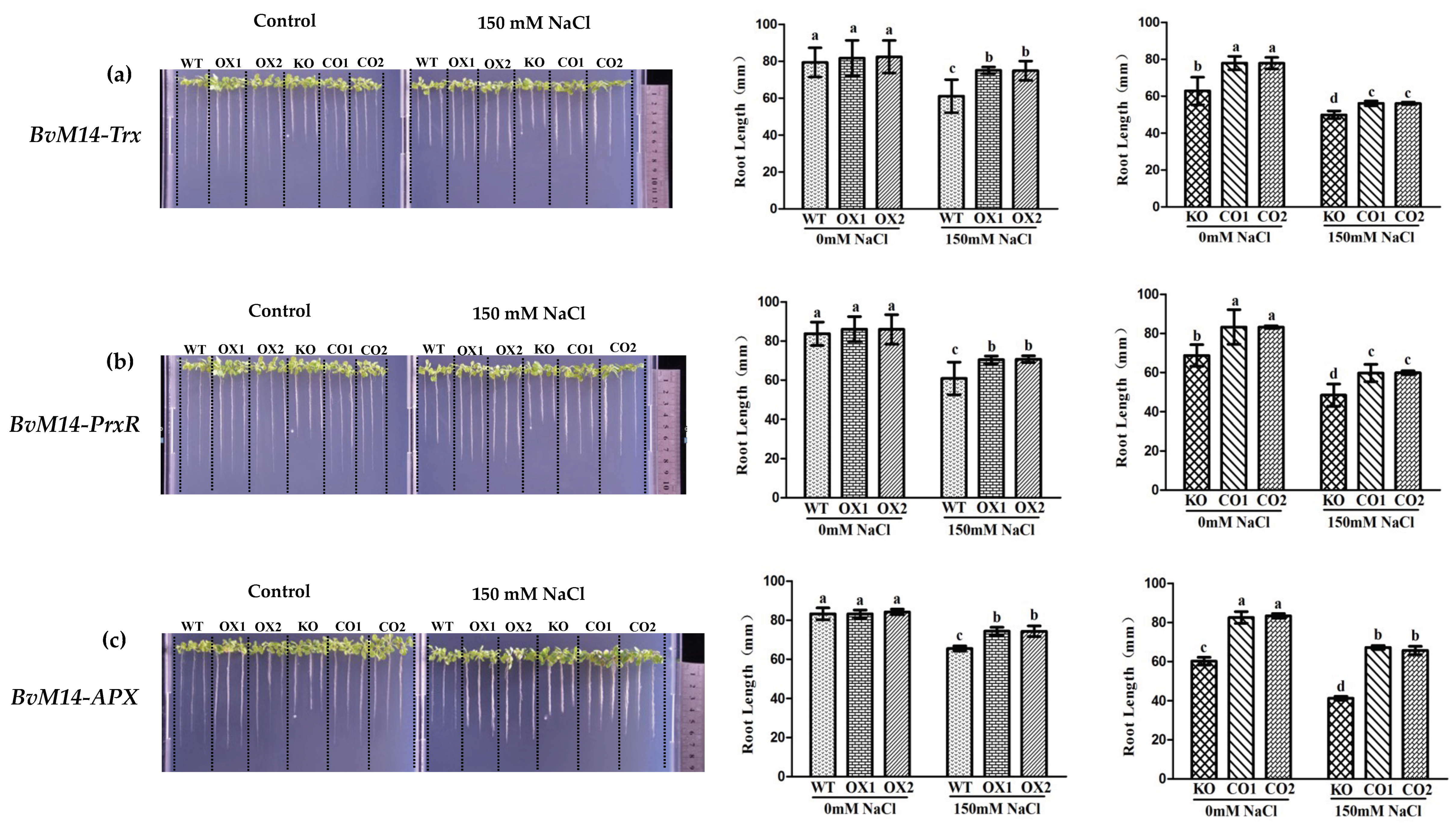
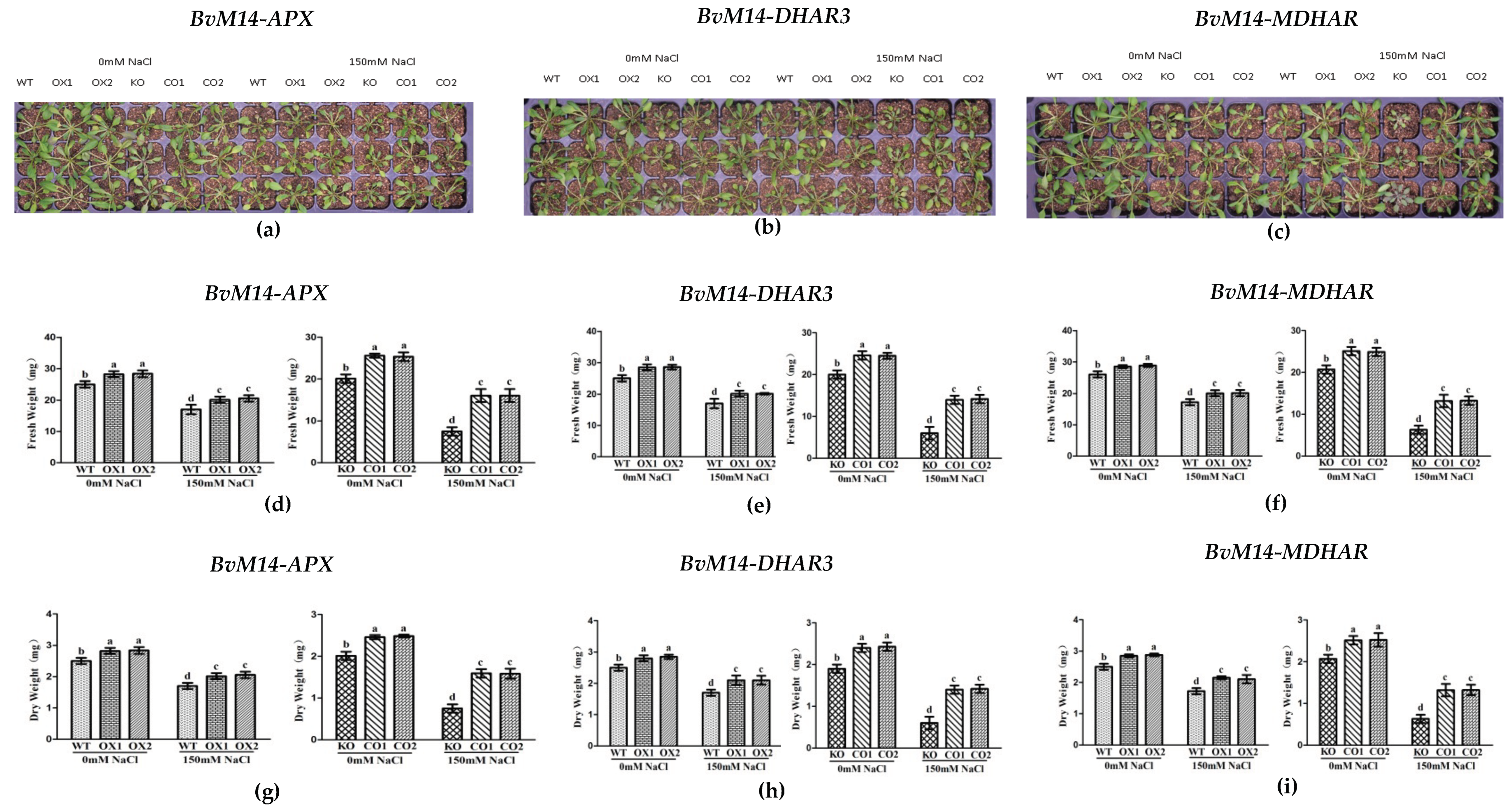
3.1. Ectopic Expression of BvM14 Antioxidant Enzymes Enhanced Plant Salt Tolerance
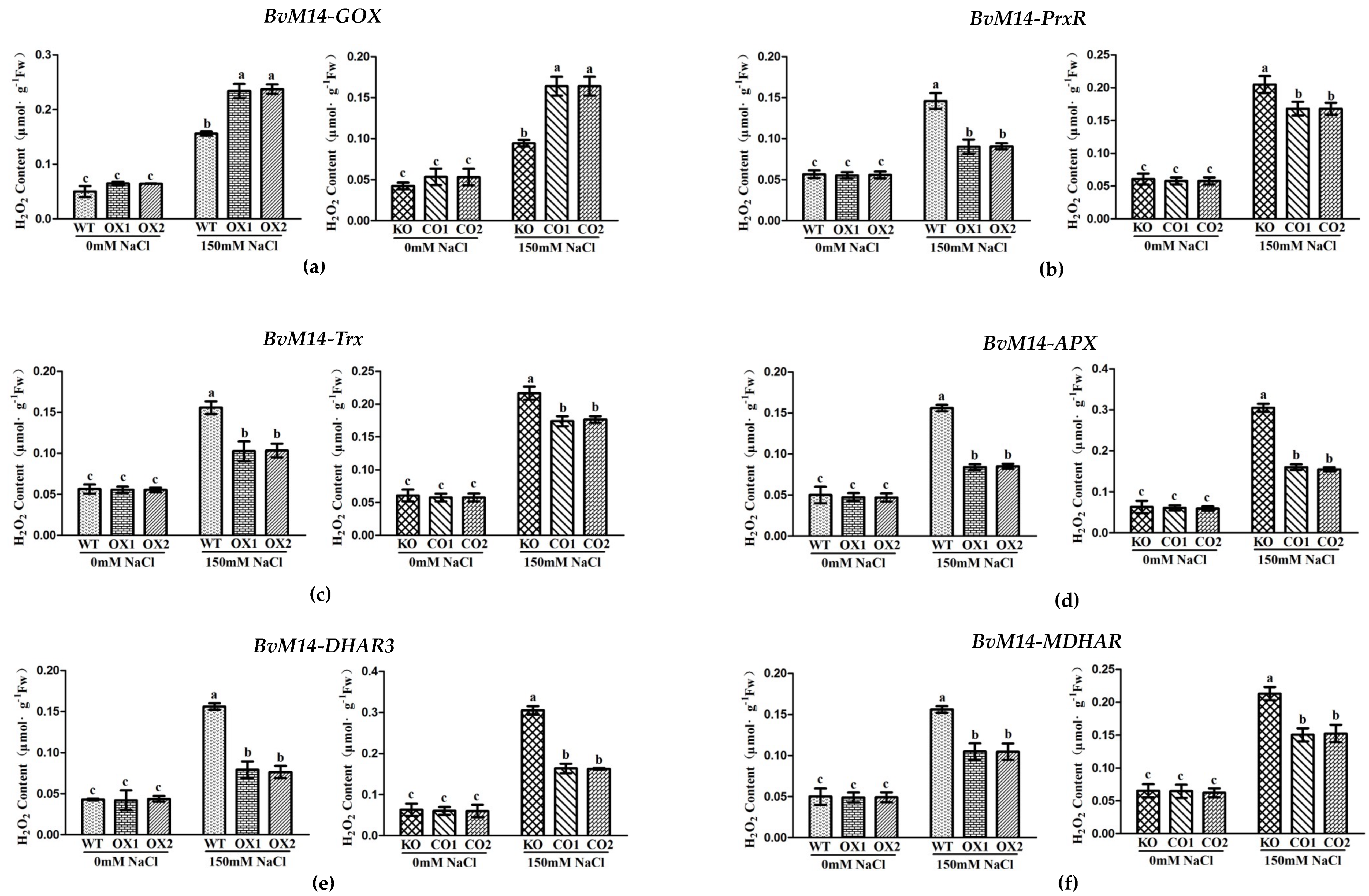
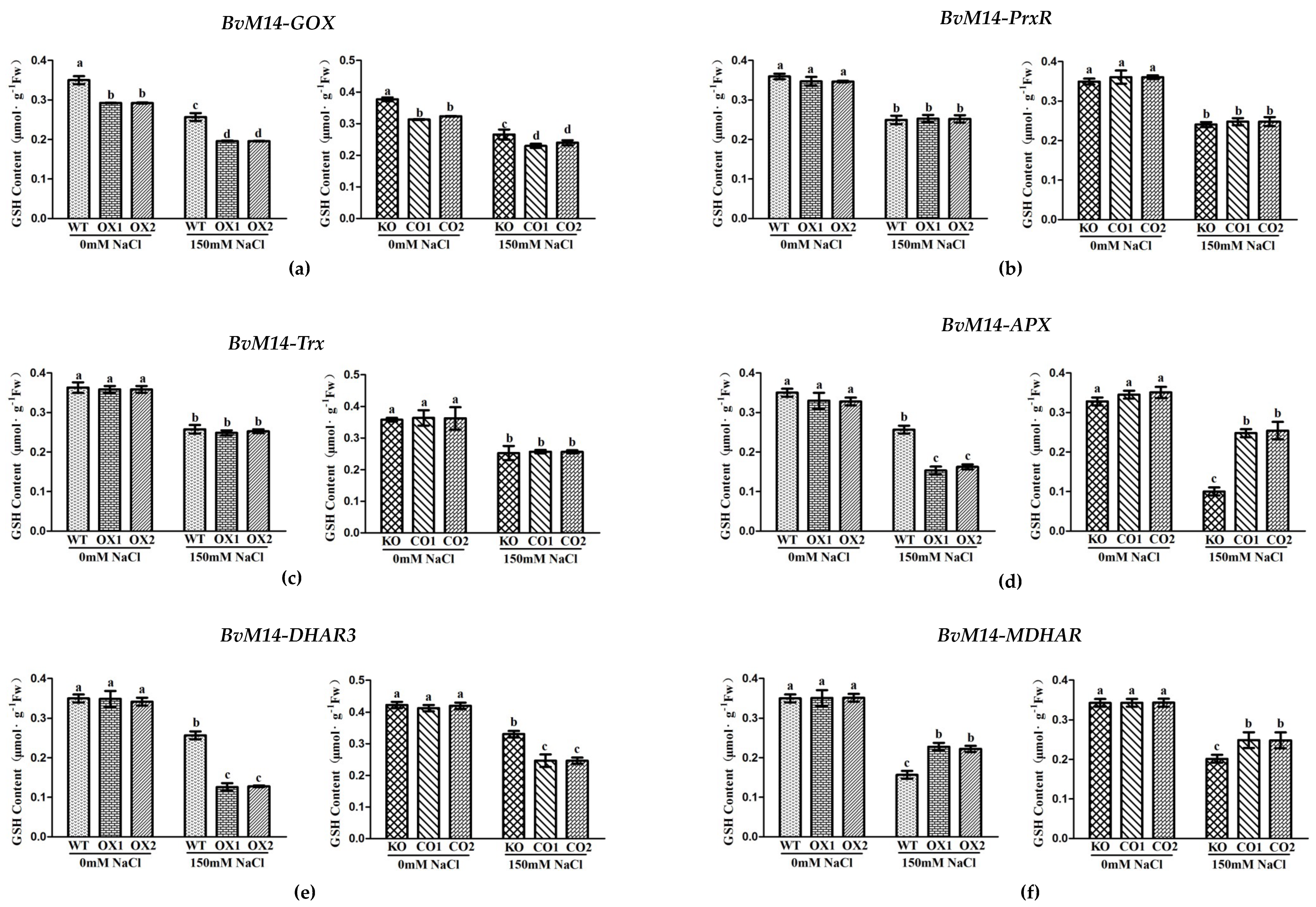
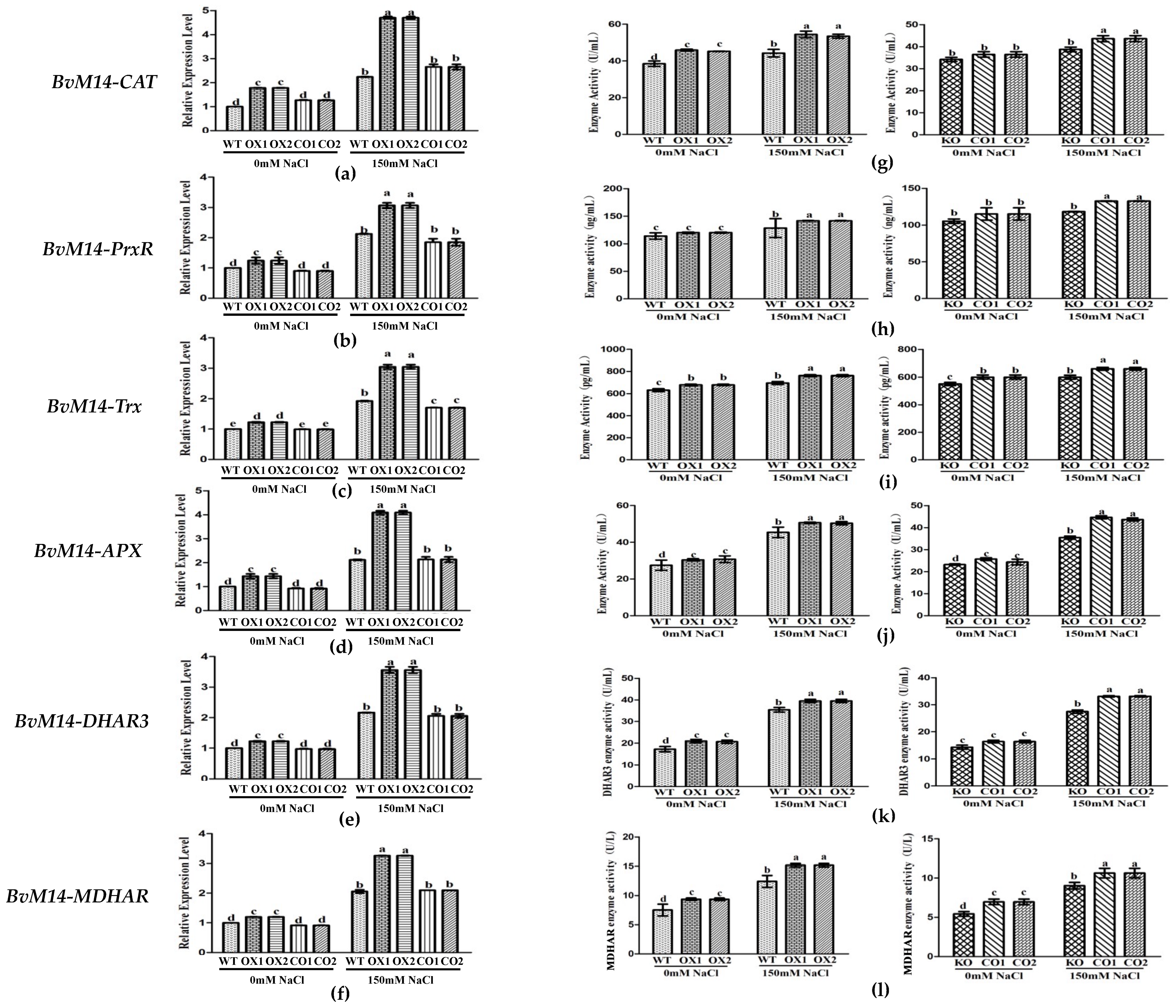
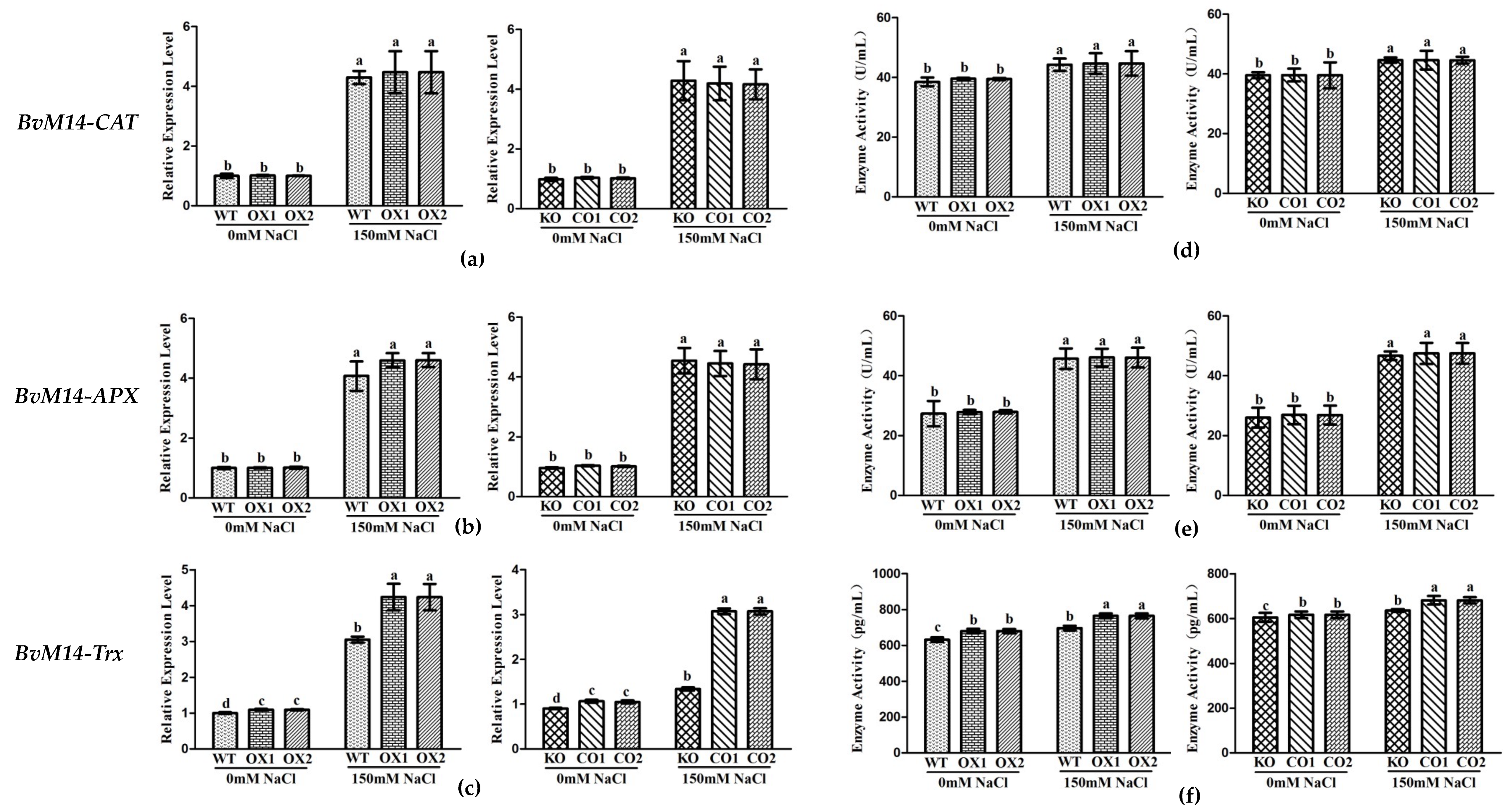
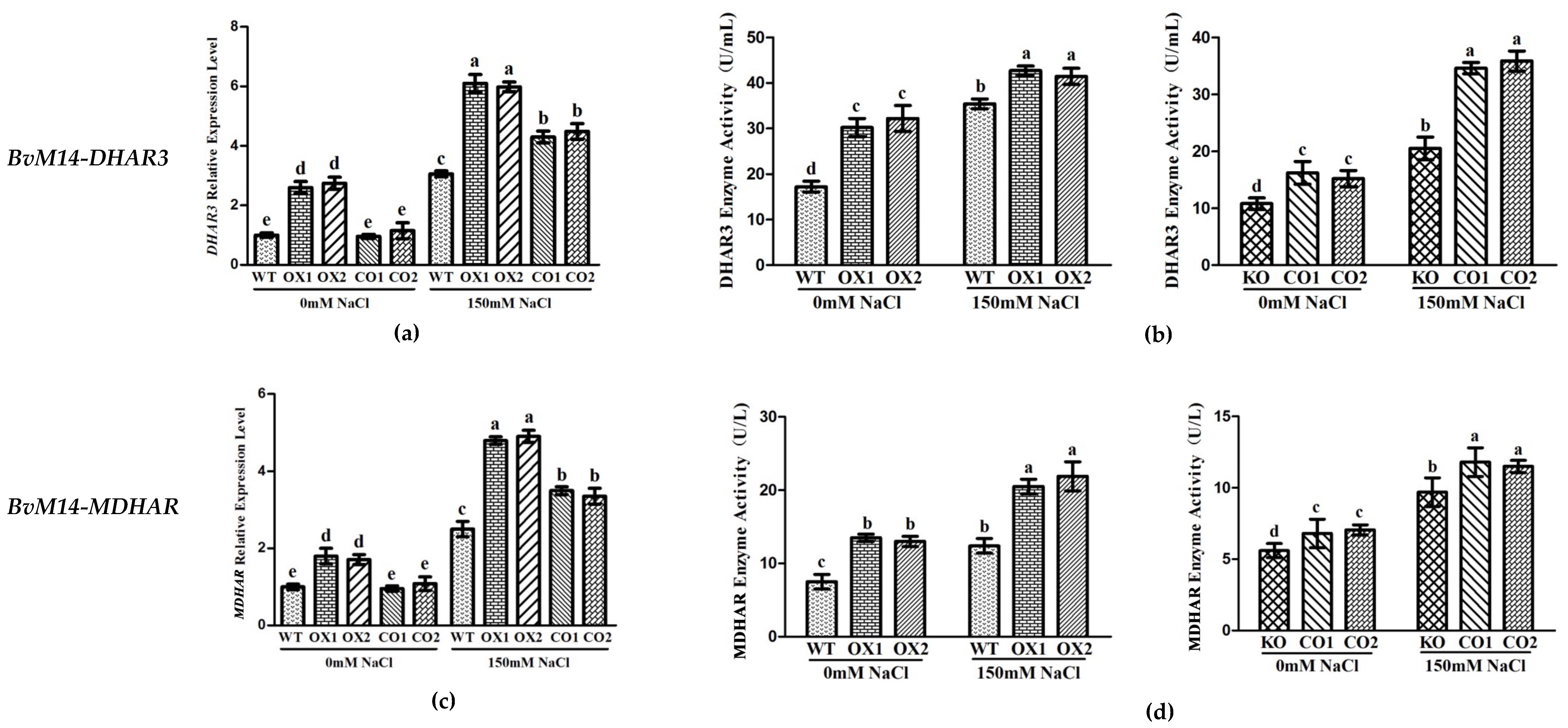
3.2. Ectopic Expression of Genes Encoding the Antioxidant Enzymes of BvM14 Improved Plant Antioxidant Capacity
3.3. Antioxidant Enzyme Activities of WT, Different KO, and Transgenic Plants under Salt Stress
3.4. Different Levels of Regulations of the Six Antioxidant Enzymes of BvM14
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Wang, K.; Ji, M.; Zhang, T.; Yang, C.; Liu, H.; Chen, S.; Li, H.; Li, H. Cys-SH based quantitative redox proteomics of salt induced response in sugar beet monosomic addition line M14. Bot Stud. 2021, 62, 16. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, X.; Zhang, J.; Li, J.; Chen, S.; Duanmu, H.; Li, H. Quantitative redox proteomics revealed molecular mechanisms of salt tolerance in the roots of sugar beet monomeric addition line M14. Bot Stud. 2022, 63, 5. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ma, C.; Wang, L.; Chen, S.; Li, H. Salt stress induced proteome and transcriptome changes in sugar beet monosomic addition line M14. J. Plant. Physiol. 2012, 169, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.; Zhu, N.; Koh, J.; Ma, C.; Pan, Y.; Yu, B.; Chen, S.; Li, H. Proteomic analysis of salt tolerance in sugar beet monosomic addition line M14. J. Proteome Res. 2013, 12, 4931–4950. [Google Scholar] [CrossRef]
- Mulet, J.M. Shaping the Sugar Beet of Tomorrow: Current Advances in Sugar Beet Biotechnology and New Breeding Techniques. In Sugar Beet Cultivation, Management and Processing; Springer: Berlin/Heidelberg, Germany, 2022; pp. 49–74. [Google Scholar]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-Dependent ROS Synthesis and ROS Scavenging by Plant Specialized Metabolites To Modulate Plant Development and Stress Responses. Chem. Res Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Steffens, B. The role of ethylene and ROS in salinity, heavy metal, and flooding responses in rice. Front Plant Sci. 2014, 5, 685. [Google Scholar] [CrossRef]
- Tyagi, S.; Shah, A.; Karthik, K.; Rathinam, M.; Rai, V.; Chaudhary, N.; Sreevathsa, R. Reactive oxygen species in plants: An invincible fulcrum for biotic stress mitigation. Appl. Microbiol. Biotechnol. 2022, 106, 5945–5955. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gong, S.; Fu, L.; Hu, G.; Li, G.; Hu, S.; Yang, J. The Involvement of Antioxidant Enzyme System, Nitrogen Metabolism and Osmoregulatory Substances in Alleviating Salt Stress in Inbred Maize Lines and Hormone Regulation Mechanisms. Plants 2022, 11, 1547. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, Y.; Fukushi, Y.; Wakabayashi, K.I.; Yoshida, K.; Hisabori, T. Oxidative regulation of chloroplast enzymes by thioredoxin and thioredoxin-like proteins in Arabidopsis thaliana. Proc. Natl. Acad. Sci USA 2021, 118, e2114952118. [Google Scholar] [CrossRef] [PubMed]
- Sainz, M.M.; Filippi, C.V.; Eastman, G.; Sotelo-Silveira, J.; Borsani, O.; Sotelo-Silveira, M. Analysis of Thioredoxins and Glutaredoxins in Soybean: Evidence of Translational Regulation under Water Restriction. Antioxidants 2022, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; ElSayed, A.I.; Moore, M.; Dietz, K.J. Redox and Reactive Oxygen Species Network in Acclimation for Salinity Tolerance in Sugar Beet. J. Exp. Bot. 2017, 68, 1283–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Pan, Y.; Zhang, Y.; Wu, C.; Ma, C.; Yu, B.; Zhu, N.; Koh, J.; Chen, S. Salt stress response of membrane proteome of sugar beet monosomic addition line M14. J. Proteom. 2015, 127 (Pt A), 18–33. [Google Scholar] [CrossRef]
- Lv, X.; Jin, Y.; Wang, Y. De novo transcriptome assembly and identification of salt-responsive genes in sugar beet M14. Comput. Biol. Chem. 2018, 75, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Nan, J.; Yu, B. OMICS Technologies and Applications in Sugar Beet. Front Plant Sci. 2016, 7, 900. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, S.; Tian, Y.; Wang, Q.; Chen, S.; Li, H.; Ma, C.; Li, H. Functional Characterization of a Sugar Beet BvbHLH93 Transcription Factor in Salt Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 3669. [Google Scholar] [CrossRef]
- Yu, B.; Li, J.; Koh, J.; Dufresne, C.; Yang, N.; Qi, S.; Zhang, Y.; Ma, C.; Duong, B.V.; Chen, S.; et al. Quantitative proteomics and phosphoproteomics of sugar beet monosomic addition line M14 in response to salt stress. J. Proteomics. 2016, 143, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, J.; Dong, J.; Song, X.; Jiang, J.; Li, H. Overexpression of Tamarix hispida ThTrx5 Confers Salt Tolerance to Arabidopsis by Activating Stress Response Signals. Int. J. Mol. Sci. 2020, 21, 1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Liu, D.; Liu, S. Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis. Plant Cell Rep. 2007, 26, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Noshi, M.; Hatanaka, R.; Tanabe, N.; Terai, Y.; Maruta, T.; Shigeoka, S. Redox regulation of ascorbate and glutathione by a chloroplastic dehydroascorbate reductase is required for high-light stress tolerance in Arabidopsis. Biosci. Biotechnol. Biochem. 2016, 80, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Qin, A.; Shi, Q.; Yu, X. Ascorbic acid contents in transgenic potato plants overexpressing two dehydroascorbate reductase genes. Mol. Biol. Rep. 2011, 38, 1557–1566. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants. 2021, 7, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.K. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]
- Leterrier, M.; Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; del Río, L.A. Peroxisomal monodehydroascorbate reductase. Genomic clone characterization and functional analysis under environmental stress conditions. Plant Physiol. 2005, 138, 2111–2123. [Google Scholar]
- Rahantaniaina, M.S.; Li, S.; Chatel-Innocenti, G.; Tuzet, A.; Issakidis-Bourguet, E.; Mhamdi, A.; Noctor, G. Cytosolic and Chloroplastic DHARs Cooperate in Oxidative Stress-Driven Activation of the Salicylic Acid Pathway. Plant Physiol. 2017, 174, 956–971. [Google Scholar] [CrossRef] [Green Version]
- Sultana, S.; Khew, C.Y.; Morshed, M.M.; Namasivayam, P.; Napis, S.; Ho, C.L. Overexpression of monodehydroascorbate reductase from a mangrove plant (AeMDHAR) confers salt tolerance on rice. J. Plant Physiol. 2012, 169, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Vanacker, H.; Guichard, M.; Bohrer, A.S.; Issakidis-Bourguet, E. Redox Regulation of Monodehydroascorbate Reductase by Thioredoxin y in Plastids Revealed in the Context of Water Stress. Antioxidants 2018, 7, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Z.; Peng, Y.; Wang, X.; Peng, D.; Li, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; et al. Clones of FeSOD, MDHAR, DHAR Genes from White Clover and Gene Expression Analysis of ROS-Scavenging Enzymes during Abiotic Stress and Hormone Treatments. Molecules 2015, 20, 20939–20954. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liao, M.; Huang, J.; Xu, Z.; Lin, Z.; Ye, N.; Zhang, Z.; Peng, X. Glycolate oxidase-dependent H(2)O(2) production regulates IAA biosynthesis in rice. BMC Plant Biol. 2021, 21, 326. [Google Scholar]
- Liu, Y.; Wu, W.; Chen, Z. Structures of glycolate oxidase from Nicotiana benthamiana reveal a conserved pH sensor affecting the binding of FMN. Biochem. Biophys. Res. Commun. 2018, 503, 3050–3056. [Google Scholar] [CrossRef]
- Schmitz, J.; Hüdig, M.; Meier, D.; Linka, N.; Maurino, V.G. The genome of Ricinus communis encodes a single glycolate oxidase with different functions in photosynthetic and heterotrophic organs. Planta 2020, 252, 100. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Xie, Z.; Li, X.; He, Z.H.; Peng, X.X. Association-Dissociation of Glycolate Oxidase with Catalase in Rice: A Potential Switch to Modulate Intracellular H2O2 Levels. Mol. Plant. 2016, 9, 737–748. [Google Scholar] [CrossRef]
- Formentin, E.; Sudiro, C.; Ronci, M.B.; Locato, V.; Barizza, E.; Stevanato, P.; Ijaz, B.; Zottini, M.; De Gara, L.; Lo Schiavo, F. Signature and Innate Antioxidative Profile Make the Difference Between Sensitivity and Tolerance to Salt in Rice Cells. Front. Plant Sci. 2018, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Liebthal, M.; Maynard, D.; Dietz, K.J. Peroxiredoxins and Redox Signaling in Plants. Antioxid. Redox. Signal. 2018, 28, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox. Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef]
- Zhai, J.; Qi, Q.; Wang, M.; Yan, J.; Li, K.; Xu, H. Overexpression of tomato thioredoxin h (SlTrxh) enhances excess nitrate stress tolerance in transgenic tobacco interacting with SlPrx protein. Plant Sci. 2022, 315, 111137. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.G.; Park, H.J.; Cha, J.Y.; Kim, J.A.; Shin, G.I.; Jeong, S.Y.; Lee, E.S.; Yun, D.J.; Lee, S.Y.; Kim, W.Y. Expression of Arabidopsis thaliana Thioredoxin-h2 in Brassica napus enhances antioxidant defenses and improves salt tolerance. Plant Physiol. Biochem. 2020, 147, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int J Mol Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [Green Version]
- Aleem, M.; Aleem, S.; Sharif, I.; Aleem, M.; Shahzad, R.; Khan, M.I.; Batool, A.; Sarwar, G.; Farooq, J.; Iqbal, A.; et al. Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response. Antioxidants 2022, 11, 1626. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Wu, J.; Zheng, X.; Zheng, S.; Sun, X.; Qiu, Q.; Lu, T. Gene knockout study reveals that cytosolic ascorbate peroxidase 2(OsAPX2) plays a critical role in growth and reproduction in rice under drought, salt and cold stresses. PLoS ONE 2013, 8, e57472. [Google Scholar] [CrossRef]
- Wang, Y.C.; Qu, G.Z.; Li, H.Y.; Wu, Y.J.; Wang, C.; Liu, G.F.; Yang, C.P. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 2010, 37, 1119–1124. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, J.; Chang, Q.; Gu, C.; Song, A.; Chen, S.; Dong, B.; Chen, F. Cold acclimation induces freezing tolerance via antioxidative enzymes, proline metabolism and gene expression changes in two chrysanthemum species. Mol. Biol. Rep. 2014, 41, 815–822. [Google Scholar] [CrossRef]
- Wang, D.R.; Yang, K.; Wang, X.; You, C.X. A C2H2-type zinc finger transcription factor, MdZAT17, acts as a positive regulator in response to salt stress. J. Plant Physiol. 2022, 275, 153737. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, L.; Xin, M.; Xing, H.; Zhang, Y.; Wu, J.; Xu, D.; Wang, Y.; Shang, Y.; Wei, L.; et al. Overexpression of GhABF3 increases cotton(Gossypium hirsutum L.) tolerance to salt and drought. BMC Plant Biol. 2022, 22, 313. [Google Scholar]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, I.; Iqbal, M.; Hakvoort, H.W.J.; de Boer, A.H.; Schat, H. Analysis of Arabidopsis thaliana HKT1 and Eutrema salsugineum/botschantzevii HKT1;2 Promoters in Response to Salt Stress in Athkt1:1 Mutant. Mol. Biotechnol. 2019, 61, 442–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wen, L.; Liao, L.; Lin, S.; Zheng, E.; Li, Y.; Zhang, Y. Comparative transcriptome analysis unveiling reactive oxygen species scavenging system of Sonneratia caseolaris under salinity stress. Front Plant Sci. 2022, 13, 953450. [Google Scholar] [CrossRef]
- Chen, Y.; Dai, Y.; Li, Y.; Yang, J.; Jiang, Y.; Liu, G.; Yu, C.; Zhong, F.; Lian, B.; Zhang, J. Overexpression of the Salix matsudana SmAP2-17 gene improves Arabidopsis salinity tolerance by enhancing the expression of SOS3 and ABI5. BMC Plant Biol. 2022, 22, 102. [Google Scholar]
- Yu, Z.; Yan, H.; Liang, L.; Zhang, Y.; Yang, H.; Li, W.; Choi, J.; Huang, J.; Deng, S. A C2H2-Type Zinc-Finger Protein from Millettia pinnata, MpZFP1, Enhances Salt Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 10832. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Yu, B.; Ma, C.; Li, H.; Jiang, D.; Nan, J.; Xu, M.; Liu, H.; Chen, S.; Duanmu, H.; et al. Functional Characterization of Sugar Beet M14 Antioxidant Enzymes in Plant Salt Stress Tolerance. Antioxidants 2023, 12, 57. https://doi.org/10.3390/antiox12010057
Li J, Yu B, Ma C, Li H, Jiang D, Nan J, Xu M, Liu H, Chen S, Duanmu H, et al. Functional Characterization of Sugar Beet M14 Antioxidant Enzymes in Plant Salt Stress Tolerance. Antioxidants. 2023; 12(1):57. https://doi.org/10.3390/antiox12010057
Chicago/Turabian StyleLi, Jinna, Bing Yu, Chunquan Ma, Hongli Li, Desheng Jiang, Jingdong Nan, Meng Xu, He Liu, Sixue Chen, Huizi Duanmu, and et al. 2023. "Functional Characterization of Sugar Beet M14 Antioxidant Enzymes in Plant Salt Stress Tolerance" Antioxidants 12, no. 1: 57. https://doi.org/10.3390/antiox12010057
APA StyleLi, J., Yu, B., Ma, C., Li, H., Jiang, D., Nan, J., Xu, M., Liu, H., Chen, S., Duanmu, H., & Li, H. (2023). Functional Characterization of Sugar Beet M14 Antioxidant Enzymes in Plant Salt Stress Tolerance. Antioxidants, 12(1), 57. https://doi.org/10.3390/antiox12010057