

The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications

 , , and
, , and 
Abstract
:1. Introduction
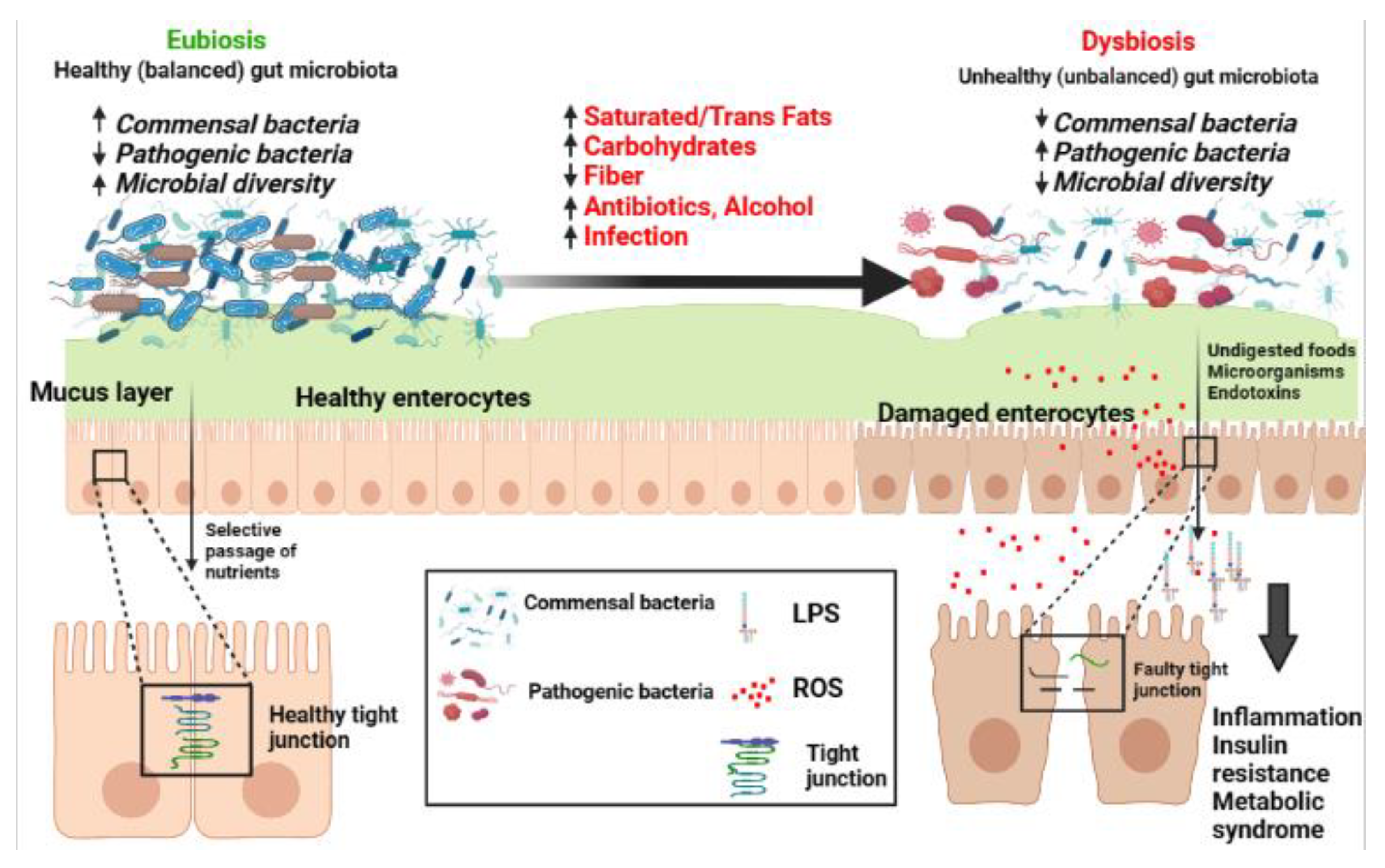
2. Gut Physiology and Microbiota
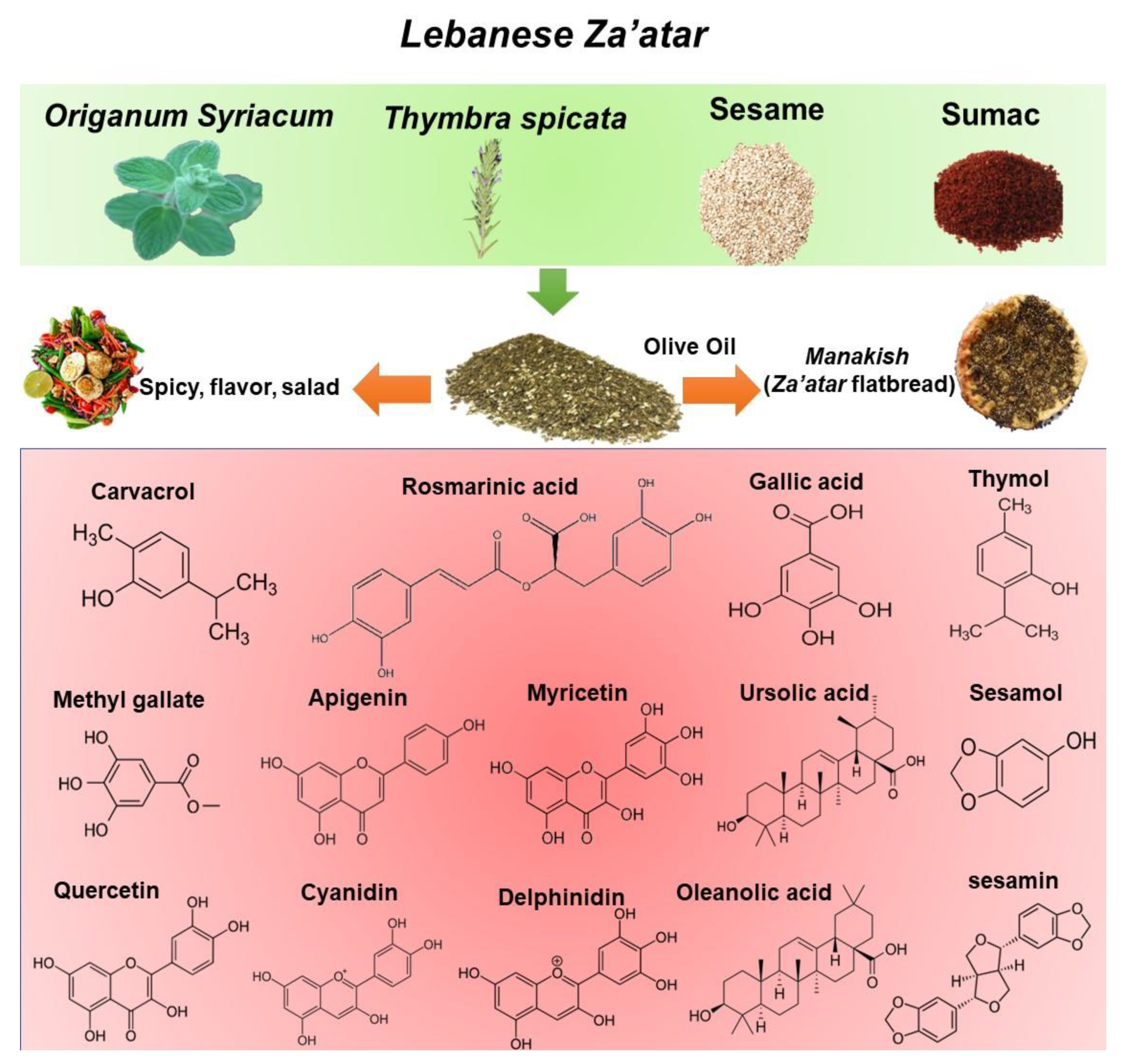
3. Za’atar Mixture
3.1. Chemical and Phytochemical Composition of Mixed Za’atar

{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Local Name (Lebanon) | Used Part | Phenolic Contents mgGAE/g Dry Extract | Fiber g/100 g | Oils/Fatty Acids g/100 g | Essential Oils | Minerals (in Order of Abundance) | Vitamins | References |
|---|---|---|---|---|---|---|---|---|---|
| Origanum Syriacum | Za’atar (Zouba’a) | Leaves | 120–250 | 15 | 0.13 | Carvacrol, thymol | ND | C | [38,48,49,50] |
| Thymbra spicata | Wild Za’atar (Za’atar dakki) | Leaves | 90–250 | ND | PA OA | Carvacrol | ND | ND | [37,38,39] |
| Rhus coriaria | Sumac | Fruits | 48–140 | 14 | 7–18 (37.7% OA, 34.8% LA, 27.4% PA, 17.3% SA) | β-caryophyllene, α-pinene | K, Ca, Mg, P | B1, B2, B6, B12, C | [51,52,53,54] |
| Sesamum indicum | Sesame | Seeds | 7–80 | 6–10.8 | 51.9 OA (43%) LA (35%), PA (11%) SA (7%) | ND | Ca, K, P, Mg, Fe | Carotene, B1, B2, B3, E | [55,56,57] |
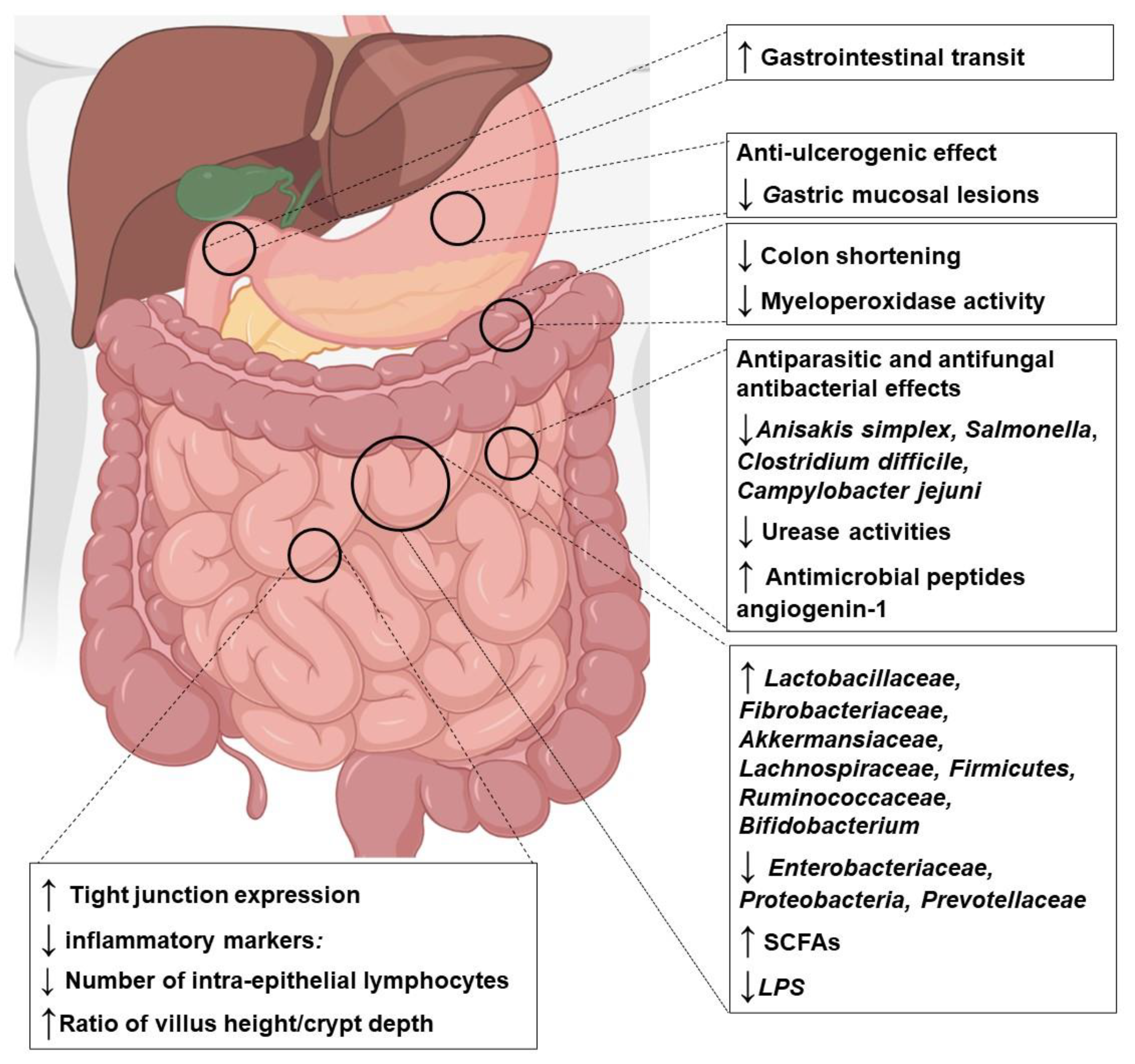
3.2. Effects of Za’atar Compounds on Microbiota and Intestinal Disorders: Cellular and Animal Studies
3.2.1. Origanum syriacum and Thymbra spicata Extracts
3.2.2. Carvacrol, Thymol, and Essential Oils (EOs)
3.2.3. Rosmarinic Acid
3.2.4. Oleanolic and Ursolic Acid
3.2.5. Rhus coriaria (Sumac)
3.2.6. Sesamum indicum (Sesame)
3.3. Clinical Studies (Table 3)
| Authors (Year) | Sample Size | Gender M/F (Age) | Participants | Format, Dose | Duration of Study | Main Findings |
|---|---|---|---|---|---|---|
| [153] | 51 | 27/39 (≥60 years) | Subjects with increased IP | PR-diet 724 mg/day | 8 weeks | ↓ Serum zonulin ↓ Blood pressure ↑ Fiber-fermenting bacteria, butyrate-producing bacteria |
| [156] | 36 | 11/25 (8–65 years) | Subjects with chronic constipation | Mango consumption, 300 g/day | 4 weeks | ↑ Stool frequency, consistency ↑ Gastrin levels, SCFA (valeric acid) ↓ Endotoxin and IL-6 |
| [157] | 32 | 22/10 (18–50 years) | healthy lean and obese individuals | Mango consumption, 400 g/day | 6 weeks | ↑ SCFA in lean individuals |
| [139] | 10 | 5/5 (18–75 years) | IBD Subjects | Mango pulp intake, 200–400 g/day | 8 weeks | ↑ SCCAI ↓ IL-8 ↓ GRO and GM-CSF ↑ Fecal butyric acid ↓ Inflammation biomarkers ↑ Lactobacillus plantarum, Lactobacillus reuteri, Lactobacillus lactis |
| [159] | 64 | 33/31 (19–79) | Subjects with symptoms and signs of SBO | Sesame oil in nasogastric tube, 150 mL/day | 1 day | ↓ SBO resolution time ↓ Hospital stays ↓ Relaparotomy rate |
4. Summary
5. Clinical Perspectives and Future Applications
- Well-designed human studies evaluating drug interaction with herbal supplements are required.
- Controlled clinical trials evaluating the acceptance, tolerance, and safety of herbal preparation are highly needed.
- Standardized preparations of herbal products or traditional foods should be adjusted to an exact content of substances with well-known nutraceutical properties.
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portincasa, P.; Bonfrate, L.; Khalil, M.; Angelis, M.D.; Calabrese, F.M.; D’Amato, M.; Wang, D.Q.-H.; Di Ciaula, A. Intestinal Barrier and Permeability in Health, Obesity and NAFLD. Biomedicines 2022, 10, 83. [Google Scholar] [CrossRef]
- Scaldaferri, F.; Pizzoferrato, M.; Gerardi, V.; Lopetuso, L.; Gasbarrini, A. The gut barrier: New acquisitions and therapeutic approaches. J. Clin. Gastroenterol. 2012, 46, S12–S17. [Google Scholar] [CrossRef]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Stress and the gut: Pathophysiology, clinical consequences, diagnostic approach and treatment options. J. Physiol. Pharmacol. 2011, 62, 591–599. [Google Scholar]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.-H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Kallus, S.J.; Brandt, L.J. The intestinal microbiota and obesity. J. Clin. Gastroenterol. 2012, 46, 16–24. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Biesalski, H.K. Nutrition meets the microbiome: Micronutrients and the microbiota. Ann. N. Y. Acad. Sci. 2016, 1372, 53–64. [Google Scholar] [CrossRef]
- Gao, K.; Mu, C.-L.; Farzi, A.; Zhu, W.-Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2019, 11, 709–723. [Google Scholar] [CrossRef]
- De Angelis, M.; Garruti, G.; Minervini, F.; Bonfrate, L.; Portincasa, P.; Gobbetti, M. The Food-gut Human Axis: The Effects of Diet on Gut Microbiota and Metabolome. Curr. Med. Chem. 2019, 26, 3567–3583. [Google Scholar] [CrossRef]
- France, M.M.; Turner, J.R. The mucosal barrier at a glance. J. Cell Sci. 2017, 130, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Salvo Romero, E.; Alonso Cotoner, C.; Pardo Camacho, C.; Casado Bedmar, M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef]
- Camilleri, M.; Madsen, K.; Spiller, R.; Van Meerveld, B.G.; Verne, G.N. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2012, 24, 503–512. [Google Scholar] [CrossRef]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of Barrier Function in Injured Intestinal Mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Belancic, A. Gut microbiome dysbiosis and endotoxemia—Additional pathophysiological explanation for increased COVID-19 severity in obesity. Obes. Med. 2020, 20, 100302. [Google Scholar] [CrossRef]
- Salguero, M.V.; Al-Obaide, M.A.I.; Singh, R.; Siepmann, T.; Vasylyeva, T.L. Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp. Ther. Med. 2019, 18, 3461–3469. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Kimera, F.; Sewilam, H.; Fouad, W.M.; Suloma, A. Sustainable production of Origanum syriacum L. using fish effluents improved plant growth, yield, and essential oil composition. Heliyon 2021, 7, e06423. [Google Scholar] [CrossRef]
- Alwafa, R.A.; Mudalal, S.; Shraim, F.; Mauriello, G. Comparison between Quality Traits of Solar-Dried and Freeze-Dried Origanum syriacum L. (Za’atar). Plants 2022, 11, 1110. [Google Scholar] [CrossRef]
- Mudalal, S.; Kanan, D.; Anabtawi, O.; Irshaid, A.; Sabbah, M.; Shtaya, M.; Shraim, F.; Mauriello, G. Application of the Hurdle Technology Concept to the Fresh Za’atar (Origanum syriacum) Preservation. Foods 2022, 11, 3002. [Google Scholar] [CrossRef]
- Alwafa, R.A.; Mudalal, S.; Mauriello, G. Origanum syriacum L. (Za’atar), from Raw to Go: A Review. Plants 2021, 10, 1001. [Google Scholar] [CrossRef]
- Khalil, M.; Caponio, G.R.; Diab, F.; Shanmugam, H.; Di Ciaula, A.; Khalifeh, H.; Vergani, L.; Calasso, M.; De Angelis, M.; Portincasa, P. Unraveling the beneficial effects of herbal Lebanese mixture “Za’atar”. History, studies, and properties of a potential healthy food ingredient. J. Funct. Foods 2022, 90, 104993. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Serale, N.; Parodi, A.; Voci, A.; Vergani, L.; Daher, A. Antitumor Activity of Ethanolic Extract from Thymbra spicata L. aerial Parts: Effects on Cell Viability and Proliferation, Apoptosis Induction, STAT3, and NF-kB Signaling. Nutr. Cancer 2020, 73, 1193–1206. [Google Scholar] [CrossRef]
- Khoury, M.; Stien, D.; Eparvier, V.; Ouaini, N.; El Beyrouthy, M. Report on the Medicinal Use of Eleven Lamiaceae Species in Lebanon and Rationalization of Their Antimicrobial Potential by Examination of the Chemical Composition and Antimicrobial Activity of Their Essential Oils. Evid. Based Complement. Altern. Med. 2016, 2016, 2547169. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Bartolucci, F.; Canale, A.; Maggi, F. Origanum syriacum subsp. syriacum: From an ingredient of Lebanese ‘manoushe’ to a source of effective and eco-friendly botanical insecticides. Ind. Crop. Prod. 2019, 134, 26–32. [Google Scholar] [CrossRef]
- Khalil, M.; Hayek, S.; Khalil, N.; Serale, N.; Vergani, L.; Calasso, M.; De Angelis, M.; Portincasa, P. Role of Sumac (Rhus coriaria L.) in the management of metabolic syndrome and related disorders: Focus on NAFLD-atherosclerosis interplay. J. Funct. Foods 2021, 87, 104811. [Google Scholar] [CrossRef]
- Toker, E.B.; Yeşilbağ, K. In vitro antiviral activity of Thymbra spicata L. extract on bovine respiratory viruses (BCoV, BPIV-3, BRSV, BVDV and BoHV-1). J. Appl. Microbiol. 2022, 132, 2625–2632. [Google Scholar] [CrossRef]
- Mesmar, J.; Abdallah, R.; Badran, A.; Maresca, M.; Baydoun, E. Origanum syriacum Phytochemistry and Pharmacological Properties: A Comprehensive Review. Molecules 2022, 27, 4272. [Google Scholar] [CrossRef]
- AlKahlout, A.; Fardoun, M.; Mesmar, J.; Abdallah, R.; Badran, A.; Nasser, S.A.; Baydoun, S.; Kobeissy, F.; Shaito, A.; Iratni, R.; et al. Origanum syriacum L. Attenuates the Malignant Phenotype of MDA-MB231 Breast Cancer Cells. Front. Oncol. 2022, 12, 922196. [Google Scholar] [CrossRef]
- Kerem, S.; Koşar, N.; Tekin, F.; Güreser, A.S.; Özbek, Ö. Investigation of antimicrobial activities and molecular characterization of the species belong to Origanum, Thymus and Thymbra genera by ISSR. Mol. Biol. Rep. 2023, 50, 289–298. [Google Scholar] [CrossRef]
- Diab, F.; Khalil, M.; Lupidi, G.; Zbeeb, H.; Salis, A.; Damonte, G.; Bramucci, M.; Portincasa, P.; Vergani, L. Influence of Simulated In Vitro Gastrointestinal Digestion on the Phenolic Profile, Antioxidant, and Biological Activity of Thymbra spicata L. Extracts. Antioxidants 2022, 11, 1778. [Google Scholar] [CrossRef]
- Alonazi, M.A.; Jemel, I.; Moubayed, N.; Alwhibi, M.; El-Sayed, N.N.E.; Ben Bacha, A. Evaluation of the in vitro anti-inflammatory and cytotoxic potential of ethanolic and aqueous extracts of Origanum syriacum and Salvia lanigera leaves. Environ. Sci. Pollut. Res. Int. 2021, 28, 19890–19900. [Google Scholar] [CrossRef]
- Sedeek, M.S.; Afifi, S.M.; Mansour, M.K.; Hassan, M.; Mehaya, F.M.; Naguib, I.A.; Abourehab, M.A.S.; Farag, M.A. Unveiling Antimicrobial and Antioxidant Compositional Differences between Dukkah and Za’atar via SPME-GCMS and HPLC-DAD. Molecules 2022, 27, 6471. [Google Scholar] [CrossRef]
- Shen, D.; Pan, M.H.; Wu, Q.L.; Park, C.H.; Juliani, H.R.; Ho, C.T.; Simon, J.E. LC-MS method for the simultaneous quantitation of the anti-inflammatory constituents in oregano (Origanum species). J. Agric. Food Chem. 2010, 58, 7119–7125. [Google Scholar] [CrossRef]
- Al Hafi, M.; El Beyrouthy, M.; Ouaini, N.; Stien, D.; Rutledge, D.; Chaillou, S. Chemical Composition and Antimicrobial Activity of Satureja, Thymus, and Thymbra Species Grown in Lebanon. Chem. Biodivers. 2017, 14, e1600236. [Google Scholar] [CrossRef]
- Gedikoğlu, A.; Sökmen, M.; Çivit, A. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Sci. Nutr. 2019, 7, 1704–1714. [Google Scholar] [CrossRef]
- Dorman, H.J.; Bachmayer, O.; Kosar, M.; Hiltunen, R. Antioxidant properties of aqueous extracts from selected lamiaceae species grown in Turkey. J. Agric. Food Chem. 2004, 52, 762–770. [Google Scholar] [CrossRef]
- Khalil, M.; Khalifeh, H.; Baldini, F.; Salis, A.; Damonte, G.; Daher, A.; Voci, A.; Vergani, L. Antisteatotic and antioxidant activities of Thymbra spicata L. extracts in hepatic and endothelial cells as in vitro models of non-alcoholic fatty liver disease. J. Ethnopharmacol. 2019, 239, 111919. [Google Scholar] [CrossRef]
- Prasad Mn, N.; Kr, S.; Prasad, D.S. A Review on Nutritional and Nutraceutical Properties of Sesame. J. Nutr. Food Sci. 2012, 2, 1–6. [Google Scholar] [CrossRef]
- Elleuch, M.; Besbes, S.; Roiseux, O.; Blecker, C.; Attia, H. Quality characteristics of sesame seeds and by-products. Food Chem. 2007, 103, 641–650. [Google Scholar] [CrossRef]
- Saydut, A.; Duz, M.; Kaya, C.; Kafadar, A.; Hamamci, C. Transesterified sesame (Sesamum indicum L.) seed oil as a biodiesel fuel. Bioresour. Technol. 2008, 99, 6656–6660. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Mansour, Z.R. Sesamol, a major lignan in sesame seeds (Sesamum indicum): Anti-cancer properties and mechanisms of action. Eur. J. Pharmacol. 2019, 855, 75–89. [Google Scholar] [CrossRef]
- Regazzoni, L.; Arlandini, E.; Garzon, D.; Santagati, N.A.; Beretta, G.; Facino, R.M. A rapid profiling of gallotannins and flavonoids of the aqueous extract of Rhus coriaria L. by flow injection analysis with high-resolution mass spectrometry assisted with database searching. J. Pharm. Biomed. Anal. 2013, 72, 202–207. [Google Scholar] [CrossRef]
- Gök, H.N.; Deliorman Orhan, D.; Gürbüz, İ.; Aslan, M. Activity-guided isolation of α-amylase, α-glucosidase, and pancreatic lipase inhibitory compounds from Rhus coriaria L. J. Food Sci. 2020, 85, 3220–3228. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. HPLC-DAD-ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Mehrdad, M.; Zebardast, M.; Abedi, G.; Koupaei, M.N.; Rasouli, H.; Talebi, M. Validated high-throughput HPLC method for the analysis of flavonol aglycones myricetin, quercetin, and kaempferol in Rhus coriaria L. using a monolithic column. J. AOAC Int. 2009, 92, 1035–1043. [Google Scholar]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Shamseddine, L.; Chidiac, J.J. Composition’s effect of Origanum syriacum essential oils in the antimicrobial activities for the treatment of denture stomatitis. Odontology 2021, 109, 327–335. [Google Scholar] [CrossRef]
- Al Hafi, M.; El Beyrouthy, M.; Ouaini, N.; Stien, D.; Rutledge, D.; Chaillou, S. Chemical Composition and Antimicrobial Activity of Origanum libanoticum, Origanum ehrenbergii, and Origanum syriacum Growing Wild in Lebanon. Chem. Biodivers. 2016, 13, 555–560. [Google Scholar] [CrossRef]
- Khalil, M.; Bazzi, A.; Zeineddine, D.; Jomaa, W.; Daher, A.; Awada, R. Repressive effect of Rhus coriaria L. fruit extracts on microglial cells-mediated inflammatory and oxidative stress responses. J. Ethnopharmacol. 2021, 269, 113748. [Google Scholar] [CrossRef]
- Sakhr, K.; El Khatib, S. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac—Rhus coriaria): A review. Heliyon 2020, 6, e03207. [Google Scholar] [CrossRef] [Green Version]
- Kizil, S.; Turk, M. Microelement contents and fatty acid compositions of Rhus coriaria L. and Pistacia terebinthus L. fruits spread commonly in the south eastern Anatolia region of Turkey. Nat. Prod. Res. 2010, 24, 92–98. [Google Scholar] [CrossRef]
- Reidel, R.V.B.; Cioni, P.L.; Majo, L.; Pistelli, L. Evolution of Volatile Emission in Rhus coriaria Organs During Different Stages of Growth and Evaluation of the Essential Oil Composition. Chem. Biodivers. 2017, 14, e1700270. [Google Scholar] [CrossRef]
- Namiki, M. Nutraceutical functions of sesame: A review. Crit. Rev. Food Sci. Nutr. 2007, 47, 651–673. [Google Scholar] [CrossRef]
- Kermani, S.G.; Saeidi, G.; Sabzalian, M.R.; Gianinetti, A. Drought stress influenced sesamin and sesamolin content and polyphenolic components in sesame (Sesamum indicum L.) populations with contrasting seed coat colors. Food Chem. 2019, 289, 360–368. [Google Scholar] [CrossRef]
- Esmaeilzadeh Kenari, R.; Mohsenzadeh, F.; Amiri, Z.R. Antioxidant activity and total phenolic compounds of Dezful sesame cake extracts obtained by classical and ultrasound-assisted extraction methods. Food Sci. Nutr. 2014, 2, 426–435. [Google Scholar] [CrossRef]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary polyphenol impact on gut health and microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 690–711. [Google Scholar] [CrossRef]
- Burdulis, D.; Sarkinas, A.; Jasutiené, I.; Stackevicené, E.; Nikolajevas, L.; Janulis, V. Comparative study of anthocyanin composition, antimicrobial and antioxidant activity in bilberry (Vaccinium myrtillus L.) and blueberry (Vaccinium corymbosum L.) fruits. Acta Pol. Pharm. 2009, 66, 399–408. [Google Scholar]
- Sourabh, A.; Kanwar, S.S.; Sud, R.G.; Ghabru, A.; Sharma, O.P. Influence of phenolic compounds of Kangra tea [Camellia sinensis (L) O Kuntze] on bacterial pathogens and indigenous bacterial probiotics of Western Himalayas. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2013, 44, 709–715. [Google Scholar] [CrossRef]
- Chan, E.W.; Soh, E.Y.; Tie, P.P.; Law, Y.P. Antioxidant and antibacterial properties of green, black, and herbal teas of Camellia sinensis. Pharmacogn. Res. 2011, 3, 266–272. [Google Scholar] [CrossRef]
- Shen, X.; Sun, X.; Xie, Q.; Liu, H.; Zhao, Y.; Pan, Y.; Hwang, C.-A.; Wu, V.C.H. Antimicrobial effect of blueberry (Vaccinium corymbosum L.) extracts against the growth of Listeria monocytogenes and Salmonella enteritidis. Food Control 2014, 35, 159–165. [Google Scholar] [CrossRef]
- Vitetta, L.; Vitetta, G.; Hall, S. Immunological Tolerance and Function: Associations Between Intestinal Bacteria, Probiotics, Prebiotics, and Phages. Front. Immunol. 2018, 9, 2240. [Google Scholar] [CrossRef]
- Larrosa, M.; Luceri, C.; Vivoli, E.; Pagliuca, C.; Lodovici, M.; Moneti, G.; Dolara, P. Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models. Mol. Nutr. Food Res. 2009, 53, 1044–1054. [Google Scholar] [CrossRef]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2010, 93, 62–72. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef]
- López, V.; Pavela, R.; Gómez-Rincón, C.; Les, F.; Bartolucci, F.; Galiffa, V.; Petrelli, R.; Cappellacci, L.; Maggi, F.; Canale, A.; et al. Efficacy of Origanum syriacum Essential Oil against the Mosquito Vector Culex quinquefasciatus and the Gastrointestinal Parasite Anisakis simplex, with Insights on Acetylcholinesterase Inhibition. Molecules 2019, 24, 2563. [Google Scholar] [CrossRef]
- El-Meligy, R.M.; Awaad, A.S.; Soliman, G.A.; Kenawy, S.A.; Alqasoumi, S.I. Prophylactic and curative anti-ulcerogenic activity and the possible mechanisms of action of some desert plants. Saudi Pharm. J. 2017, 25, 387–396. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Ma, X.; Geng, S.; Jiang, X.; Huang, Q.; Hu, C.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [Google Scholar] [CrossRef]
- Hall, H.N.; Wilkinson, D.J.; Le Bon, M. Oregano essential oil improves piglet health and performance through maternal feeding and is associated with changes in the gut microbiota. Anim. Microbiome 2021, 3, 2. [Google Scholar] [CrossRef]
- Abouelezz, K.; Abou-Hadied, M.; Yuan, J.; Elokil, A.A.; Wang, G.; Wang, S.; Wang, J.; Bian, G. Nutritional impacts of dietary oregano and Enviva essential oils on the performance, gut microbiota and blood biochemicals of growing ducks. Animal 2019, 13, 2216–2222. [Google Scholar] [CrossRef]
- Wang, G.Y.; Chen, S.Y.; Chen, Y.Y.; Hong, C.J.; Hsu, Y.H.; Yen, G.C. Protective effect of rosmarinic acid-rich trichodesma khasianum clarke leaves against ethanol-induced gastric mucosal injury in vitro and in vivo. Phytomedicine 2021, 80, 153382. [Google Scholar] [CrossRef]
- Ou, J.; Huang, J.; Zhao, D.; Du, B.; Wang, M. Protective effect of rosmarinic acid and carnosic acid against streptozotocin-induced oxidation, glycation, inflammation and microbiota imbalance in diabetic rats. Food Funct. 2018, 9, 851–860. [Google Scholar] [CrossRef]
- Mooyottu, S.; Flock, G.; Upadhyay, A.; Upadhyaya, I.; Maas, K.; Venkitanarayanan, K. Protective Effect of Carvacrol against Gut Dysbiosis and Clostridium difficile Associated Disease in a Mouse Model. Front. Microbiol. 2017, 8, 625. [Google Scholar] [CrossRef]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [Google Scholar] [CrossRef]
- Yin, D.; Du, E.; Yuan, J.; Gao, J.; Wang, Y.; Aggrey, S.E.; Guo, Y. Supplemental thymol and carvacrol increases ileum Lactobacillus population and reduces effect of necrotic enteritis caused by Clostridium perfringes in chickens. Sci. Rep. 2017, 7, 7334. [Google Scholar] [CrossRef]
- Ran, C.; Hu, J.; Liu, W.; Liu, Z.; He, S.; Dan, B.C.; Diem, N.N.; Ooi, E.L.; Zhou, Z. Thymol and Carvacrol Affect Hybrid Tilapia through the Combination of Direct Stimulation and an Intestinal Microbiota-Mediated Effect: Insights from a Germ-Free Zebrafish Model. J. Nutr. 2016, 146, 1132–1140. [Google Scholar] [CrossRef]
- Upadhyay, A.; Arsi, K.; Wagle, B.R.; Upadhyaya, I.; Shrestha, S.; Donoghue, A.M.; Donoghue, D.J. Trans-Cinnamaldehyde, Carvacrol, and Eugenol Reduce Campylobacter jejuni Colonization Factors and Expression of Virulence Genes in Vitro. Front. Microbiol. 2017, 8, 713. [Google Scholar] [CrossRef]
- Kelly, C.; Gundogdu, O.; Pircalabioru, G.; Cean, A.; Scates, P.; Linton, M.; Pinkerton, L.; Magowan, E.; Stef, L.; Simiz, E.; et al. The In Vitro and In Vivo Effect of Carvacrol in Preventing Campylobacter Infection, Colonization and in Improving Productivity of Chicken Broilers. Foodborne Pathog. Dis. 2017, 14, 341–349. [Google Scholar] [CrossRef]
- Mousavi, S.; Schmidt, A.M.; Escher, U.; Kittler, S.; Kehrenberg, C.; Thunhorst, E.; Bereswill, S.; Heimesaat, M.M. Carvacrol ameliorates acute campylobacteriosis in a clinical murine infection model. Gut Pathog. 2020, 12, 2. [Google Scholar] [CrossRef]
- Wagle, B.R.; Donoghue, A.M.; Shrestha, S.; Upadhyaya, I.; Arsi, K.; Gupta, A.; Liyanage, R.; Rath, N.C.; Donoghue, D.J.; Upadhyay, A. Carvacrol attenuates Campylobacter jejuni colonization factors and proteome critical for persistence in the chicken gut. Poult. Sci. 2020, 99, 4566–4577. [Google Scholar] [CrossRef]
- Wei, H.K.; Xue, H.X.; Zhou, Z.X.; Peng, J. A carvacrol-thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Kang, G.D.; Lim, S.; Kim, D.H. Oleanolic acid ameliorates dextran sodium sulfate-induced colitis in mice by restoring the balance of Th17/Treg cells and inhibiting NF-κB signaling pathway. Int. Immunopharmacol. 2015, 29, 393–400. [Google Scholar] [CrossRef]
- Chun, J.; Lee, C.; Hwang, S.W.; Im, J.P.; Kim, J.S. Ursolic acid inhibits nuclear factor-κB signaling in intestinal epithelial cells and macrophages, and attenuates experimental colitis in mice. Life Sci. 2014, 110, 23–34. [Google Scholar] [CrossRef]
- Liu, B.; Piao, X.; Guo, L.; Liu, S.; Chai, F.; Gao, L. Ursolic acid protects against ulcerative colitis via anti-inflammatory and antioxidant effects in mice. Mol. Med. Rep. 2016, 13, 4779–4785. [Google Scholar] [CrossRef]
- Sheng, Q.; Li, F.; Chen, G.; Li, J.; Li, J.; Wang, Y.; Lu, Y.; Li, Q.; Li, M.; Chai, K. Ursolic Acid Regulates Intestinal Microbiota and Inflammatory Cell Infiltration to Prevent Ulcerative Colitis. J. Immunol. Res. 2021, 2021, 6679316. [Google Scholar] [CrossRef]
- Hao, W.; Kwek, E.; He, Z.; Zhu, H.; Liu, J.; Zhao, Y.; Ma, K.Y.; He, W.S.; Chen, Z.Y. Ursolic acid alleviates hypercholesterolemia and modulates the gut microbiota in hamsters. Food Funct. 2020, 11, 6091–6103. [Google Scholar] [CrossRef]
- Wan, S.Z.; Liu, C.; Huang, C.K.; Luo, F.Y.; Zhu, X. Ursolic Acid Improves Intestinal Damage and Bacterial Dysbiosis in Liver Fibrosis Mice. Front. Pharmacol. 2019, 10, 1321. [Google Scholar] [CrossRef]
- Wan, S.; Huang, C.; Wang, A.; Zhu, X. Ursolic acid improves the bacterial community mapping of the intestinal tract in liver fibrosis mice. PeerJ 2020, 8, e9050. [Google Scholar] [CrossRef]
- Peng, F.; Zhang, H.; He, X.; Song, Z. Effects of Ursolic Acid on Intestinal Health and Gut Bacteria Antibiotic Resistance in Mice. Front. Physiol. 2021, 12, 650190. [Google Scholar] [CrossRef]
- Dong, N.; Xue, C.; Zhang, L.; Zhang, T.; Wang, C.; Bi, C.; Shan, A. Oleanolic acid enhances tight junctions and ameliorates inflammation in Salmonella typhimurium-induced diarrhea in mice via the TLR4/NF-κB and MAPK pathway. Food Funct. 2020, 11, 1122–1132. [Google Scholar] [CrossRef]
- Jain, A.K.; Wen, J.X.; Blomenkamp, K.S.; Arora, S.; Blaufuss, T.A.; Rodrigues, J.; Long, J.P.; Neuschwander-Tetri, B.A.; Teckman, J.H. Oleanolic Acid Improves Gut Atrophy Induced by Parenteral Nutrition. JPEN J. Parenter. Enter. Nutr. 2016, 40, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Li, Y.; Lv, H.; Zhang, L.; Bi, C.; Dong, N.; Shan, A.; Wang, J. Oleanolic Acid Targets the Gut-Liver Axis to Alleviate Metabolic Disorders and Hepatic Steatosis. J. Agric. Food Chem. 2021, 69, 7884–7897. [Google Scholar] [CrossRef]
- Xue, C.; Lv, H.; Li, Y.; Dong, N.; Wang, Y.; Zhou, J.; Shi, B.; Shan, A. Oleanolic acid reshapes the gut microbiota and alters immune-related gene expression of intestinal epithelial cells. J. Sci. Food Agric. 2021, 102, 764–773. [Google Scholar] [CrossRef]
- Farag, M.A.; Abdelwareth, A.; Sallam, I.E.; El Shorbagi, M.; Jehmlich, N.; Fritz-Wallace, K.; Schape, S.S.; Rolle-Kampczyk, U.; Ehrlich, A.; Wessjohann, L.A.; et al. Metabolomics reveals impact of seven functional foods on metabolic pathways in a gut microbiota model. J. Adv. Res. 2020, 23, 47–59. [Google Scholar] [CrossRef]
- Doğan, A.; Çelik, İ. Healing effects of sumac (Rhus coriaria) in streptozotocin-induced diabetic rats. Pharm. Biol. 2016, 54, 2092–2102. [Google Scholar] [CrossRef]
- Isik, S.; Tayman, C.; Cakir, U.; Koyuncu, I.; Taskin Turkmenoglu, T.; Cakir, E. Sumac (Rhus coriaria) for the prevention and treatment of necrotizing enterocolitis. J. Food Biochem. 2019, 43, e13068. [Google Scholar] [CrossRef]
- Jang, S.E.; Hyam, S.R.; Jeong, J.J.; Han, M.J.; Kim, D.H. Penta-O-galloyl-β-D-glucose ameliorates inflammation by inhibiting MyD88/NF-κB and MyD88/MAPK signalling pathways. Br. J. Pharmacol. 2013, 170, 1078–1091. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Gao, T.; Li, L.; Chen, Y.; Xiao, D.; Liu, W.; Zou, B.; Lu, B.; Tian, X.; et al. A holistic view of gallic acid-induced attenuation in colitis based on microbiome-metabolomics analysis. Food Funct. 2019, 10, 4046–4061. [Google Scholar] [CrossRef]
- Shree, A.; Islam, J.; Vafa, A.; Mohammad Afzal, S.; Sultana, S. Gallic acid prevents 1, 2-Dimethylhydrazine induced colon inflammation, toxicity, mucin depletion, and goblet cell disintegration. Environ. Toxicol. 2020, 35, 652–664. [Google Scholar] [CrossRef]
- Song, J.L.; Choi, J.H.; Seo, J.H.; Park, K.Y. Fermented Ganjangs (Soy Sauce and Sesame Sauce) Attenuates Colonic Carcinogenesis in Azoxymethane/Dextran Sodium Sulfate-Treated C57BL/6J Mice. J. Med. Food 2018, 21, 905–914. [Google Scholar] [CrossRef]
- Yuan, T.; Chu, C.; Shi, R.; Cui, T.; Zhang, X.; Zhao, Y.; Shi, X.; Hui, Y.; Pan, J.; Qian, R.; et al. ApoE-Dependent Protective Effects of Sesamol on High-Fat Diet-Induced Behavioral Disorders: Regulation of the Microbiome-Gut-Brain Axis. J. Agric. Food Chem. 2019, 67, 6190–6201. [Google Scholar] [CrossRef]
- Ren, B.; Yuan, T.; Zhang, X.; Wang, L.; Pan, J.; Liu, Y.; Zhao, B.; Zhao, W.; Liu, Z.; Liu, X. Protective Effects of Sesamol on Systemic Inflammation and Cognitive Impairment in Aging Mice. J. Agric. Food Chem. 2020, 68, 3099–3111. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, T.; Xi, Y.; Li, L.; Mo, F.; Liu, X.; Liu, Z.; Gao, J.M.; Yuan, T. Sesamol Attenuates Amyloid Peptide Accumulation and Cognitive Deficits in APP/PS1 Mice: The Mediating Role of the Gut-Brain Axis. J. Agric. Food Chem. 2021, 69, 12717–12729. [Google Scholar] [CrossRef]
- Periasamy, S.; Hsu, D.Z.; Chandrasekaran, V.R.; Liu, M.Y. Sesame oil accelerates healing of 2,4,6-trinitrobenzenesulfonic acid-induced acute colitis by attenuating inflammation and fibrosis. JPEN J. Parenter. Enter. Nutr. 2013, 37, 674–682. [Google Scholar] [CrossRef]
- Kondamudi, P.K.; Kovelamudi, H.; Mathew, G.; Nayak, P.G.; Rao, M.C.; Shenoy, R.R. Investigation of sesamol on myeloperoxidase and colon morphology in acetic acid-induced inflammatory bowel disorder in albino rats. Sci. World J. 2014, 2014, 802701. [Google Scholar] [CrossRef]
- Khan, S.; Kumar, A.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Protective effect of sesamol against ⁶⁰Co γ-ray-induced hematopoietic and gastrointestinal injury in C57BL/6 male mice. Free Radic. Res. 2015, 49, 1344–1361. [Google Scholar] [CrossRef]
- Dafni, A.; Böck, B. Medicinal plants of the Bible-revisited. J. Ethnobiol. Ethnomed. 2019, 15, 57. [Google Scholar] [CrossRef]
- Carballeda-Sangiao, N.; Sánchez-Alonso, I.; Navas, A.; Arcos, S.C.; de Palencia, P.F.; Careche, M.; González-Muñoz, M. Anisakis simplex products impair intestinal epithelial barrier function and occludin and zonula occludens-1 localisation in differentiated Caco-2 cells. PLoS Negl. Trop. Dis. 2020, 14, e0008462. [Google Scholar] [CrossRef]
- Polimeno, L.; Loiacono, M.; Pesetti, B.; Mallamaci, R.; Mastrodonato, M.; Azzarone, A.; Annoscia, E.; Gatti, F.; Amoruso, A.; Ventura, M.T. Anisakiasis, an underestimated infection: Effect on intestinal permeability of Anisakis simplex-sensitized patients. Foodborne Pathog. Dis. 2010, 7, 809–814. [Google Scholar] [CrossRef]
- Daouk, R.K.; Dagher, S.M.; Sattout, E.J. Antifungal Activity of the Essential Oil of Origanum syriacum L. J. Food Prot. 1995, 58, 1147–1149. [Google Scholar] [CrossRef]
- Kiliç, T. Analysis of essential oil composition of Thymbra spicata var. spicata: Antifungal, antibacterial and antimycobacterial activities. Z. Nat. C J. Biosci. 2006, 61, 324–328. [Google Scholar] [CrossRef]
- Karimi, M.; Naserifar, R.; Abbasi, N.; Abdi, J.; Maspi, N. In vitro antileishmanial activity of hydroalcoholic Thymbra spicata extract on Leishmania major promastigotes. Ann. Parasitol. 2021, 67, 237–241. [Google Scholar] [CrossRef]
- Haroun, M.F.; Al-Kayali, R.S. Synergistic effect of Thymbra spicata L. extracts with antibiotics against multidrug-resistant Staphylococcus aureus and Klebsiella pneumoniae strains. Iran. J. Basic Med. Sci. 2016, 19, 1193–1200. [Google Scholar]
- Hawrelak, J.A.; Cattley, T.; Myers, S.P. Essential oils in the treatment of intestinal dysbiosis: A preliminary in vitro study. Altern. Med. Rev. 2009, 14, 380–384. [Google Scholar]
- Thapa, D.; Louis, P.; Losa, R.; Zweifel, B.; Wallace, R.J. Essential oils have different effects on human pathogenic and commensal bacteria in mixed faecal fermentations compared with pure cultures. Microbiology 2015, 161, 441–449. [Google Scholar] [CrossRef]
- Logan, A.C.; Beaulne, T.M. The treatment of small intestinal bacterial overgrowth with enteric-coated peppermint oil: A case report. Altern. Med. Rev. 2002, 7, 410–417. [Google Scholar]
- Ceppa, F.; Faccenda, F.; De Filippo, C.; Albanese, D.; Pindo, M.; Martelli, R.; Marconi, P.; Lunelli, F.; Fava, F.; Parisi, G. Influence of essential oils in diet and life-stage on gut microbiota and fillet quality of rainbow trout (Oncorhynchus mykiss). Int. J. Food Sci. Nutr. 2018, 69, 318–333. [Google Scholar] [CrossRef]
- Mooyottu, S.; Kollanoor-Johny, A.; Flock, G.; Bouillaut, L.; Upadhyay, A.; Sonenshein, A.L.; Venkitanarayanan, K. Carvacrol and trans-cinnamaldehyde reduce Clostridium difficile toxin production and cytotoxicity in vitro. Int. J. Mol. Sci. 2014, 15, 4415–4430. [Google Scholar] [CrossRef]
- Burt, S.A.; Adolfse, S.J.; Ahad, D.S.; Tersteeg-Zijderveld, M.H.; Jongerius-Gortemaker, B.G.; Post, J.A.; Brüggemann, H.; Santos, R.R. Cinnamaldehyde, Carvacrol and Organic Acids Affect Gene Expression of Selected Oxidative Stress and Inflammation Markers in IPEC-J2 Cells Exposed to Salmonella typhimurium. Phytother. Res. PTR 2016, 30, 1988–2000. [Google Scholar] [CrossRef]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef]
- Hsu, J.E.; Lo, S.H.; Lin, Y.Y.; Wang, H.T.; Chen, C.Y. Effects of essential oil mixtures on nitrogen metabolism and odor emission via in vitro simulated digestion and in vivo growing pig experiments. J. Sci. Food Agric. 2021, 102, 1939–1947. [Google Scholar] [CrossRef]
- Firmino, J.P.; Vallejos-Vidal, E.; Balebona, M.C.; Ramayo-Caldas, Y.; Cerezo, I.M.; Salomón, R.; Tort, L.; Estevez, A.; Moriñigo, M.; Reyes-López, F.E.; et al. Diet, Immunity, and Microbiota Interactions: An Integrative Analysis of the Intestine Transcriptional Response and Microbiota Modulation in Gilthead Seabream (Sparus aurata) Fed an Essential Oils-Based Functional Diet. Front. Immunol. 2021, 12, 625297. [Google Scholar] [CrossRef]
- Liu, J. Oleanolic acid and ursolic acid: Research perspectives. J. Ethnopharmacol. 2005, 100, 92–94. [Google Scholar] [CrossRef]
- Szakiel, A.; Ruszkowski, D.; Grudniak, A.; Kurek, A.; Wolska, K.I.; Doligalska, M.; Janiszowska, W. Antibacterial and antiparasitic activity of oleanolic acid and its glycosides isolated from marigold (Calendula officinalis). Planta Med. 2008, 74, 1709–1715. [Google Scholar] [CrossRef]
- Chen, J.C.; Chang, Y.S.; Wu, S.L.; Chao, D.C.; Chang, C.S.; Li, C.C.; Ho, T.Y.; Hsiang, C.Y. Inhibition of Escherichia coli heat-labile enterotoxin-induced diarrhea by Chaenomeles speciosa. J. Ethnopharmacol. 2007, 113, 233–239. [Google Scholar] [CrossRef]
- Zhang, W.; Gan, D.; Jian, J.; Huang, C.; Luo, F.; Wan, S.; Jiang, M.; Wan, Y.; Wang, A.; Li, B.; et al. Protective Effect of Ursolic Acid on the Intestinal Mucosal Barrier in a Rat Model of Liver Fibrosis. Front. Physiol. 2019, 10, 956. [Google Scholar] [CrossRef]
- Li, Y.; Matsuda, H.; Yoshikawa, M. Effects of oleanolic acid glycosides on gastrointestinal transit and ileus in mice. Bioorg. Med. Chem. 1999, 7, 1201–1205. [Google Scholar] [CrossRef]
- Andersson, D.; Cheng, Y.; Duan, R.D. Ursolic acid inhibits the formation of aberrant crypt foci and affects colonic sphingomyelin hydrolyzing enzymes in azoxymethane-treated rats. J. Cancer Res. Clin. Oncol. 2008, 134, 101–107. [Google Scholar] [CrossRef]
- Zhang, P.; Cheng, Y.; Duan, R.D. Ursolic acid inhibits acid sphingomyelinase in intestinal cells. Phytother. Res. PTR 2013, 27, 173–178. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, H.G.; Im, H.J.; Lee, J.S.; Lee, S.B.; Kim, W.Y.; Lee, H.W.; Lee, S.K.; Byun, C.K.; Son, C.G. Antiemetic and Myeloprotective Effects of Rhus verniciflua Stoke in a Cisplatin-Induced Rat Model. Evid. Based Complement. Altern. Med. 2017, 2017, 9830342. [Google Scholar] [CrossRef]
- Barka, Z.B.; Grintzalis, K.; Polet, M.; Heude, C.; Sommer, U.; Ben Miled, H.; Rhouma, K.B.; Mohsen, S.; Tebourbi, O.; Schneider, Y.J. A combination of NMR and liquid chromatography to characterize the protective effects of Rhus tripartita extracts on ethanol-induced toxicity and inflammation on intestinal cells. J. Pharm. Biomed. Anal. 2018, 150, 347–354. [Google Scholar] [CrossRef]
- Barka, Z.B.; Aouadhi, C.; Tlili, M.; Alimi, H.; Miled, H.B.; Rhouma, K.B.; Sakly, M.; Ksouri, R.; Schneider, Y.J.; Maaroufi, A.; et al. Evaluation of the anti-diarrheal activity of the hydromethanolic root extract of Rhus tripartita (Ucria) (Anacardiacae). Biomed. Pharmacother. 2016, 83, 827–834. [Google Scholar] [CrossRef]
- Tangpu, V.; Yadav, A.K. Antidiarrhoeal activity of Rhus javanica ripen fruit extract in albino mice. Fitoterapia 2004, 75, 39–44. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, O.; Ma, N.; Yi, J.; Mi, H.; Cai, S. The preventive effect and underlying mechanism of Rhus chinensis Mill. fruits on dextran sulphate sodium-induced ulcerative colitis in mice. Food Funct. 2021, 12, 9965–9978. [Google Scholar] [CrossRef]
- Lee, Y.M.; Park, J.P.; Lim, K.T.; Lee, S.J. Intestinal epithelial cell apoptosis due to a hemolytic toxin from Vibrio vulnificus and protection by a 36 kDa glycoprotein from Rhus verniciflua Stokes. Food Chem. Toxicol. 2019, 125, 46–54. [Google Scholar] [CrossRef]
- Mahernia, S.; Bagherzadeh, K.; Mojab, F.; Amanlou, M. Urease Inhibitory Activities of some Commonly Consumed Herbal Medicines. Iran. J. Pharm. Res. 2015, 14, 943–947. [Google Scholar]
- Kim, H.; Venancio, V.P.; Fang, C.; Dupont, A.W.; Talcott, S.T.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr. Res. 2020, 75, 85–94. [Google Scholar] [CrossRef]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Hidalgo, M.; Martin-Santamaria, S.; Recio, I.; Sanchez-Moreno, C.; de Pascual-Teresa, B.; Rimbach, G.; de Pascual-Teresa, S. Potential anti-inflammatory, anti-adhesive, anti/estrogenic, and angiotensin-converting enzyme inhibitory activities of anthocyanins and their gut metabolites. Genes Nutr. 2012, 7, 295–306. [Google Scholar] [CrossRef]
- Kiss, A.K.; Piwowarski, J.P. Ellagitannins, Gallotannins and their Metabolites—The Contribution to the Anti-Inflammatory Effect of Food Products and Medicinal Plants. Curr. Med. Chem. 2018, 25, 4946–4967. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y.; et al. Impact of Gallic Acid on Gut Health: Focus on the Gut Microbiome, Immune Response, and Mechanisms of Action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef]
- Shimizu, S.; Fujii, G.; Takahashi, M.; Nakanishi, R.; Komiya, M.; Shimura, M.; Noma, N.; Onuma, W.; Terasaki, M.; Yano, T.; et al. Sesamol suppresses cyclooxygenase-2 transcriptional activity in colon cancer cells and modifies intestinal polyp development in Apc (Min/+) mice. J. Clin. Biochem. Nutr. 2014, 54, 95–101. [Google Scholar] [CrossRef]
- Sheng, H.; Hirose, Y.; Hata, K.; Zheng, Q.; Kuno, T.; Asano, N.; Yamada, Y.; Hara, A.; Osawa, T.; Mori, H. Modifying effect of dietary sesaminol glucosides on the formation of azoxymethane-induced premalignant lesions of rat colon. Cancer Lett. 2007, 246, 63–68. [Google Scholar] [CrossRef]
- Chavali, S.R.; Utsunomiya, T.; Forse, R.A. Increased survival after cecal ligation and puncture in mice consuming diets enriched with sesame seed oil. Crit. Care Med. 2001, 29, 140–143. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Chen, K.T.; Li, Y.H.; Chuang, Y.C.; Liu, M.Y. Sesamol delays mortality and attenuates hepatic injury after cecal ligation and puncture in rats: Role of oxidative stress. Shock 2006, 25, 528–532. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Li, Y.H.; Chien, S.P.; Liu, M.Y. Effects of sesame oil on oxidative stress and hepatic injury after cecal ligation and puncture in rats. Shock 2004, 21, 466–469. [Google Scholar] [CrossRef]
- Hsu, D.Z.; Chien, S.P.; Li, Y.H.; Chuang, Y.C.; Chang, Y.C.; Liu, M.Y. Sesame oil attenuates hepatic lipid peroxidation by inhibiting nitric oxide and superoxide anion generation in septic rats. JPEN J. Parenter. Enter. Nutr. 2008, 32, 154–159. [Google Scholar] [CrossRef]
- Rezaeipour, V.; Barsalani, A.; Abdullahpour, R. Effects of phytase supplementation on growth performance, jejunum morphology, liver health, and serum metabolites of Japanese quails fed sesame (Sesamum indicum) meal-based diets containing graded levels of protein. Trop. Anim. Health Prod. 2016, 48, 1141–1146. [Google Scholar] [CrossRef]
- Song, J.L.; Choi, J.H.; Seo, J.H.; Lim, Y.I.; Park, K.Y. Anti-colitic effects of kanjangs (fermented soy sauce and sesame sauce) in dextran sulfate sodium-induced colitis in mice. J. Med. Food 2014, 17, 1027–1035. [Google Scholar] [CrossRef]
- Del Bo, C.; Bernardi, S.; Cherubini, A.; Porrini, M.; Gargari, G.; Hidalgo-Liberona, N.; González-Domínguez, R.; Zamora-Ros, R.; Peron, G.; Marino, M.; et al. A polyphenol-rich dietary pattern improves intestinal permeability, evaluated as serum zonulin levels, in older subjects: The MaPLE randomised controlled trial. Clin. Nutr. 2021, 40, 3006–3018. [Google Scholar] [CrossRef]
- Luo, H.Q.; Shen, J.; Chen, C.P.; Ma, X.; Lin, C.; Ouyang, Q.; Xuan, C.X.; Liu, J.; Sun, H.B.; Liu, J. Lipid-lowering effects of oleanolic acid in hyperlipidemic patients. Chin. J. Nat. Med. 2018, 16, 339–346. [Google Scholar] [CrossRef]
- Asghari, E.; Rashidlamir, A.; Hosseini, S.R.A.; Moazzami, M.; Samarghandian, S.; Farkhondeh, T. Synergism Effects of Ursolic Acid Supplementation on the Levels of Irisin, C-reactive Protein, IL-6, and TNF-alpha During High-intensity Resistance Training in Low Activity Men. Cardiovasc. Hematol. Disord. Drug Targets 2020, 20, 138–144. [Google Scholar] [CrossRef]
- Venancio, V.P.; Kim, H.; Sirven, M.A.; Tekwe, C.D.; Honvoh, G.; Talcott, S.T.; Mertens-Talcott, S.U. Polyphenol-rich Mango (Mangifera indica L.) Ameliorate Functional Constipation Symptoms in Humans beyond Equivalent Amount of Fiber. Mol. Nutr. Food Res. 2018, 62, e1701034. [Google Scholar] [CrossRef]
- Barnes, R.C.; Kim, H.; Fang, C.; Bennett, W.; Nemec, M.; Sirven, M.A.; Suchodolski, J.S.; Deutz, N.; Britton, R.A.; Mertens-Talcott, S.U.; et al. Body Mass Index as a Determinant of Systemic Exposure to Gallotannin Metabolites during 6-Week Consumption of Mango (Mangifera indica L.) and Modulation of Intestinal Microbiota in Lean and Obese Individuals. Mol. Nutr. Food Res. 2019, 63, e1800512. [Google Scholar] [CrossRef]
- Wu, W.H.; Kang, Y.P.; Wang, N.H.; Jou, H.J.; Wang, T.A. Sesame ingestion affects sex hormones, antioxidant status, and blood lipids in postmenopausal women. J. Nutr. 2006, 136, 1270–1275. [Google Scholar] [CrossRef]
- Ji, Z.L.; Li, J.S.; Yuan, C.W.; Chen, W.D.; Zhang, Y.N.; Ju, X.T.; Tang, W.H. Therapeutic value of sesame oil in the treatment of adhesive small bowel obstruction. Am. J. Surg. 2010, 199, 160–165. [Google Scholar] [CrossRef]
- Yuan, M.; Chen, X.; Su, T.; Zhou, Y.; Sun, X. Supplementation of Kiwifruit Polyphenol Extract Attenuates High Fat Diet Induced Intestinal Barrier Damage and Inflammation via Reshaping Gut Microbiome. Front. Nutr. 2021, 8, 702157. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Peron, G.; Hidalgo-Liberona, N.; González-Domínguez, R.; Garcia-Aloy, M.; Guglielmetti, S.; Bernardi, S.; Kirkup, B.; Kroon, P.A.; Cherubini, A.; Riso, P.; et al. Exploring the Molecular Pathways Behind the Effects of Nutrients and Dietary Polyphenols on Gut Microbiota and Intestinal Permeability: A Perspective on the Potential of Metabolomics and Future Clinical Applications. J. Agric. Food Chem. 2020, 68, 1780–1789. [Google Scholar] [CrossRef]
- Mullin, G.E.; Singh, M.; Parian, A.; Weil, A.T. Integrative Gastroenterology; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Drossman, D.A.; Hasler, W.L. Rome IV-Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef]



| Plants | Sample Used | Model | Disorder | Main Funding | Refs. |
|---|---|---|---|---|---|
| O. syracium and derived compounds | EOs | In vitro Anisakis simplex Parasite | Gastrointestinal Parasite | ↑ Parasite mortality ↓ Penetration ability | [68] |
| Extract | In vivo Ethanol-induced ulcer in Swiss albino mice | Peptic ulcer | ↓ Ulcer score ↓ Gastric damage | [69] | |
| EOs | In vivo Pigs | Microbiota | ↑ Bacilli, Lactobacillales, Streptococcaceae, Veillonellaceae | [70] | |
| OEO | In vivo Pigs | Microbiota | ↑ Lactobacillaceae, Fibrobacteriaceae, Akkermansiaceae, Lachnospiraceae ↓ Enterobacteriaceae | [71] | |
| OEO | In vivo Ducks | Microbiota-performance | ↓ Coliforms, total aerobes, lactose-negative Enterobacteria | [72] | |
| RA | In vivo Ethanol-induced gastric mucosal injury in Male BALB/c mice | Peptic ulcer | ↓ Oxidative stress ↓ Inflammation ↓ Apoptosis | [73] | |
| RA-rich extract | In vivo Male BALB/c mice | Microbiota | ↑ Muribaculaceae and Ruminococcaceae families ↓ Prevotellaceae family ↑ SCFAs | [73] | |
| RA | In vivo Streptozotocin-induced diabetic rats | Diabetes-Microbiota | ↑ Actinobacteria, Bacteroides, Faecalibacterium, Lachnospiraceae, Prevotella | [74] | |
| CVL | In vivo Clostridium difficile-infected C57BL/6 mice | Gut dysbiosis-Microbiota | ↓ Diarrhea ↑ Firmicutes ↓ Proteobacteria | [75] | |
| CVL-Thymol | In vivo C. perfringens-infected broiler chickens | Necrotic enteritis | ↓ Gut lesions ↓ TLR2, TNF-α ↑ Occludin ↑ Lactobacillus crispatus, Lactobacillus agilis ↓ Lactobacillus salivarius, Lactobacillus johnsonii | [76,77] | |
| EO | In vivo Hybrid Tilapia-Germ Free Zebrafish | Immunity-Microbiota | ↓ IL-1β, IL-8 ↑ Claudin1, Occludin2 | [78] | |
| CVL | In vivo Chickens and mice In vitro Caco2 cells | Campylobacter jejuni infection (campylobacteriosis) | ↓ C. jejuni adhesion, invasion, and translocation ↑ Lactobacillus spp ↓ Inflammation ↓ Apoptosis ↓ IFN-γ, TNF, MCP-1 and IL-6 | [79,80,81,82] | |
| CVL-thymol blend | In vivo Weaning piglets | Gut disorders | ↓ Oxidative stress markers ↑ Lactobacillus genus ↓ Enterococcus genus | [83] | |
| Oleanolic acid Ursolic acid | In vivo Mice | Colitis | ↓ NF-κB and MAPK activation pathway ↓ IL-1β, TNF-α, IL-6 ↑ IL-10 ↓ Colon shortening, myeloperoxidase activity ↑ ZO-1, occludin, claudin-1 | [84,85,86,87] | |
| Ursolic acid | In vivo Hamster | Hypercholesterolemia-Gut Microbiota | ↓ Intestinal cholesterol absorption ↓ Firmicutes, Ruminococcaceae ↑ Bacteroidetes, Rikenellaceae, Bifidobacteriaceae | [88] | |
| Ursolic acid | In vivo Mice | Liver fibrosis-intestinal damage | ↓ TNF-α, MDA, LPS ↑ ZO-1, occludin ↑ Intestinal antimicrobial peptides, angiogenin-1 ↑ Lactobacillus, Bifidobacterium, Ruminiclostridium | [89,90] | |
| Ursolic acid | In vivo Mice | Antibiotic Resistance-Microbiota | ↓ TNF-α, IL-6, LPS, DAO ↑ ZO-1, occludin ↑ Lactobacillus, ↓ Burkholderiales, Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria | [91] | |
| Oleanolic acid | In vivo Mice | Diarrhea-intestinal inflammation | ↑ ZO-1, occluding ↓ Intestinal damage ↓ NF-κB and MAPK activation pathway ↓ IL-1β, TNF-α, IL-6 | [92] | |
| Oleanolic acid | In vivo Pigs | Gut atrophy | ↑ Gut mass ↑ Villous/crypt ratio ↑ TGR5 expression | [93] | |
| Oleanolic acid | In vivo Rats | MetS-intestine damage-Microbiota | ↓ LPS, DAO, d-lactate ↑ ZO-1, occluding ↓ Intestinal damage ↓ NF-κB ↓ IL-1β, TNF-α, IL-6 ↓ Firmicutes/ Bacteroidetes ratio ↑ Ruminiclostridium, Ruminococcaceae | [94,95] | |
| Rhus coriaria and derived compounds | Extract | In vitro Bacterial microbita culture | Microbiota | ↑ SCFA ↑ Fructose | [96] |
| Extract | In vivo Streptozotocin (STZ)-induced diabetic rats | Diabetes-gut damage | ↓ Blood glucose ↓ TG, TC ↓ AST, ALT, LDH, ALP, MDA | [97] | |
| extract | In vitro Rat pups | necrotizing enterocolitis | ↓ Oxidative stress markers ↓ Apoptosis ↓ Inflammation | [98] | |
| Penta-O-galloyl-β-D-glucose | In vivo Rats | Colitis | ↓ NF-κB and MAPK activation pathway ↓ IL-1β, TNF-α, IL-6 ↑ IL-10 ↓ Colon shortening, myeloperoxidase activity | [99] | |
| GA | In vivo Rats | ulcerative colitis | ↑ Lactobacillaceae, Prevotellaceae ↓ Firmicutes, Proteobacteria ↑ Carbohydrate and bile acid metabolism ↓ Amino acid metabolism | [100] | |
| GA | In vivo Rats | Colon toxicity | ↓ Oxidative stress markers ↓ Apoptosis ↓ Inflammation ↓ Goblet cell disintegration ↓ Mucin depletion ↑ GPx, GR, GST, CAT, GSH | [101] | |
| Sesame and derived compounds | Sesame sauce | In vivo Mice | Colonic Carcinogenesis | ↓ TNF-α, IL-6, IF-γ, IL-17α, iNOS, COX-2 | [102] |
| Sesamol | In vivo Mice | Intestinal integrity-Microbiota | ↑ Gut barrier integrity ↓ LPS release ↓ TNF-α, IL-6 ↑ Bacillales, Fusobacterium Lactococus ↑ SCFAs | [103] | |
| Sesamol | In vivo Mice | Aging-systemic inflammation | ↑ Gut barrier integrity ↓ LPS release ↑ Bifidobacterium, Akkermansia ↓ Clostridium | [104] | |
| Sesamol | In vivo Mice | Alzheimer-Intestinal Integrity-Microbiota | ↑ Gut barrier integrity ↓ LPS release ↑ Gut barrier integrity ↓ LPS release ↓ Helicobacter hepaticus, Clostridium ↑ Rikenellaceae, Bifidobacterium | [105] | |
| Sesame oil | In vivo Rats | Acute colitis | ↓ Colitis index ↓ Inflammation ↓ Fibrosis ↓ Acidic mucin ↑ Natural mucin | [106] | |
| Sesamol | In vivo Rats | Inflammatory bowel disorders | ↓ MPO, MDA. NO | [107] | |
| Sesamol | In vivo Mice | Gastrointestinal injury | ↓ Mortality ↓ Lipid peroxidation ↓ Apoptosis ↑ Regeneration of crypt cells ↓ Gut bacteria translocation | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, M.; Abdallah, H.; Razuka-Ebela, D.; Calasso, M.; De Angelis, M.; Portincasa, P. The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications. Antioxidants 2023, 12, 426. https://doi.org/10.3390/antiox12020426
Khalil M, Abdallah H, Razuka-Ebela D, Calasso M, De Angelis M, Portincasa P. The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications. Antioxidants. 2023; 12(2):426. https://doi.org/10.3390/antiox12020426
Chicago/Turabian StyleKhalil, Mohamad, Hala Abdallah, Danute Razuka-Ebela, Maria Calasso, Maria De Angelis, and Piero Portincasa. 2023. "The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications" Antioxidants 12, no. 2: 426. https://doi.org/10.3390/antiox12020426
APA StyleKhalil, M., Abdallah, H., Razuka-Ebela, D., Calasso, M., De Angelis, M., & Portincasa, P. (2023). The Impact of Za’atar Antioxidant Compounds on the Gut Microbiota and Gastrointestinal Disorders: Insights for Future Clinical Applications. Antioxidants, 12(2), 426. https://doi.org/10.3390/antiox12020426