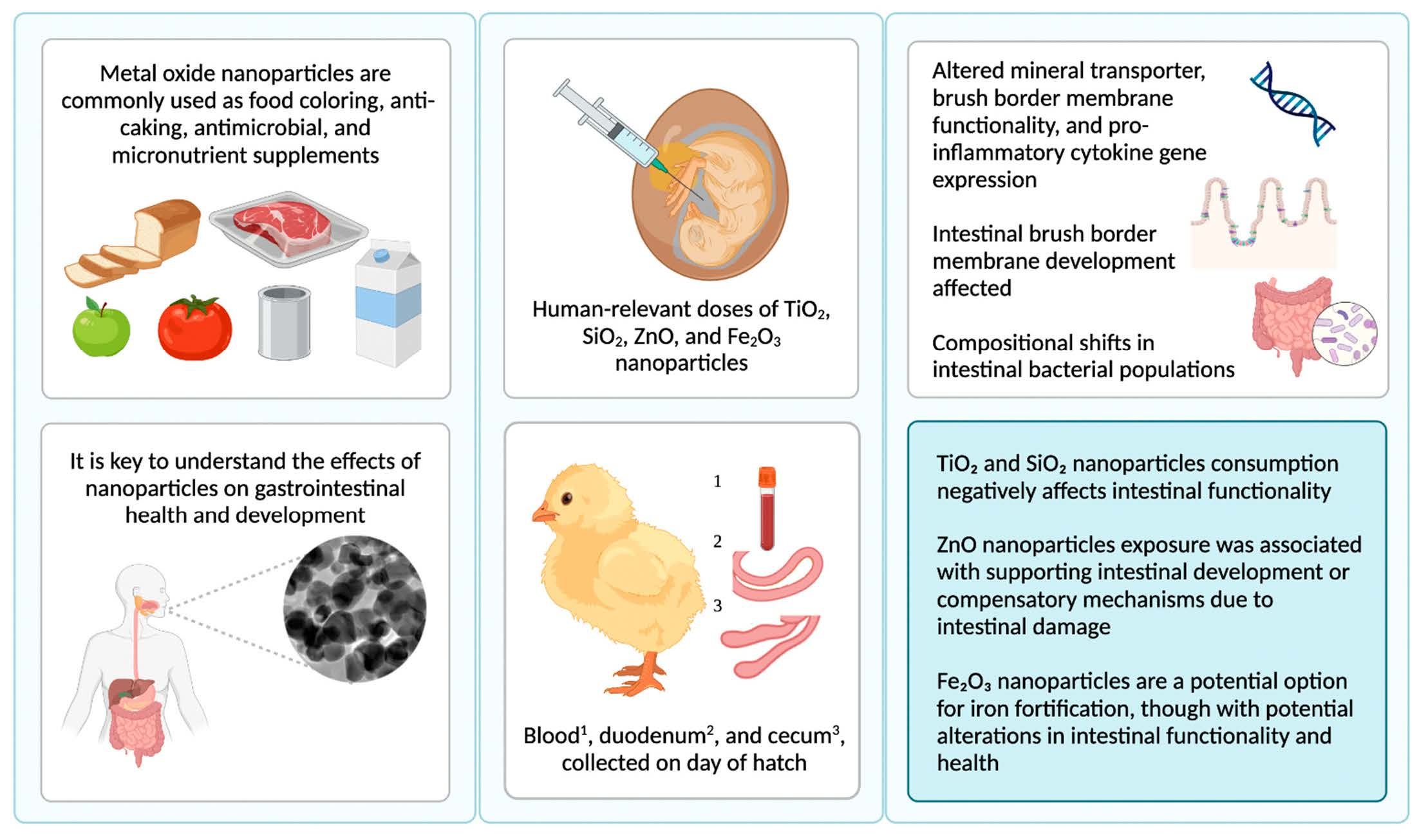
1. Introduction
Nanoparticles (NP) are commonly ingested due to their widespread usage in the food and agriculture industries, and their impact on gastrointestinal (GI) development is not well understood [
1,
2,
3,
4]. Within the food industry, NP are utilized due to their unique properties in enhancing food texture, flavor, color, consistency, stability, and preservation (antibacterial activity). Further, NP have been utilized to enhance nutrient bioavailability, where Fe
2O
3 and ZnO NP have been found to have higher relative bioavailability, faster dissociation, and minimal effects on organoleptic properties when compared with conventional fortificants [
5,
6]. NP are also found in food packaging applications to improve packaging flexibility and gas barrier properties and thus can migrate into the food matrix [
7,
8,
9]. In agriculture, NP are used for developing novel agrochemicals (i.e., fertilizers and pesticides) and sensors for the identification of nutrients or contaminants within food products or water [
10,
11]. Numerous studies have previously demonstrated food-grade metal oxides in the nanometer range and detected their presence in many food consumer products [
12,
13,
14,
15]. As human exposure to NP through food, water, and the environment is inevitable, it is key to understand the effects of NP on GI health and development.
Following ingestion, NP interact with the GI tract, where GI tract biological features and NP physicochemical properties can impact the effects of NP on GI health and development [
3,
7]. The pH varies within different compartments of the GI tract, which can affect NP aggregation and surface chemistry [
16]. In addition to serving as the key site of nutrient digestion and absorption, the small intestine also serves as protective secretory and immune functions [
17]. The mucus layer, which is produced by goblet cells and is comprised of mucopolysaccharides and glycoproteins, provides a physical barrier to luminal bacteria and concentrates antibacterial peptides and proteins secreted from Paneth cells and enterocytes near the epithelium [
17,
18]. Additionally, this mucin houses intestinal microbiota, which contributes to the digestion of minerals and interacts with epithelial cells to maintain an effective gut barrier [
19,
20,
21]. Potential outcomes of NP ingestion include absorption, by which NP can gain access to blood and thus to other organs, deposition on mucosa, and gut microbiota modulation, which have critical roles in maintaining normal gut physiologic, metabolic, and immune functions [
22,
23]. Accordingly, the consequences of NP ingestion from food sources on GI functions require further scrutiny, and the evaluation of NP must consider the effects of this perturbation on microbiota composition, intestinal functionality, and inflammatory status.
Many food additive NP, such as TiO
2, SiO
2, ZnO, and Fe
2O
3 NP, have been reported to differentially behave and interact with microorganisms, mainly due to their size-related physicochemical properties, agglomeration, and aggregation level, as well as their dissolution rate (ion release) [
24,
25,
26]. Although each NP presents different molecular compositions, similar toxicological strategies have been hypothesized and described according to each NP type. Reactive oxygen species (ROS)-mediated toxicity is believed to be the primary mechanism of in vitro NP cytotoxicity, including TiO
2, SiO
2, ZnO, and Fe
2O
3 NP, and in some cases, has also been reported to be associated with DNA damage and genotoxicity [
27,
28,
29]. However, the disturbance of cell membrane integrity due to mechanical interactions between NP clusters may be another major mechanism of TiO
2 and SiO
2 cytotoxicity where these types of food-grade NP altered the distribution of tight junction proteins due to oxidative stress-induced changes [
30,
31,
32]. ZnO and Fe
2O
3 NP cytotoxic mechanisms have also been closely related to their dissolution in the media. ZnO and Fe
2O
3 are amphoteric molecules, and as such, can behave differentially depending on the pH of the solution and are more prone to the release of ions in acidic solutions [
7]. Several studies have proposed the release of Zn
2+ in the media as a key cytotoxic and antibacterial mechanism for ZnO NP, which has a significant effect on amino acid metabolism, enzyme system disruption, and inactive transport inhibition [
33].
Our previous work with an in vitro model of the intestinal epithelium has shown that exposure to TiO
2, SiO
2, ZnO, and Fe
2O
3 NP changes the functionality of intestinal epithelial cells and causes inflammation [
7,
14,
34,
35,
36]. TiO
2 NP have been shown to negatively impact the mucus layer by decreasing mucin thickness and increasing neutral and acidic mucins when in the presence of
Escherichia coli and
Lacticaseibacillus rhamnosus, leading to potential pathological conditions [
36]. Guo et al. (2017) demonstrated that TiO
2 NP could significantly alter brush border membrane (BBM) enzyme functionality, iron (Fe) and zinc (Zn) nutrient transport, intestinal alkaline phosphatase activity, and tight junction functionality [
35]. SiO
2 NP have been shown in vitro to increase glucose transportation, decrease intestinal nutrient absorption, significantly alter the gene expression levels of nutrient transport proteins, and induce ROS and pro-inflammatory signaling [
34]. ZnO NP were shown to likely dissolve during digestion in vitro, though it was found that ZnO NP negatively affected glucose absorption and mineral absorption by altering the cell microvilli surface [
14]. Further, our previous in vitro study found that BBM enzyme activity was modulated following exposure to in vitro digested TiO
2, SiO
2, ZnO, and Fe
2O
3 NP, and in the presence of naturally present gut microbiota,
E. coli or
L. rhamnosus, some of the BBM enzyme activities that were affected by food additive NP exposure were ameliorated due to bacterial adsorption of metal oxide NP [
7].
In the well-established in vivo
Gallus gallus model [
37,
38,
39], we previously demonstrated that physiologically relevant doses of chemical-grade TiO
2 and SiO
2 NP have the potential to negatively affect intestinal functionality and health, which is congruent with previous in vitro study findings [
40]. The
G. gallus model has previously been utilized to assess mineral bioavailability due to its sensitivity to dietary mineral content manipulation and thus can serve as a model for human dietary mineral bioavailability and absorption [
38,
39,
41,
42]. Further, there is >85% gene homology in intestinal BBM proteins involved in mineral transport between humans and
G. gallus [
43]. Finally, the
G. gallus has a complex gut microbiome, significantly and directly influenced by host genetics, the environment, and diet, with a significant resemblance in the gut microbiota of
G. gallus and humans at the phylum level [
20,
44,
45].
Our present in vivo study assessed the impact of food-grade TiO
2, SiO
2, ZnO, and Fe
2O
3 NP at human-relevant dosages on intestinal health, function, and microbial populations. Due to widespread exposure resulting from human consumption of food additive NP, this present study utilized food-grade NP to study the impact of NP exposure on GI functionality. This study is a follow-up to our previous in vivo study, where we assessed the effects of chemical-grade TiO
2, SiO
2, and ZnO NP on gut health and functionality in vivo using the embryonic stage of the
G. gallus [
40]. Based on our previous in vitro results where the physiologically relevant doses of TiO
2, SiO
2, ZnO, and Fe
2O
3 NP were found to alter the activity of most of the tested digestive enzymes in a non-toxic and non-concentration-dependent way, we focused our attention on only human-relevant dose exposures [
7,
14,
34,
35]. This study utilized the intraamniotic administration approach to assess and screen the effects of food-grade TiO
2, SiO
2, ZnO, and Fe
2O
3 NP at human-relevant dosages on BBM functionality through evaluating duodenal gene expression of biomarkers of mineral status, BBM digestive and absorptive ability, immune function, and inflammation in vivo in the
G. gallus. A secondary objective was to evaluate the effects of these food-grade metal oxide NP on cecal bacterial populations by quantifying the relative abundances of health-promoting populations (
Bifidobacterium spp. and
Lactobacillus spp.) versus those of potentially pathogenic bacteria (
E. coli and
Clostridium spp.). We hypothesize that food-grade NP will have a negative effect on BBM functionality and development and alter populations of intestinal bacteria.
4. Discussion
NP are widely used as food additives within the food industry, where TiO
2 and SiO
2 NP are commonly used as food coloring or anti-caking agents, and ZnO and Fe₃O₂ NP are added as antimicrobials and coloring agents, respectively, and can be used as micronutrient supplements. Food additive NP exposure effects on GI functionality and development are still unclear; thus, our study aimed to assess the potential effects of physiologically relevant dosages of food-grade NP ingestion in vivo. Our well-established in vivo model of
G. gallus was previously utilized to elucidate the effects of ingesting functional foods and food additives at physiologically relevant doses [
38,
39,
41,
42]. In this in vivo model, chemical-grade TiO
2 and SiO
2 NP were previously demonstrated to have the potential to negatively affect intestinal functionality and health [
40]. Due to the widespread consumption of foods containing NP, our present study builds upon our previous study of chemical-grade NP to assess the effects of commonly utilized food-grade metal oxide NP. This present study utilized the intraamniotic administration approach in the
G. gallus model to assess the effect of food-grade metal oxide NP (TiO
2, SiO
2, ZnO, and Fe
2O
3) at human-relevant doses on duodenal BBM development and functionality (gene expression and histomorphology) and the relative abundance of representative cecal microbial species immediately post-hatch. These results show that food-grade metal oxide NP delivered via intraamniotic administration affected BBM functional gene expression, intestinal development, and cecal bacterial populations.
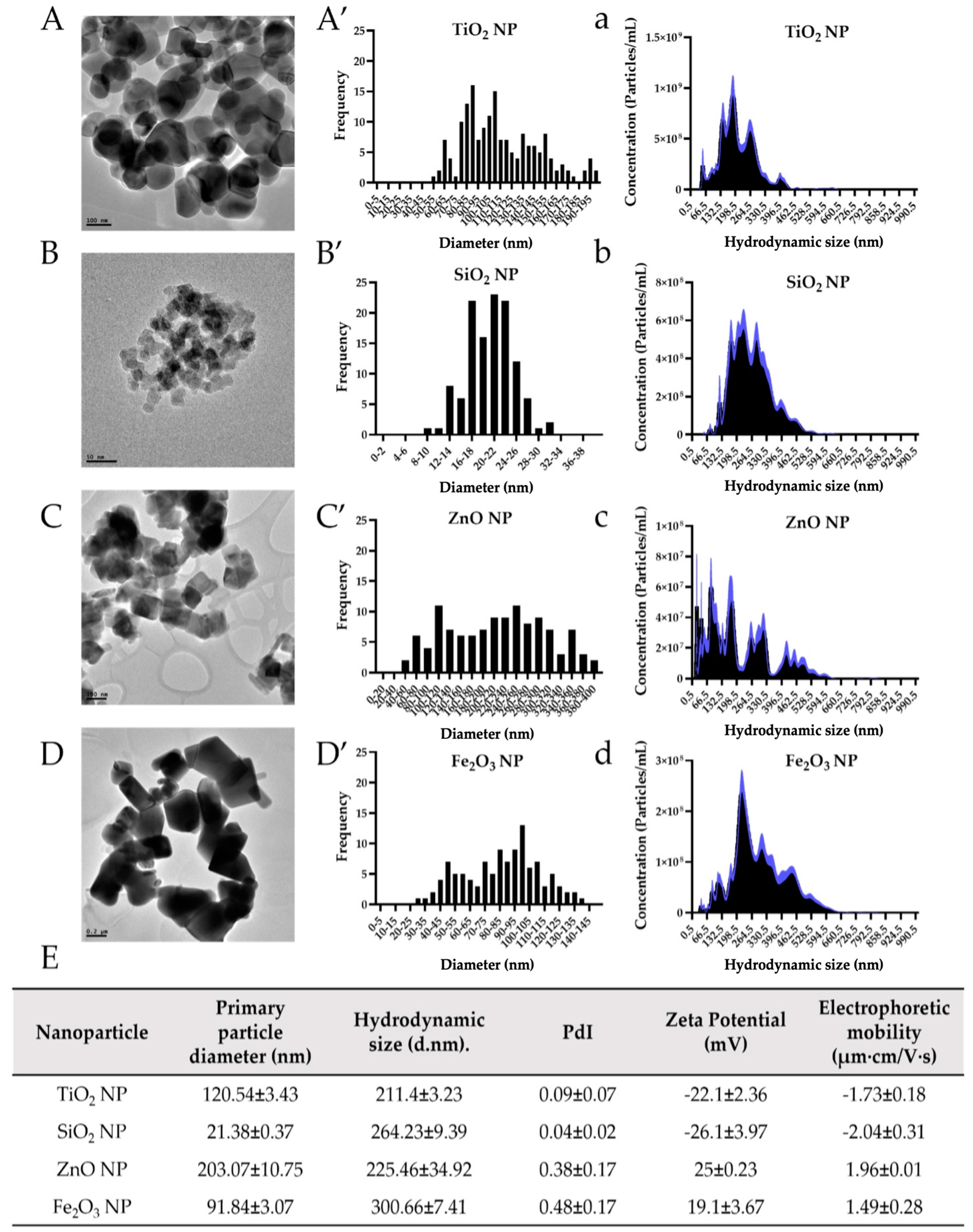
NP are well-known to be more reactive with the environment than their bulk and chemical-grade counterparts, mainly because of their size-specific physicochemical properties [
2]. Previous studies have described how different environments with different chemical properties can provoke substantial changes in NP suspension, such as erosion, ion release, agglomeration or aggregation phenomenon, protein corona formation, and attractive and repulsive forces [
64,
65]. In our previous studies, we found that more complex solutions with high protein content, such as DMEM (Dulbecco’s Modified Eagle Medium) cell culture media, fetal bovine serum (FBS), and in vitro gastric digesta as well as changes in the pH, and food matrices have a direct influence in NP physicochemical changes such as agglomeration and dissolution (ionic release) [
7,
35,
36]. These changes can drastically impact NP interaction with live organisms, with NP eventually becoming more hazardous. For example, while agglomerations of TiO
2 NP suspended in cell culture media (DMEM + FBS) caused structural damage to the in vitro epithelium, the release of ions from ZnO NP in cell culture medium impaired gut-derived bacteria (e.g.,
E. coli and
L. rhamnosus) and altered the activity of in vitro digestive brush border enzymes (e.g., intestinal alkaline phosphatase, aminopeptidase-N, and sucrase-isomaltase) [
7,
31]. An extensive investigation into the effects of in vitro digestion on several engineered NP found that while TiO
2 (0.42%) and Fe
2O
3 NP (2.27%) showed minor dissolution in simulated cascade digestion (saliva to gastric to intestinal fluids), SiO
2 and ZnO NP were partially (65.5%) and fully (100%) dissolved because of the drastic pH reduction in the stomach phase, respectively, although Si
4+ ions, but not ZnO, were found to reprecipitate in the intestinal phase [
66]. Therefore, from a toxicological point of view, NP biotransformation is an essential factor in further investigating and unveiling any potential effects from exposure. Zn and Fe ionic controls did not result in human cell cytotoxicity, statistically significant decreases in bacterial viability, or changes in brush border membrane (BBM) enzyme activity in previous in vitro work [
7]; therefore, we did not include these conditions in the current study. As many studies investigating NP environmental or biological interactions typically utilize ultrasonication procedures to prepare test suspensions from commercially available NP powders, we have followed both OECD and NIST guidelines as well as standardized protocols for NP sonication (NanoGenotox) to facilitate reproducibility among studies [
47,
67,
68]. Therefore, after the sonication procedure whereby the power delivered to the sample was 7.32 W, all four food-grade NP were fully characterized by TEM, DLS, NTA, and LDE. While the primary particle size of TiO
2, SiO
2, ZnO, and Fe
2O
3 NP was 120, 21, 203, and 92 nm, respectively, the values slightly increased when the NP size was measured in the nanopure H
2O suspension by NTA. Most metal oxide NP (e.g., TiO
2, ZnO, CeO, and Fe
2O
3 NP) form relatively large clusters or agglomerates once they are hydrated in nanopure H
2O and/or natural aqueous matrices [
64,
69]. This agglomeration is consistent with the principles of colloidal chemistry and is expected to be strongly influenced by the ionic strength and pH of aqueous environments in which NP are suspended [
70]. By DLS, French et al. (2009) showed that TiO
2 NP between 4 to 5 nm readily formed stable aggregates between 20 and 1000 nm in an aqueous suspension adjusted to a high ionic strength of 0.0165 M [
70]. In this study, food additive TiO
2, SiO
2, ZnO, and Fe
2O
3 NP were suspended and sonicated in sterile nanopure H
2O, which presented low ionic strength and low buffering capacity, and its hydrodynamic size also increased up to 211, 264, 225, and 301 nm, respectively. This could be explained by particles colliding and heating during sonication, which can influence NP properties such as morphology, surface oxide characteristics, and surface charge. Moreover, many commercially available NP powders were synthesized using high-temperature vapor phase processes, producing droplets that may coalesce fully or partially with others and form larger primary particles that are often too strong to break via sonication [
46].
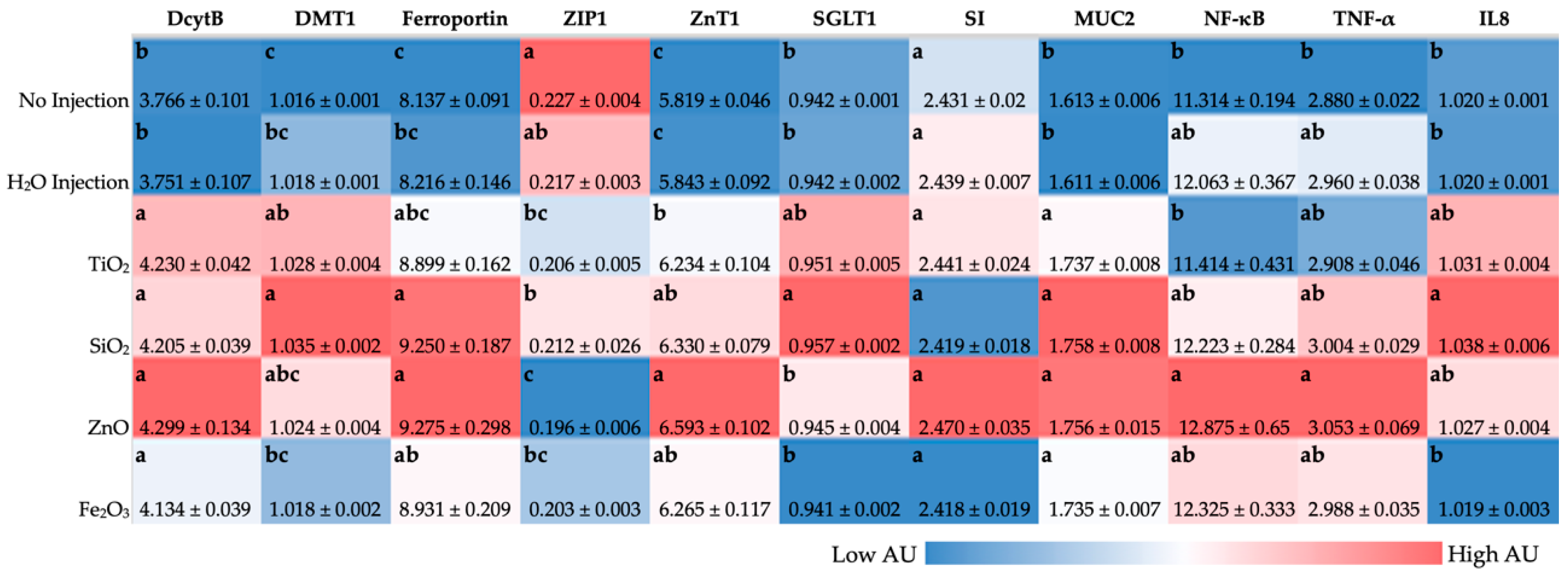
In previous intraamniotic administration studies, the downregulation of Fe duodenal transporter genes was associated with improved Fe status, where due to Fe-sufficient conditions, additional Fe transporters were not necessary as a mechanism to compensate for Fe insufficiency [
42,
53]. In line with our previous in vivo and in vitro studies, the administration of tested food-grade metal oxide NP generally upregulated DcytB, DMT1, and ferroportin, suggesting Fe insufficient luminal and basolateral conditions [
37,
40,
56]. For the expression of proteins related to cellular Zn uptake, transport, and storage, the gene expression of ZIP1 was significantly downregulated with food additive NP exposure compared with the NI control (
Figure 3). ZIP1 upregulation has been associated with Zn abundant and deficient conditions—ZIP1 is present in cellular organelles in Zn abundant media, but only on the cell membrane surface when Zn is deficient [
71,
72]. ZnT1 is located on the basolateral side of enterocytes and exports Zn
2+ from inside the enterocyte into circulation [
72,
73,
74]. Food-grade NP exposure resulted in significantly upregulated ZnT1 gene expression compared with the controls, in agreement with our previous in vivo and in vitro studies where ZnT1 upregulation was associated with NP administration [
14,
40]. ZnO NP solubilizes in acidic conditions, such as in the intestinal lumen, and can provide bioavailable Zn to the embryo, resulting in the upregulation of ZnT1 and downregulation of ZIP1 [
14,
71,
75]. Additionally, food additive ZnO NP exposure resulted in the significant upregulation of pro-inflammatory genes TNF-α and NF-κB (
Figure 3). Nuclear Factor kappa beta (NF-κB) is a transcriptional factor activated by various intra- and extra-cellular stimuli such as cytokines, reactive oxygen species (ROS), or bacterial metabolites. When NF-κB is stimulated, it translocates within the cell nucleus and is involved in various biological functions, including the release of proinflammatory mediators, such as tumor necrosis factor-alpha (TNF-α) and interleukins [
76,
77]. TNF-α is an immune cell regulator, while interleukins are a group of cytokines that play a physiological role in inflammation and a pathological role in systemic inflammatory states. On one side, changes in Zn metabolism gene expression are associated with pro-inflammatory conditions, which have previously been shown to increase Zn absorption, and on the other hand, a major target of Zn is NF-κB, where Zn is capable of modulating NF-κB activity, and NF-κB expression positively correlates with TNF-α expression [
40,
56,
78,
79,
80]. Alterations in mineral transporter expression can potentially be attributed to food-grade NP agglomeration, where NP agglomerates were observed to settle down in the BBM in an in vitro model of the GI tract, resulting in significantly fewer microvilli in the NP treatment conditions, indicative of reduced BBM development [
7]. SGLT1 gene expression was upregulated with food additive TiO
2 and SiO
2 NP exposure (
Figure 3). In previous studies, an upregulation in BBM functional gene expression suggested an improvement in
G. gallus hatchling intestinal development, digestive capabilities, and the potential for increased micronutrient (Fe or Zn) absorption [
81]; however, a previous in vivo study on chemical-grade NP exposure resulted in increased expression of BBM functional genes, which the authors posited may be indicative of intestinal development or of a compensatory mechanism to improve absorption due to intestinal damage [
40].
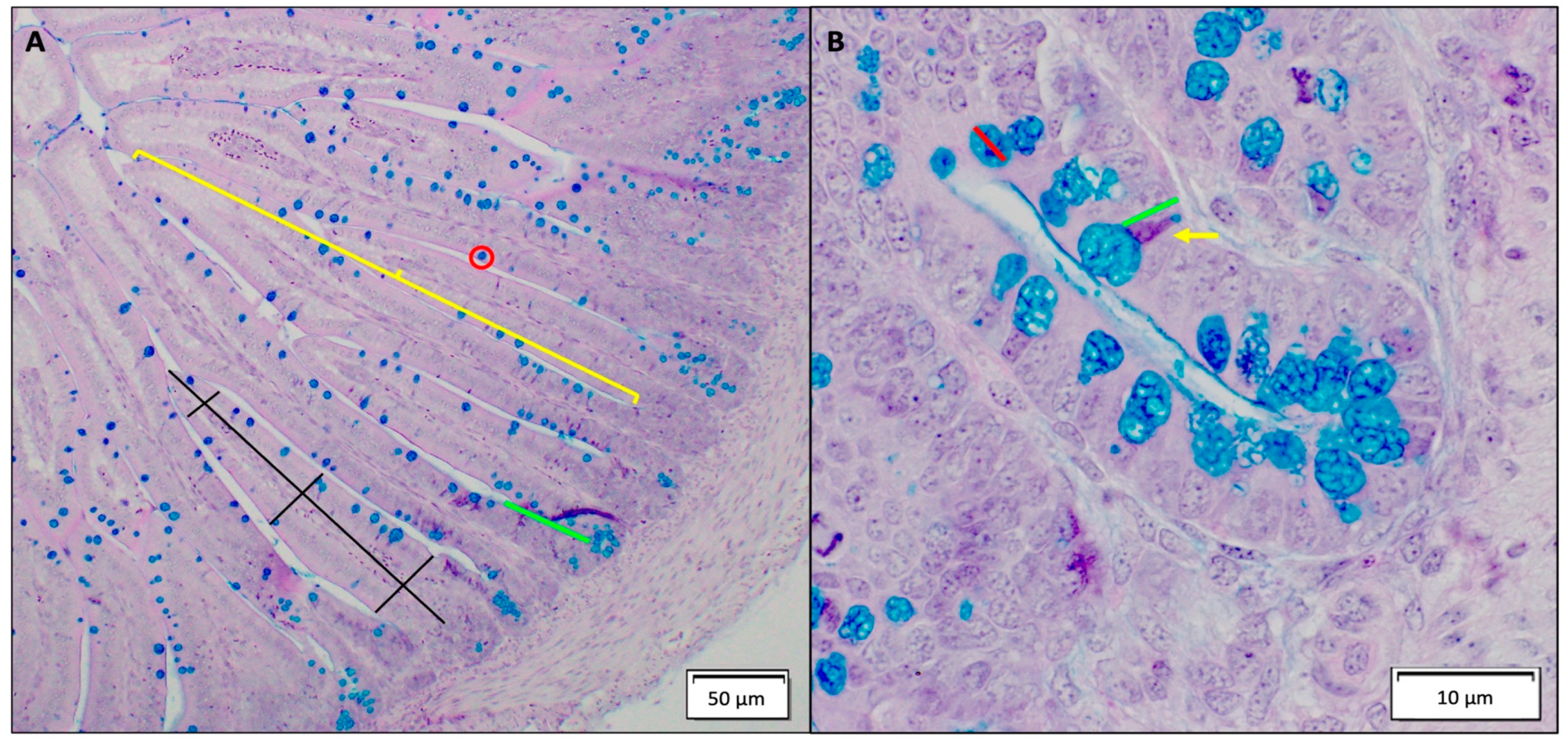
The intraamniotic administration of food-grade metal oxide NP impacted intestinal development. Exposure to food additive TiO
2, SiO
2, and ZnO NP resulted in a decrease in villus surface area (
Table 5), potentially indicative of decreased digestive enzyme and absorptive capacity [
82]. Crypt depth was found to be significantly shallower for all food additives NP-exposed groups when compared to the controls (
Table 5), where NP exposure was shown to previously be associated with increased enterocyte proliferation, either resulting from a compensatory mechanism for intestinal damage with TiO
2 and SiO
2 NP exposure, or nutritionally beneficial ZnO and Fe
2O
3 NP exposure [
83,
84,
85]. Similarly, Khajeh Bami et al. (2018) found that 42 days of ZnO NP exposure resulted in decreased crypt depth in
G. gallus [
85]. This could indicate that the broilers were utilizing food-grade ZnO and Fe
2O
3 NP for nutritional benefit, which could be supported by IL8 downregulation compared to SiO
2 and TiO
2 NP exposure (
Figure 3), which indicates a decreased level of systemic inflammation due to IL-8 mediation of the inflammatory immune response.
The major goblet cell mucins in the small and large intestines are MUC2 proteins, which are gel-forming secretory mucins that bind with intestinal bacteria and are associated with facilitating hydrolysis and absorption of nutrients [
36,
57,
86,
87]. Paneth cells secrete antimicrobial peptides into mucin produced by goblet cells, and MUC2 gene expression correlates with increased Paneth cell number and size [
88]. MUC2 gene expression was significantly upregulated with NP exposure, though a significant reduction in villi goblet cell diameter was associated with NP treatment compared with the H
2O injection control (
Figure 3). Taken together, this could be due to NP exposure-induced alterations in gut microbiota populations, resulting in NP biofilm formation on the intestinal mucosa surface, potentially hindering the ability of goblet cells to produce mucin, and where MUC2 upregulation could be a compensatory mechanism to prevent intestinal damage [
36]. A decrease in total villi goblet cells, specifically acidic villi goblet cells, was observed with food-grade TiO
2, SiO
2, and Zn NP exposure (
Table 6), indicating a decrease in synthesis and secretion of acidic luminal mucin by duodenal goblet cells [
38,
89]. With food additive Fe
2O
3 NP exposure, a significant increase in the total villi goblet cell number and a significant increase in the proportion of acidic and increase in mixture goblet cells was found in comparison with other food additive NP treated groups, with total villi acidic goblet cells maintained versus the controls (
Table 6). In addition to serving as a protective intestinal epithelial barrier, this mucin also functions as a habitat that supports probiotic bacterial proliferation, increased bacterial metabolite production, and the promotion of epithelial cell function [
86,
90].
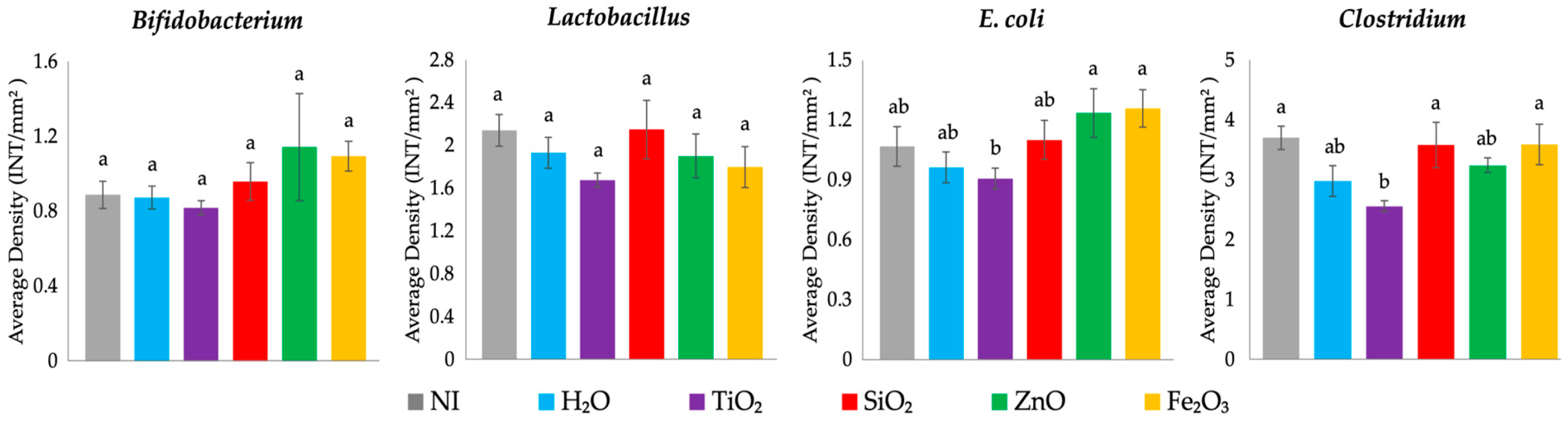
Previous in vitro and in vivo studies have shown that metal oxide exposure can lead to compositional shifts in GI bacterial populations abundance [
91]. ZnO and Fe
2O
3 NP are utilized for mineral fortification of foods, and exposure to these potentially nutritional NP resulted in an increased population of health-promoting bacteria,
Bifidobacterium, though not statistically significant. Further, food additive ZnO and Fe
2O
3 NP exposure was associated with a significant upregulation of MUC2 gene expression and increased Paneth cell number per crypt, which can be associated with providing an environment conducive to
Bifidobacterium spp. proliferation, attributed to increased acidic mucin production [
92,
93]. The potentially pathogenic
Clostridium spp. and
E. coli were increased following exposure to food-grade SiO
2, ZnO, or Fe
2O
3 NP compared with the H
2O control (
Figure 4). Gene expression resulting from food additive SiO
2, ZnO, or Fe
2O
3 NP exposure demonstrated an increase in pro-inflammatory cytokine expression, NF-κB and TNF-α (
Figure 3), and a significantly increased number of Paneth cells per crypt (
Table 8), which can be linked with the increased abundance of potentially opportunistic
Clostridium spp. and possibly pathogenic
E. coli (
Figure 4). Previous work has shown that SiO
2 and ZnO NP possessed antimicrobial properties; however, in this current study, food-grade NP were used where the dosage was too low or NP was too large to be antimicrobial [
7,
34,
40,
94]. Food additive TiO
2 NP exposure was associated with decreased populations of Gram-positive (
Bifidobacterium spp.,
Lacticaseibacillus spp., and
Clostridium spp.) and Gram-negative (
E. coli) bacteria (
Figure 4), where TiO
2 NP exposure has previously been found to lead to gut microbiota dysbiosis and decreased abundance of beneficial
Bifidobacterium and
Lacticaseibacillus genera [
95,
96]. Additional studies are warranted to assess cecal microbiota shifts, intestinal functionality, and developmental changes post-hatch and during a long-term feeding trial with human-relevant dosages of food-grade NP. The potential hazards and benefits of the tested food additive NP should be further explored to determine a relevant dose at which the potential associated risks and benefits are balanced. Further, future studies should clarify if results associated with NP exposure could be associated with the NP particles themselves or NP ions alone.




 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}