
Interaction and Redox Chemistry between Iron, Dopamine, and Alpha-Synuclein C-Terminal Peptides
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Instrumentation
2.2.1. RP-HPLC
2.2.2. Mass Spectrometry
2.2.3. HPLC-MS
2.2.4. UV-Vis Spectroscopy
2.3. General Methods
2.3.1. Peptide Synthesis
2.3.2. Effect of αS-Peptides on FeIII-DA Interaction
2.3.3. Kinetic Studies on the Oxidation of DA Promoted by FeIII-Peptide Complexes
2.3.4. Kinetic Studies on DA Oxidation Promoted by CuII-Peptide Complexes
2.3.5. Identification of αS-Peptides Modification by HPLC-MS
2.3.6. Ac-αS119–132 Enzymatic Oxidation
3. Results and Discussion
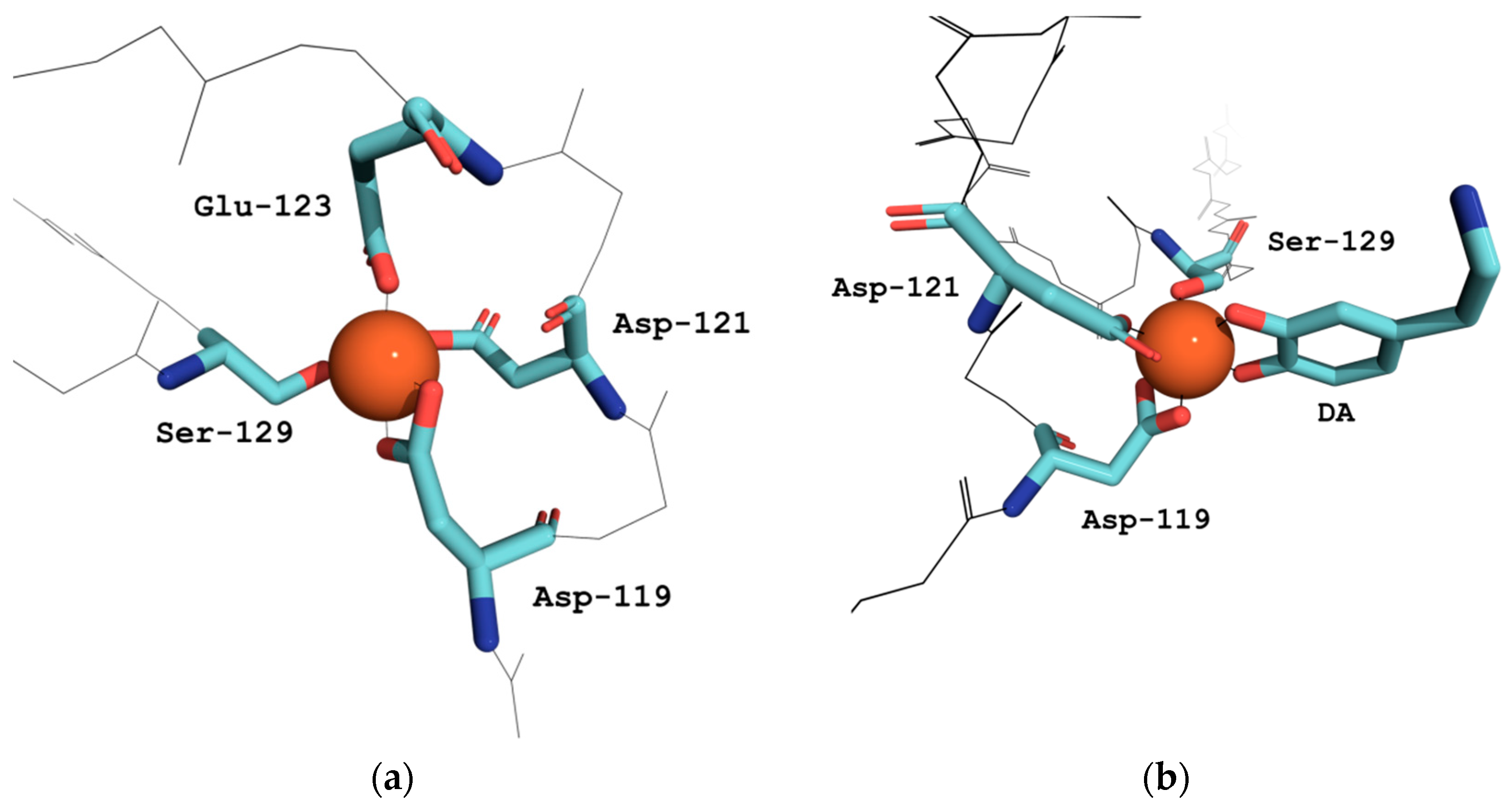
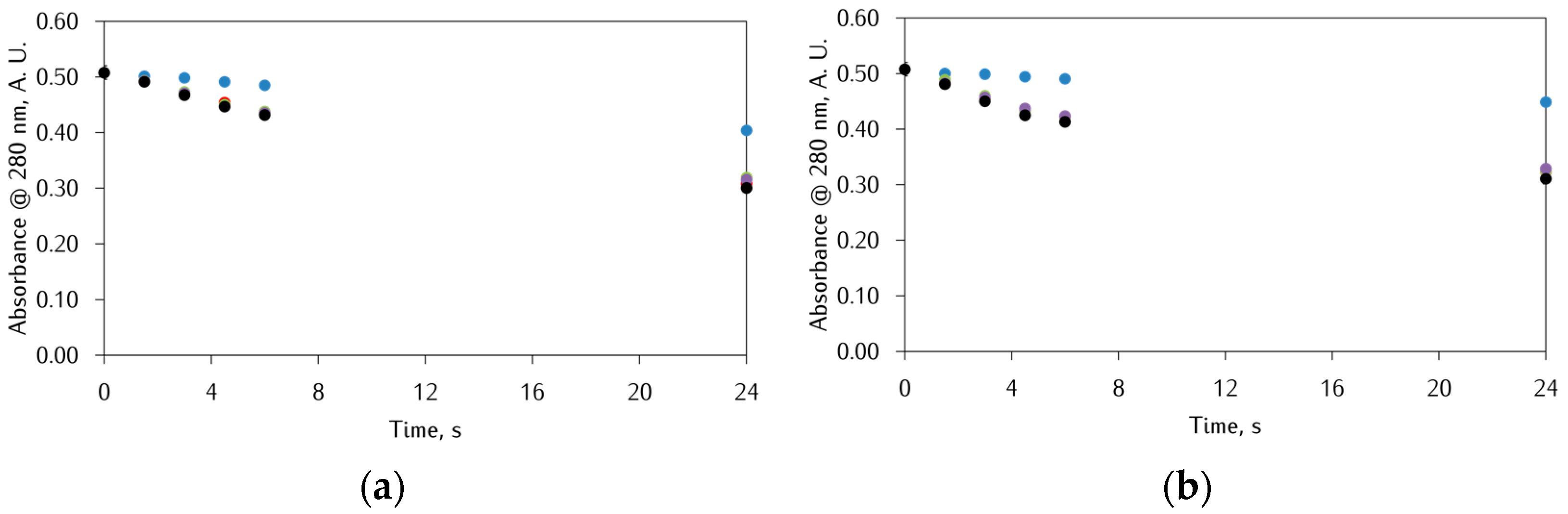
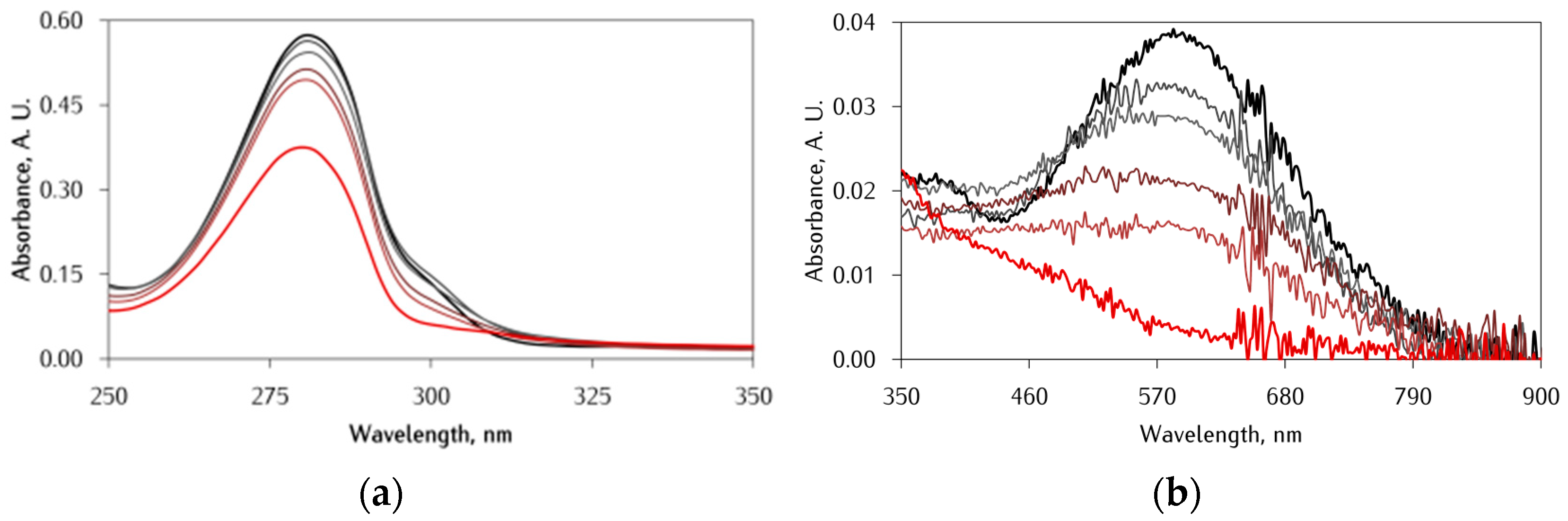
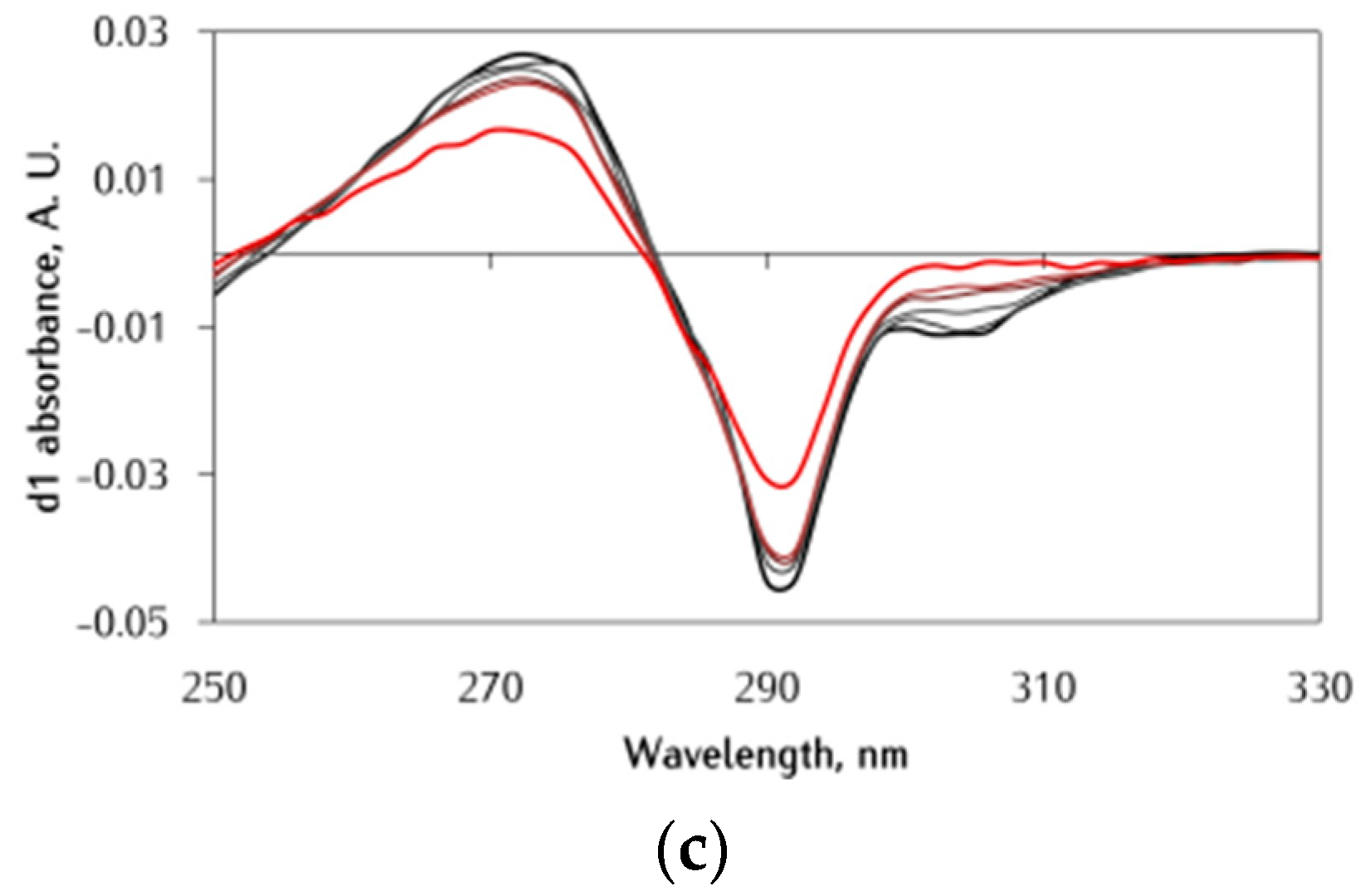
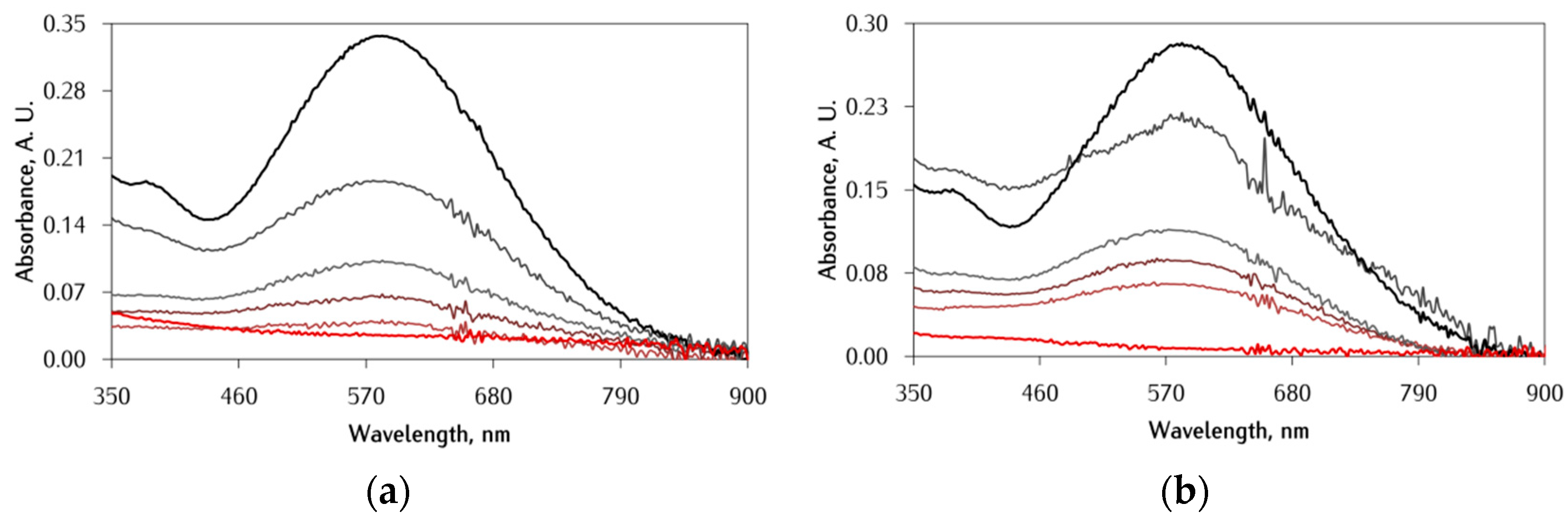
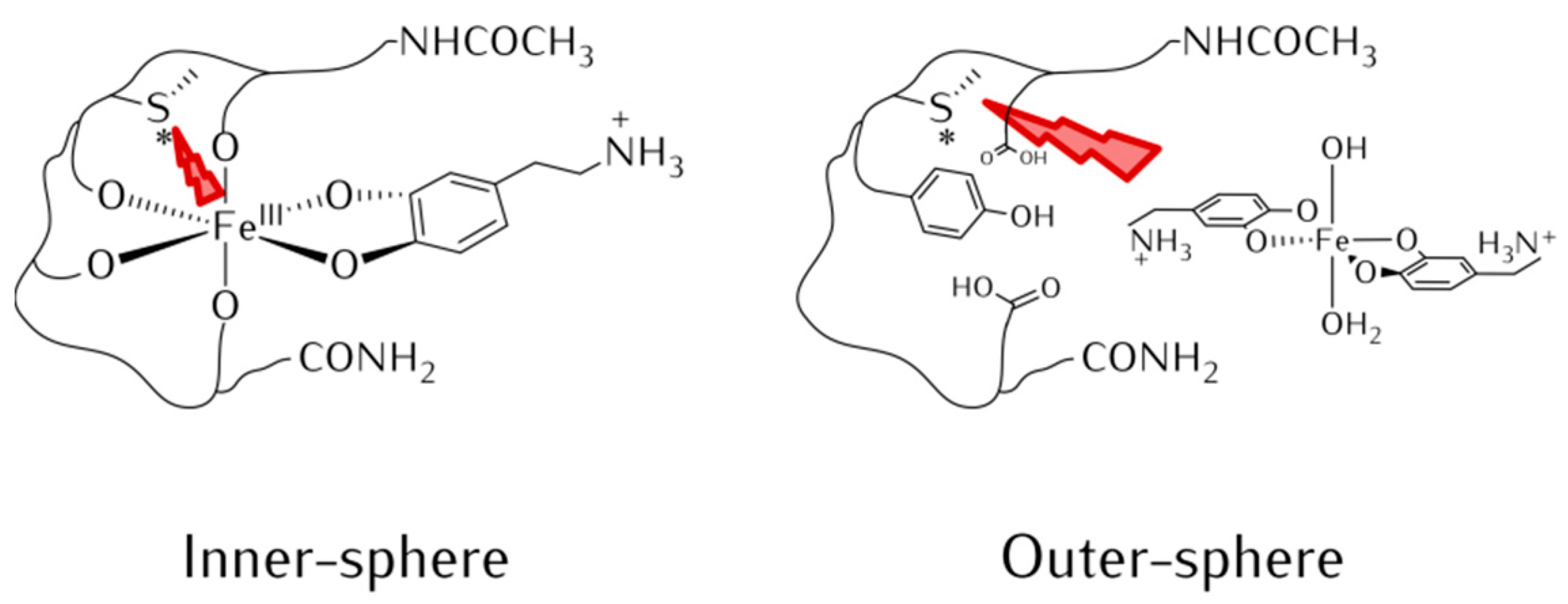
3.1. Effect of αS C-Terminal Peptides on FeIII-DA Interaction
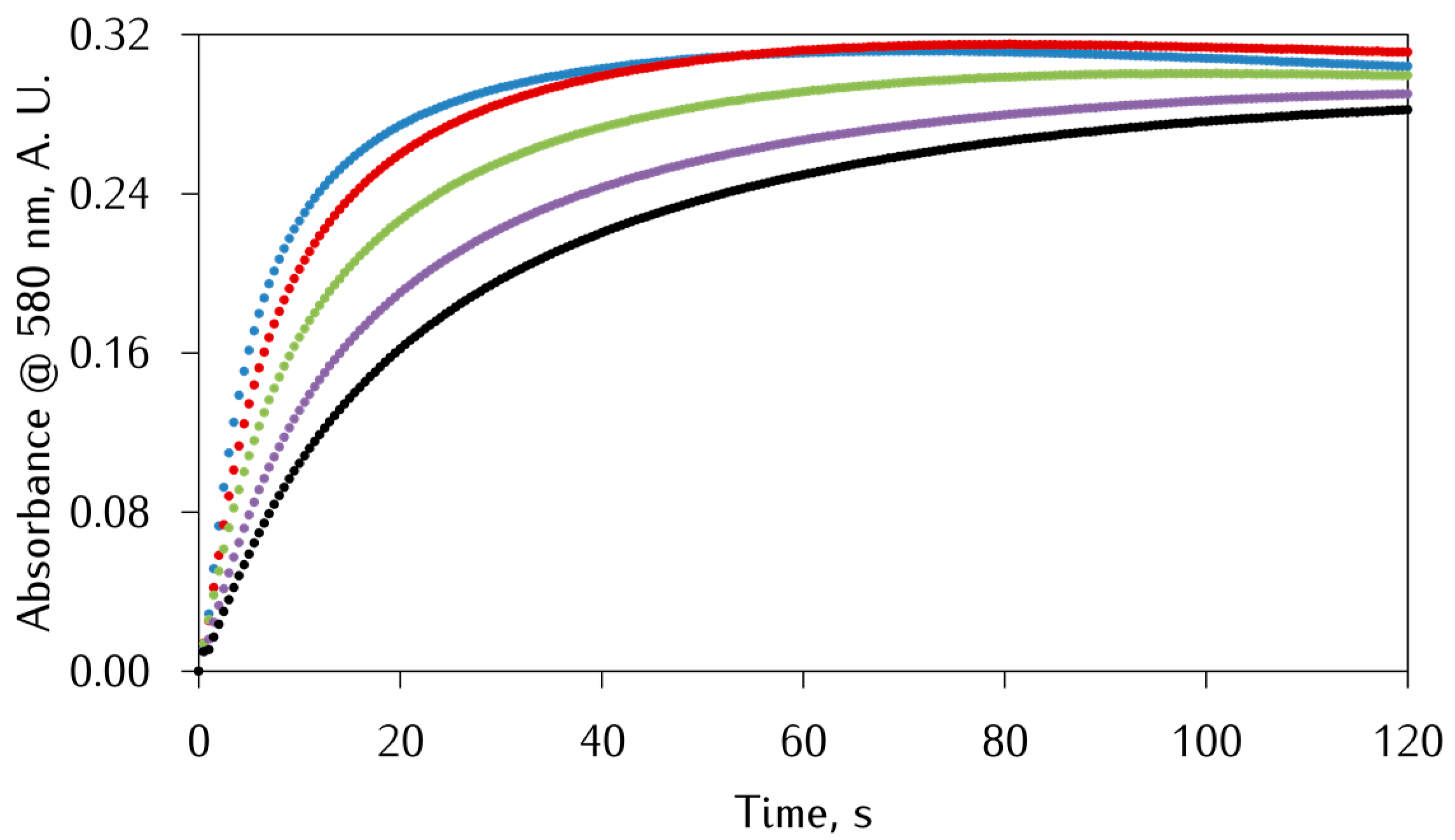
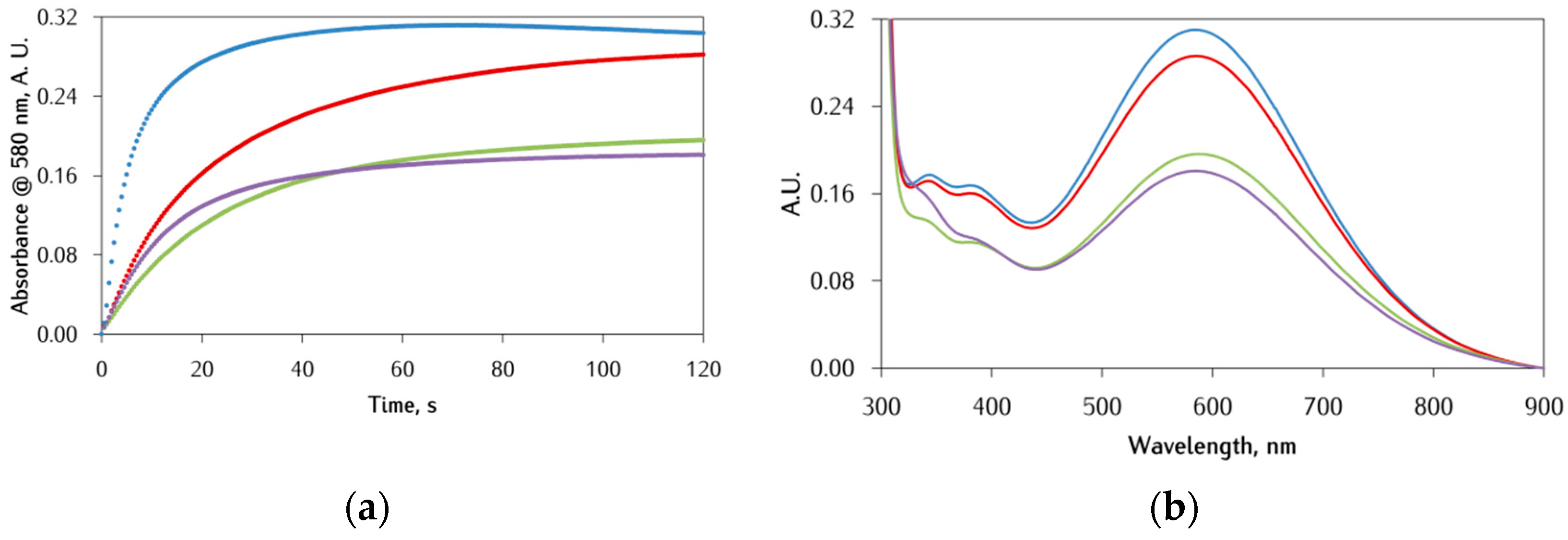
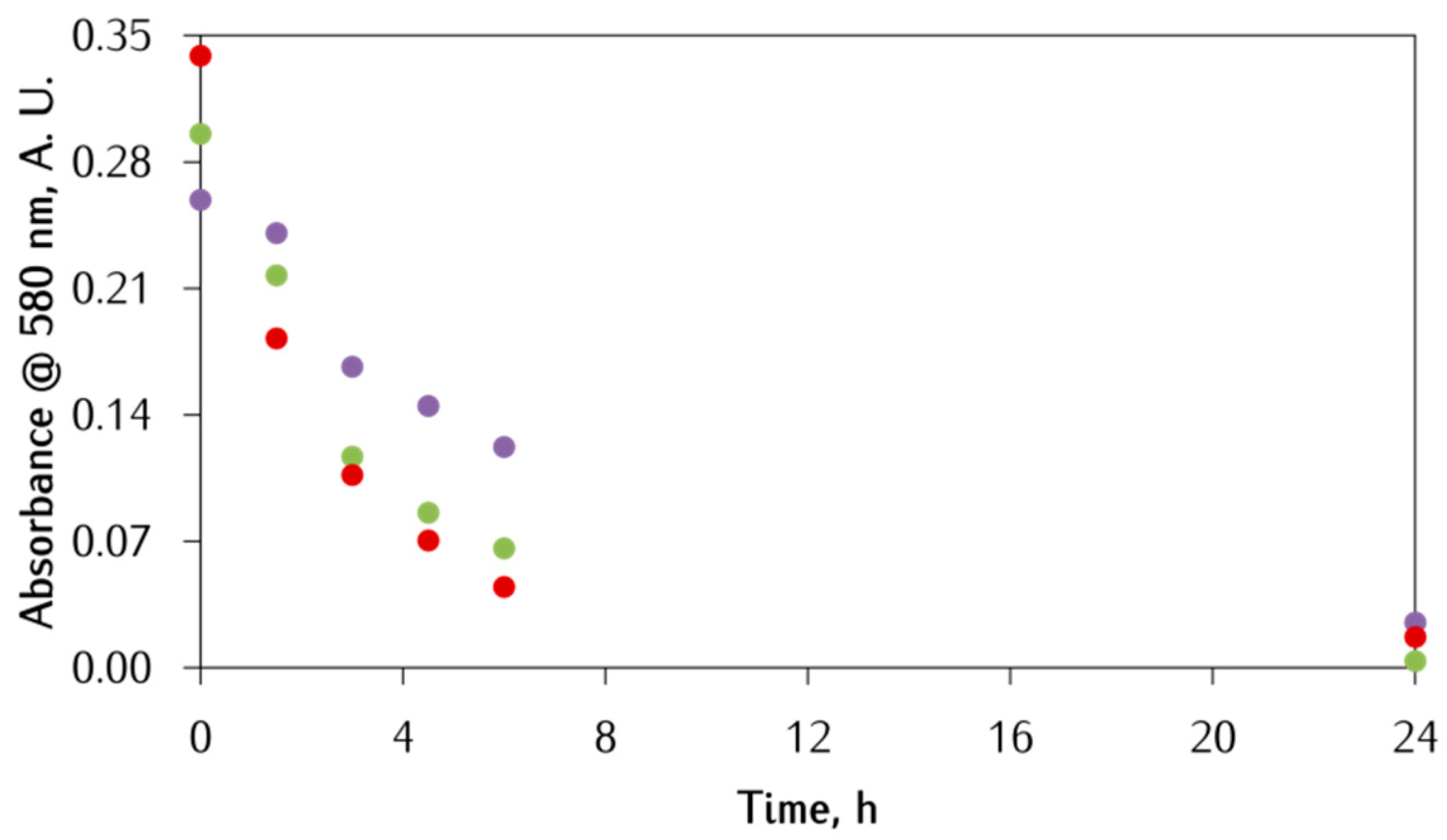
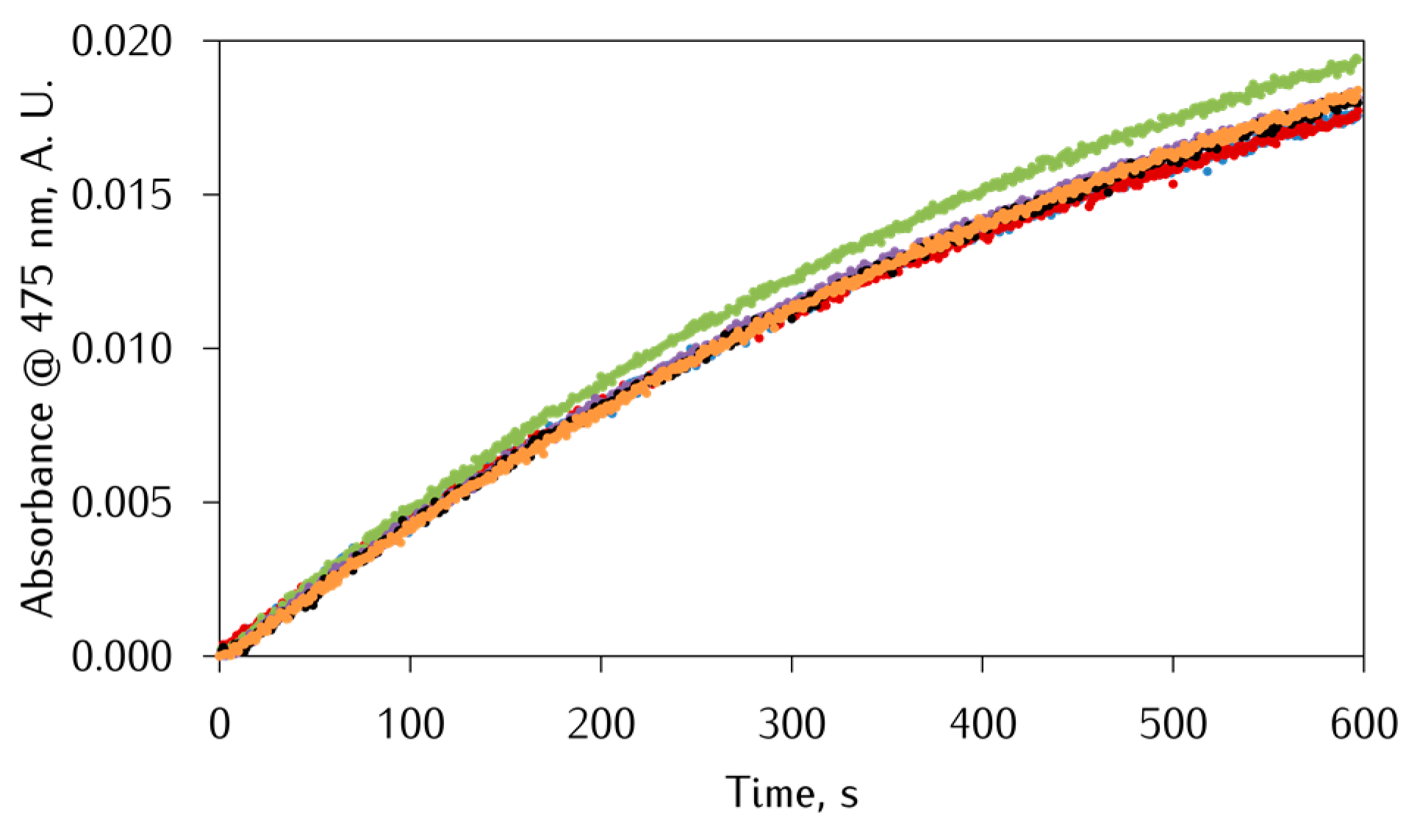
3.2. Kinetic Studies on the Oxidation of Catechol Substrates Promoted by FeIII-Peptide Complexes
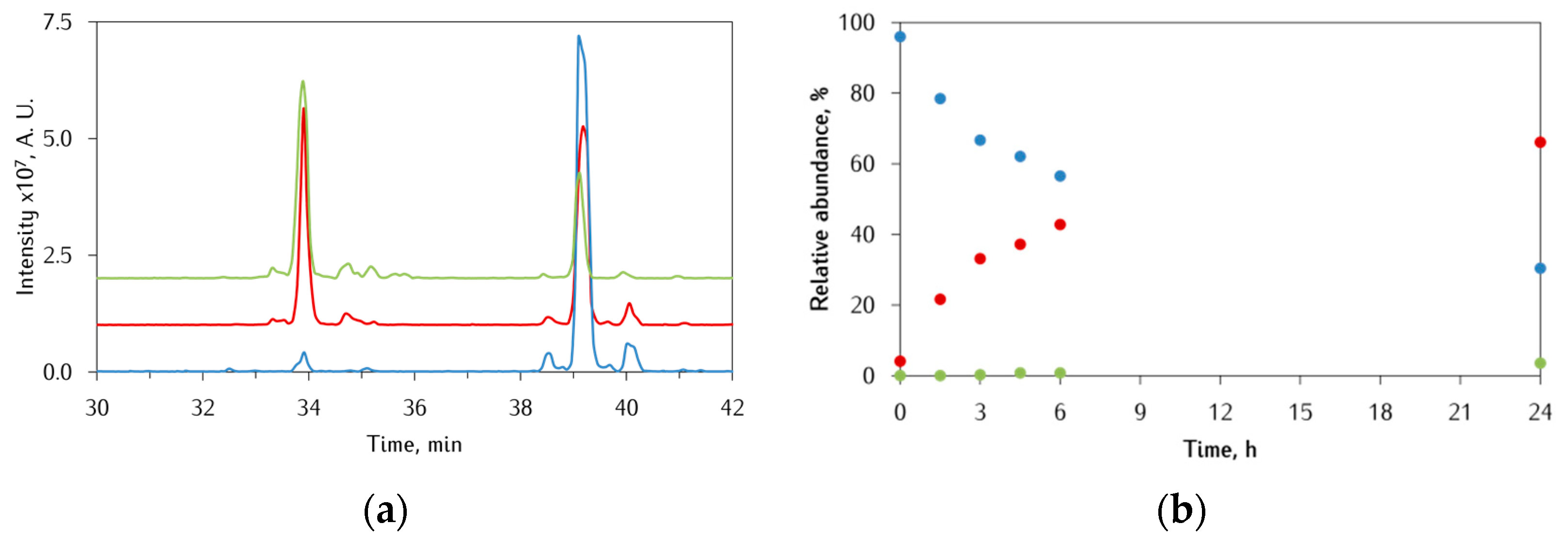
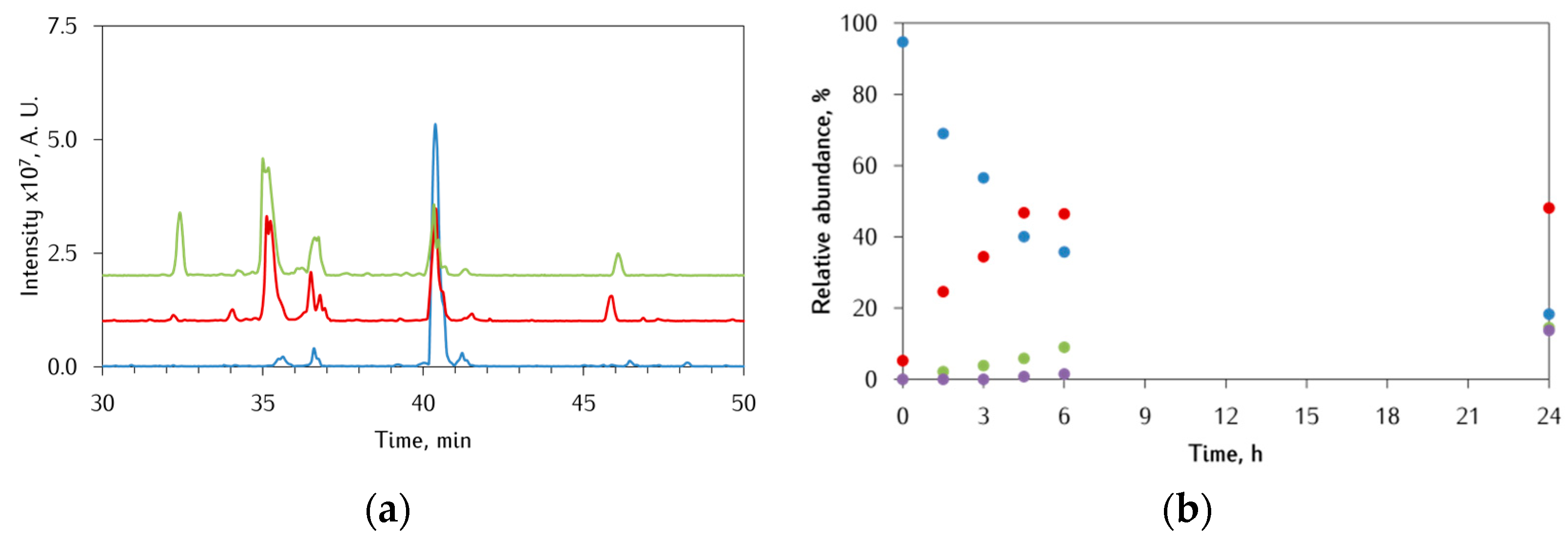
3.3. Identification of αS-Peptide Modifications by HPLC-MS
3.3.1. Ac-αS119–132 Peptide
3.3.2. Ac-αSpS119–132 Peptide
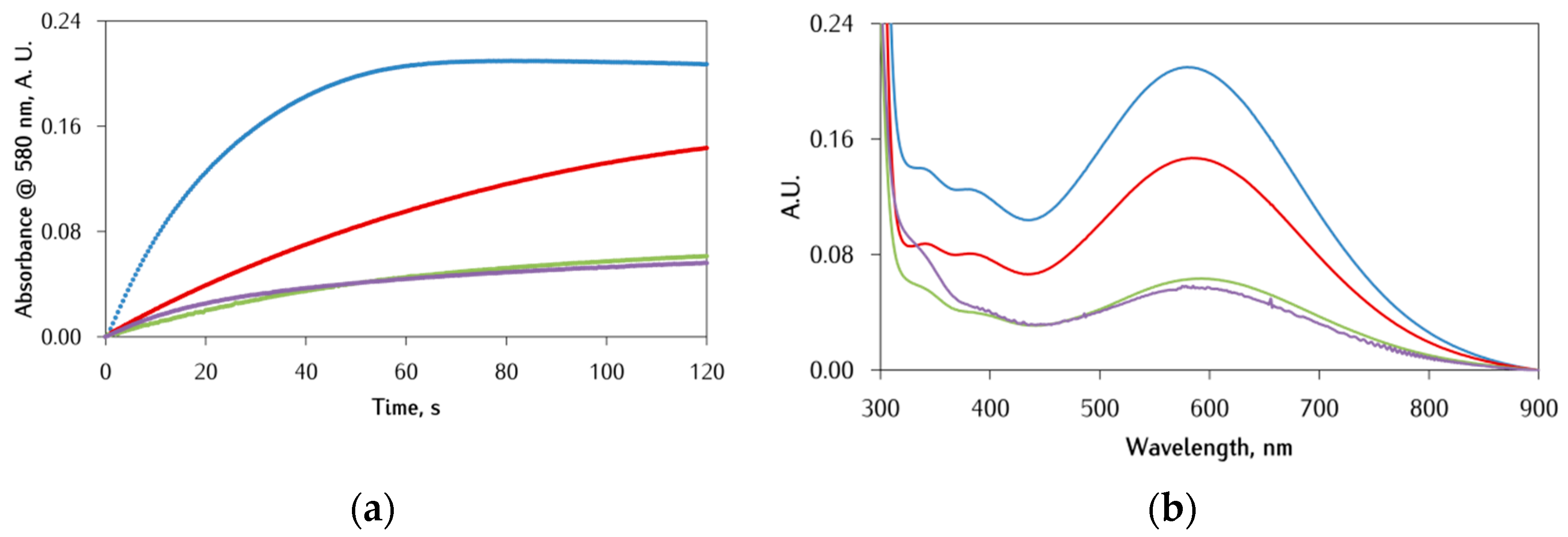
3.4. Comparison with Copper
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wise, R.M.; Wagener, A.; Fietzek, U.M.; Klopstock, T.; Mosharov, E.V.; Zucca, F.A.; Sulzer, D.; Zecca, L.; Burbulla, L.F. Interactions of dopamine, iron, and alpha-synuclein linked to dopaminergic neuron vulnerability in Parkinson’s disease and Neurodegeneration with Brain Iron Accumulation disorders. Neurobiol Dis. 2022, 175, 105920. [Google Scholar] [CrossRef]
- Hare, D.J.; Double, K.L. Iron and dopamine: A toxic couple. Brain 2016, 139, 1026–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingor, P.; Carboni, E.; Koch, J.C. Alpha-synuclein and iron: Two keys unlocking Parkinson’s disease. J. Neural Transm. 2017, 124, 973–981. [Google Scholar] [CrossRef]
- Rothman, R.J.; Serroni, A.; Farber, J.L. Cellular pool of transient ferric iron, chelatable by deferoxamine and distinct from ferritin, that is involved in oxidative cell injury. Mol. Pharm. 1992, 42, 703–710. [Google Scholar]
- Brawley, H.N.; Kreinbrink, A.C.; Hierholzer, J.D.; Vali, S.W.; Lindahl, P.A. Labile Iron Pool of Isolated Escherichia coli Cytosol Likely Includes Fe-ATP and Fe-Citrate but not Fe-Glutathione or Aqueous Fe. J. Am. Chem. Soc. 2023, 145, 2104–2117. [Google Scholar] [CrossRef]
- Zucca, F.A.; Segura-Aguilar, J.; Ferrari, E.; Muñoz, P.; Paris, I.; Sulzer, D.; Sarna, T.; Casella, L.; Zecca, L. Interactions of iron, dopamine and neuromelanin pathways in brain aging and Parkinson’s disease. Prog. Neurobiol. 2017, 155, 96–119. [Google Scholar] [CrossRef]
- Monzani, E.; Nicolis, S.; Dell’Acqua, S.; Capucciati, A.; Bacchella, C.; Zucca, F.A.; Mosharov, E.V.; Sulzer, D.; Zecca, L.; Casella, L. Dopamine, Oxidative Stress and Protein-Quinone Modifications in Parkinson’s and Other Neurodegenerative Diseases. Angew. Chem. Int. Ed. Engl. 2019, 58, 6512–6527. [Google Scholar] [CrossRef] [PubMed]
- Bisaglia, M.; Mammi, S.; Bubacco, L. Kinetic and structural analysis of the early oxidation products of dopamine: Analysis of the interactions with alpha-synuclein. J. Biol. Chem. 2007, 282, 15597–15605. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Pham, A.N.; Waite, T.D. Elucidation of the interplay between Fe(II), Fe(III), and dopamine with relevance to iron solubilization and reactive oxygen species generation by catecholamines. J. Neurochem. 2016, 137, 955–968. [Google Scholar] [CrossRef]
- Sun, Y.; Pham, A.N.; Hare, D.J.; Waite, T.D. Kinetic modeling of pH-dependent oxidation of dopamine by iron and its relevance to Parkinson’s disease. Front. Neurosci. 2018, 12, 859. [Google Scholar] [CrossRef] [PubMed]
- Viles, J.H. Metal ions and amyloid fiber formation in neurodegenerative diseases. Copper, zinc and iron in Alzheimer’s, Parkinson’s and prion diseases. Coord. Chem. Rev. 2012, 256, 2271–2284. [Google Scholar] [CrossRef]
- Binolfi, A.; Rasia, R.M.; Bertoncini, C.W.; Ceolin, M.; Zweckstetter, M.; Griesinger, C.; Jovin, T.M.; Fernández, C.O. Interaction of alpha-synuclein with divalent metal ions reveals key differences: A link between structure, binding specificity and fibrillation enhancement. J. Am. Chem. Soc. 2006, 128, 9893–9901. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Prudent, M.; Fauvet, B.; Lashuel, H.A.; Girault, H.H. Phosphorylation of α-Synuclein at Y125 and S129 Alters Its Metal Binding Properties: Implications for Understanding the Role of α-Synuclein in the Pathogenesis of Parkinson’s Disease and Related Disorders. ACS Chem. Neurosci. 2011, 2, 667–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.L.; Franz, K.J. Phosphorylation of an α-synuclein peptide fragment enhances metal binding. J. Am. Chem. Soc. 2005, 127, 9662–9663. [Google Scholar] [CrossRef]
- Rao, K.S.J. Thermodynamics imprinting reveals differential binding of metals to α-synuclein: Relevance to parkinson’s disease. Biochem. Biophys. Res. Commun. 2007, 359, 115–120. [Google Scholar] [CrossRef]
- Stefánsson, A. Iron (III) hydrolysis and solubility at 25 degrees C. Environ. Sci. Technol. 2007, 41, 6117–6123. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger, L.; DeLano, W. PyMOL. 2020. Available online: http://www.pymol.org/pymol (accessed on 1 October 2022).
- Anderson, J.P.; Walker, D.E.; Goldstein, J.M.; de Laat, R.; Banducci, K.; Caccavello, R.J.; Barbour, R.; Huang, J.; Kling, K.; Lee, M.; et al. Phosphorylation of Ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J. Biol. Chem. 2006, 281, 29739–29752. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.L.; Franz, K.J. Phosphorylation-dependent metal binding by α-synuclein peptide fragments. J. Biol. Inorg. Chem. 2007, 12, 234–247. [Google Scholar] [CrossRef]
- Xu, Y.; Deng, Y.; Qing, H. The phosphorylation of α-synuclein: Development and implication for the mechanism and therapy of the Parkinson’s disease. J. Neurochem. 2015, 135, 4–18. [Google Scholar] [CrossRef]
- Chen, L.; Periquet, M.; Wang, X.; Negro, A.; McLean, P.J.; Hyman, B.T.; Feany, M.B. Tyrosine and serine phosphorylation of alpha-synuclein have opposing effects on neurotoxicity and soluble oligomer formation. J. Clin. Investig. 2009, 119, 3257–3265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, W.S.; Jonas, A.; Clayton, D.F.; George, J.M. Stabilization of alpha-synuclein secondary structure upon binding to synthetic membranes. J. Biol. Chem. 1998, 273, 9443–9449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurochka, A.S.; Yushchenko, D.A.; Bouř, P.; Shvadchak, V.V. Influence of Lipid Membranes on α-Synuclein Aggregation. ACS Chem. Neurosci. 2021, 12, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Bacchella, C.; Camponeschi, F.; Kolkowska, P.; Kola, A.; Tessari, I.; Baratto, M.C.; Bisaglia, M.; Monzani, E.; Bubacco, L.; Mangani, S.; et al. Copper Binding and Redox Activity of α-Synuclein in Membrane-Like Environment. Biomolecules 2023, 13, 287. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mörman, C.; Sternke-Hoffmann, R.; Huang, C.Y.; Prota, A.; Ma, P.; Luo, J. Cu(2+) ions modulate the interaction between α-synuclein and lipid membranes. J. Inorg. Biochem. 2022, 236, 111945. [Google Scholar] [CrossRef] [PubMed]
- Bacchella, C.; Nicolis, S.; Dell’Acqua, S.; Rizzarelli, E.; Monzani, E.; Casella, L. Membrane Binding Strongly Affecting the Dopamine Reactivity Induced by Copper Prion and Copper/Amyloid-beta (Abeta) Peptides. A Ternary Copper/Abeta/Prion Peptide Complex Stabilized and Solubilized in Sodium Dodecyl Sulfate Micelles. Inorg. Chem. 2020, 59, 900–912. [Google Scholar] [CrossRef]
- Sun, Y.; Pham, A.N.; Hider, R.C.; Zheng, H.; Waite, T.D. Effectiveness of the Iron Chelator CN128 in Mitigating the Formation of Dopamine Oxidation Products Associated with the Progression of Parkinson’s Disease. ACS Chem. Neurosci. 2020, 11, 3646–3657. [Google Scholar] [CrossRef]
- Pirota, V.; Dell’Acqua, S.; Monzani, E.; Nicolis, S.; Casella, L. Copper-Aβ Peptides and Oxidation of Catecholic Substrates: Reactivity and Endogenous Peptide Damage. Chem.—A Eur. J. 2016, 22, 16964–16973. [Google Scholar] [CrossRef]
- Morgan, B.; Lahav, O. The effect of pH on the kinetics of spontaneous Fe(II) oxidation by O2 in aqueous solution—Basic principles and a simple heuristic description. Chemosphere 2007, 68, 2080–2084. [Google Scholar] [CrossRef]
- Sever, M.J.; Wilker, J.J. Visible absorption spectra of metal–catecholate and metal–tironate complexes. J. Chem. Society. Dalton Trans. 2004, 4, 1061–1072. [Google Scholar] [CrossRef]
- Herlinger, E.; Jameson, R.F.; Linert, W. Spontaneous autoxidation of dopamine. J. Chem. Soc. Perkin Trans. 1995, 2, 259–263. [Google Scholar] [CrossRef]
- Jiang, D.; Shi, S.; Zhang, L.; Liu, L.; Ding, B.; Zhao, B.; Yagnik, G.; Zhou, F. Inhibition of the Fe(III)-catalyzed dopamine oxidation by ATP and its relevance to oxidative stress in Parkinson’s disease. ACS Chem. Neurosci. 2013, 4, 1305–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachin, K.M.; Karpe, S.A.; Singh, M.; Bhattarai, A. Self-assembly of sodium dodecylsulfate and dodecyltrimethylammonium bromide mixed surfactants with dyes in aqueous mixtures. R. Soc. Open Sci. 2019, 6, 181979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niraula, T.P.; Shah, S.K.; Chatterjee, S.K.; Bhattarai, A. Effect of methanol on the surface tension and viscosity of sodiumdodecyl sulfate (SDS) in aqueous medium at 298.15–323.15 K. Karbala Int. J. Mod. Sci. 2018, 4, 26–34. [Google Scholar] [CrossRef]
- Barreto, W.J.; Barreto, S.R.G.; Ponzoni, S.; Kawano, Y.; Di Mauro, E.; Magosso, H.A.; Silva, W.P. Preparation and characterization of a stable semiquinone-iron complex. Mon. Chem. 2005, 136, 701–712. [Google Scholar] [CrossRef]
- Young, A.R. Chromophores in human skin. Phys. Med. Biol. 1997, 42, 789–802. [Google Scholar] [CrossRef]
- Greenough, M.A.; Camakaris, J.; Bush, A.I. Metal dyshomeostasis and oxidative stress in Alzheimer’s disease. Neurochem. Int. 2013, 62, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Morris, H.; Cronin, M.T. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrastro, I.; Pasha, S.; Jensen, K.T.; Pitt, A.R.; Spickett, C.M. Mass spectrometry-based methods for identifying oxidized proteins in disease: Advances and challenges. Biomolecules 2015, 5, 378–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacchella, C.; Dell’Acqua, S.; Nicolis, S.; Monzani, E.; Casella, L. The reactivity of copper complexes with neuronal peptides promoted by catecholamines and its impact on neurodegeneration. Coord. Chem. Rev. 2022, 471, 214756. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
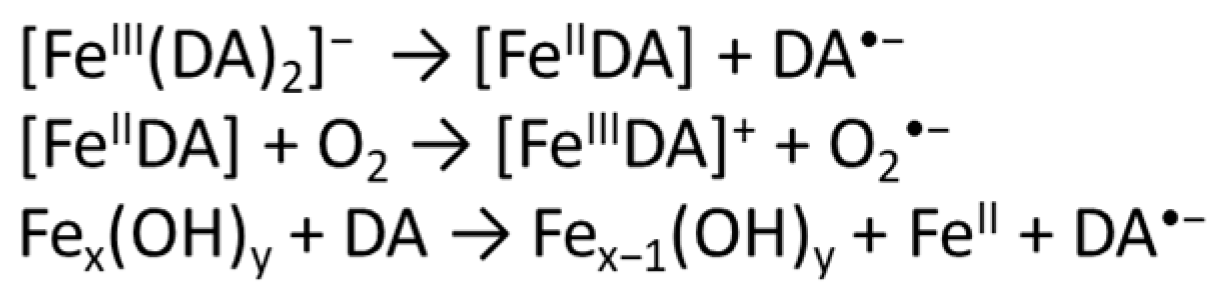{kind=link}
{kind=link}
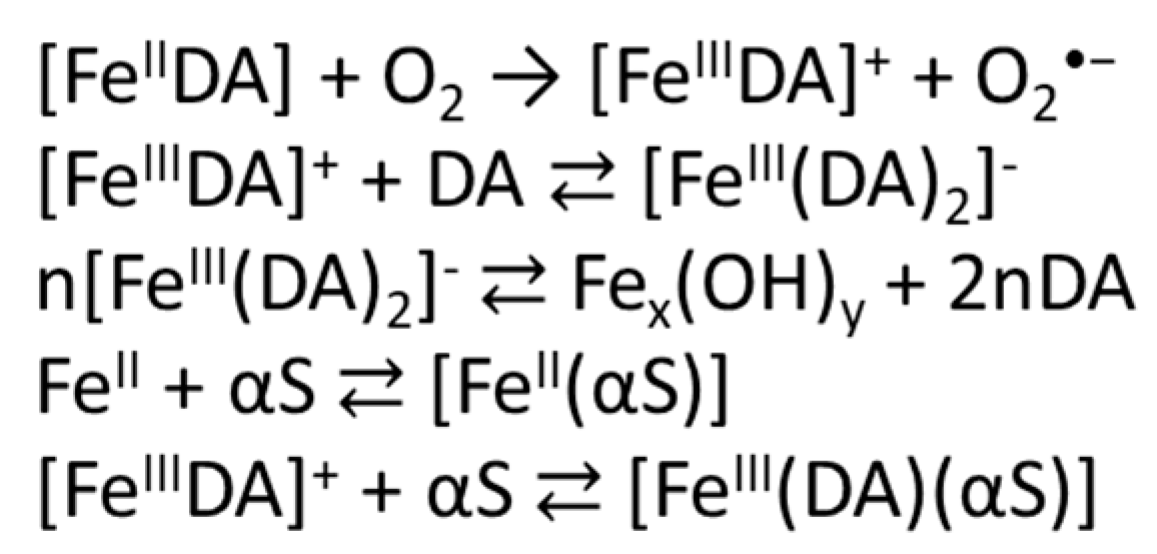{kind=link}
{kind=link}
| DA:Fe:Peptide (Equiv.) | Incubation Time (h) | Ac-αS119–132 +16 (%) |
|---|---|---|
| 2:1:2 | 6 | 43 |
| 2:1:4 | 6 | 33 |
| 20:1:2 | 6 | 15 |
| 20:1:4 | 6 | 28 |
| DA:Fe:Peptide (Mol Ratio) | Incubation Time (h) | Ac-αSpS119–132 +16 (%) |
|---|---|---|
| 2:1:2 | 6 | 56 |
| 2:1:4 | 6 | 29 |
| 20:1:2 | 6 | 54 |
| 20:1:4 | 6 | 41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schifano, F.; Dell’Acqua, S.; Nicolis, S.; Casella, L.; Monzani, E. Interaction and Redox Chemistry between Iron, Dopamine, and Alpha-Synuclein C-Terminal Peptides. Antioxidants 2023, 12, 791. https://doi.org/10.3390/antiox12040791
Schifano F, Dell’Acqua S, Nicolis S, Casella L, Monzani E. Interaction and Redox Chemistry between Iron, Dopamine, and Alpha-Synuclein C-Terminal Peptides. Antioxidants. 2023; 12(4):791. https://doi.org/10.3390/antiox12040791
Chicago/Turabian StyleSchifano, Fabio, Simone Dell’Acqua, Stefania Nicolis, Luigi Casella, and Enrico Monzani. 2023. "Interaction and Redox Chemistry between Iron, Dopamine, and Alpha-Synuclein C-Terminal Peptides" Antioxidants 12, no. 4: 791. https://doi.org/10.3390/antiox12040791
APA StyleSchifano, F., Dell’Acqua, S., Nicolis, S., Casella, L., & Monzani, E. (2023). Interaction and Redox Chemistry between Iron, Dopamine, and Alpha-Synuclein C-Terminal Peptides. Antioxidants, 12(4), 791. https://doi.org/10.3390/antiox12040791